The lin-4 Gene Controls Fat Accumulation and Longevity in Caenorhabditis elegans

Abstract
:1. Introduction
2. Results and Discussion
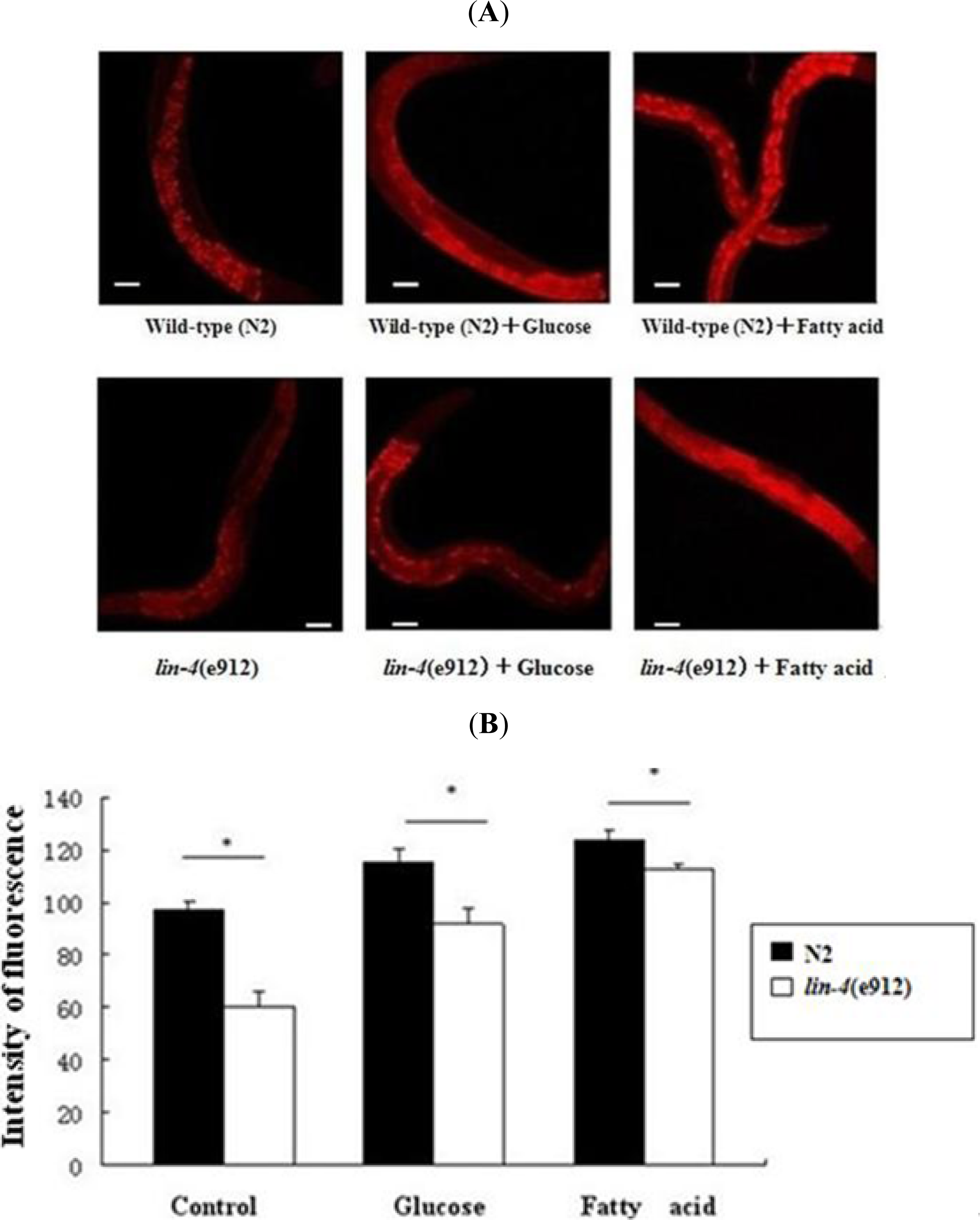
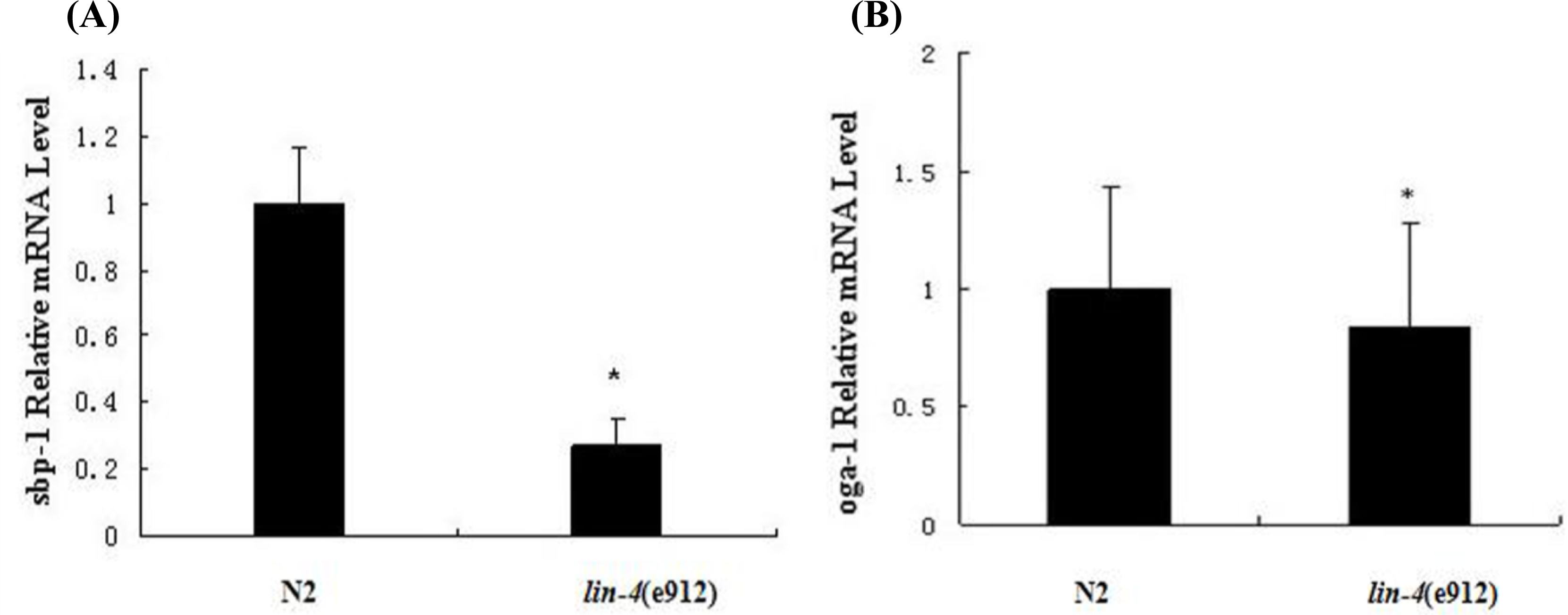
2.1. The Fat Content Is Remarkably Reduced in lin-4 Mutants
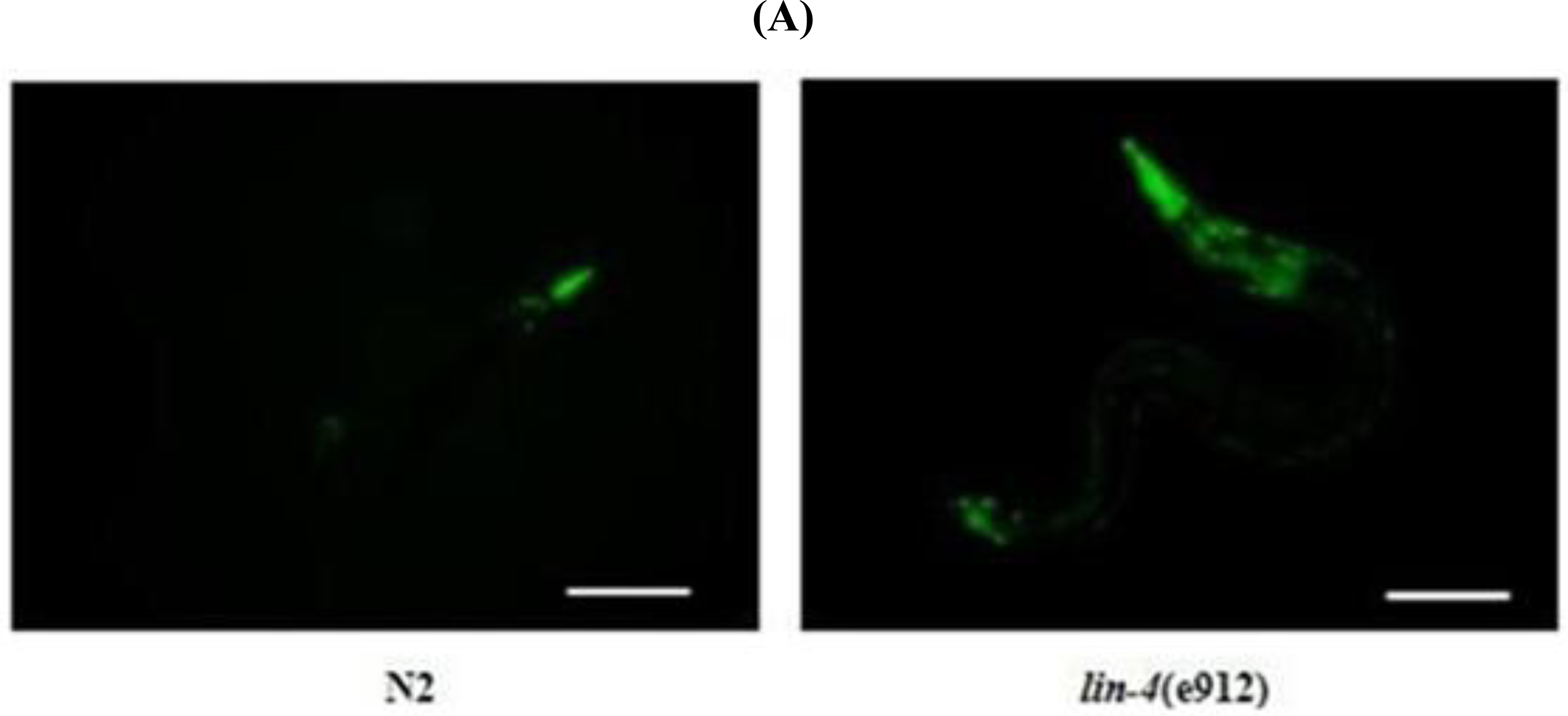
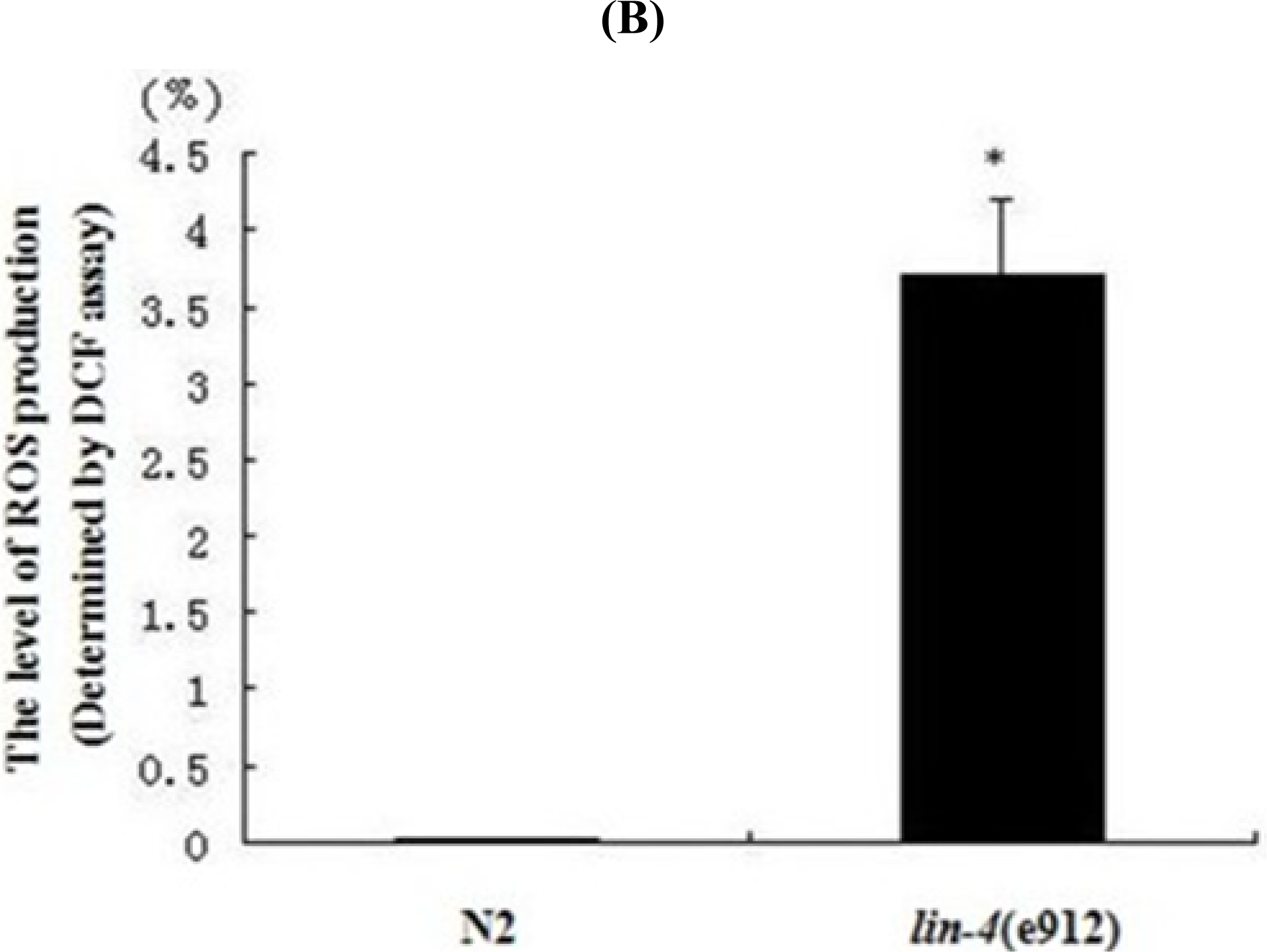
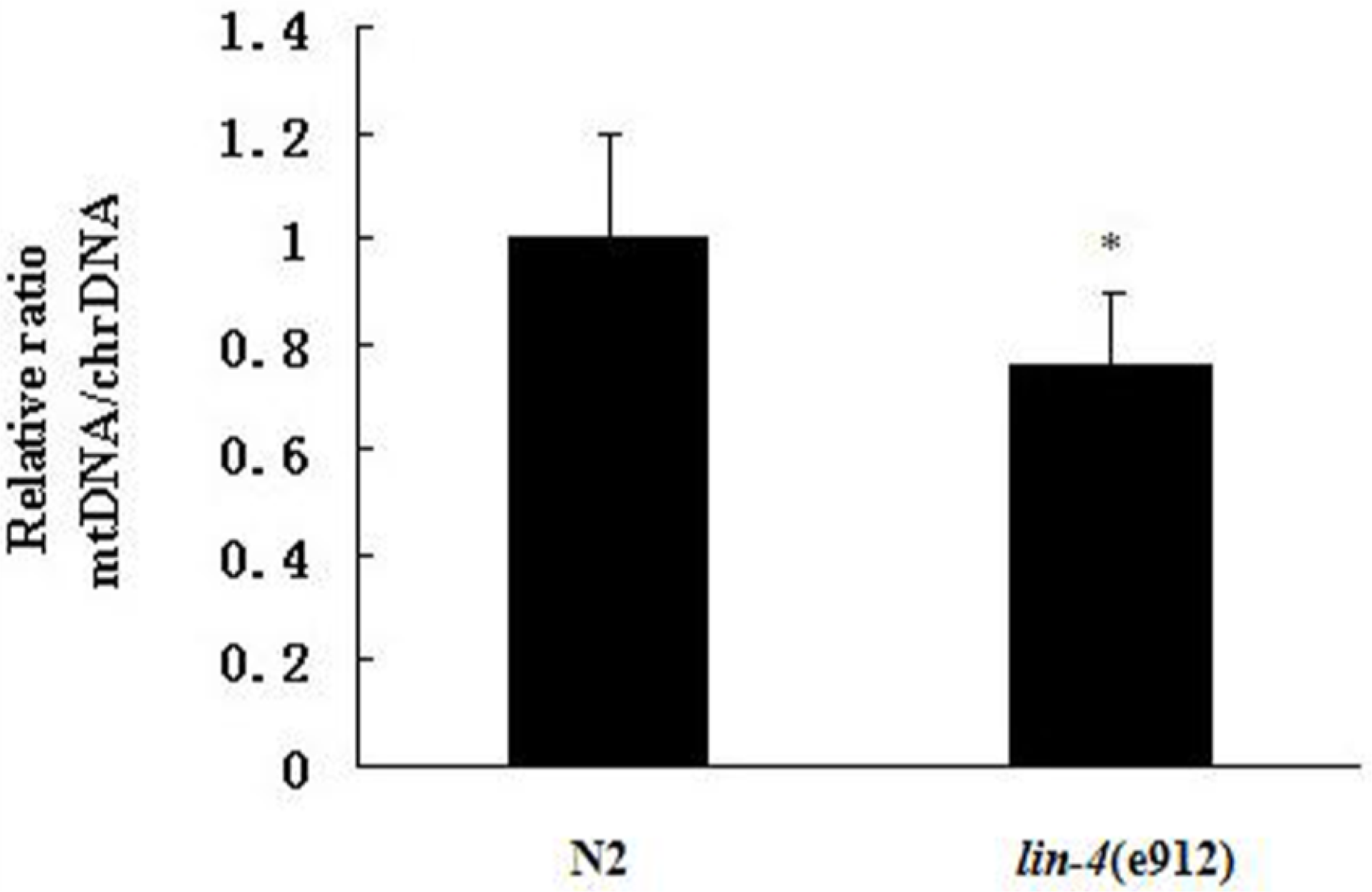
2.2. ROS Accumulation and Decreased Copy Number of mtDNA in lin-4 Mutants
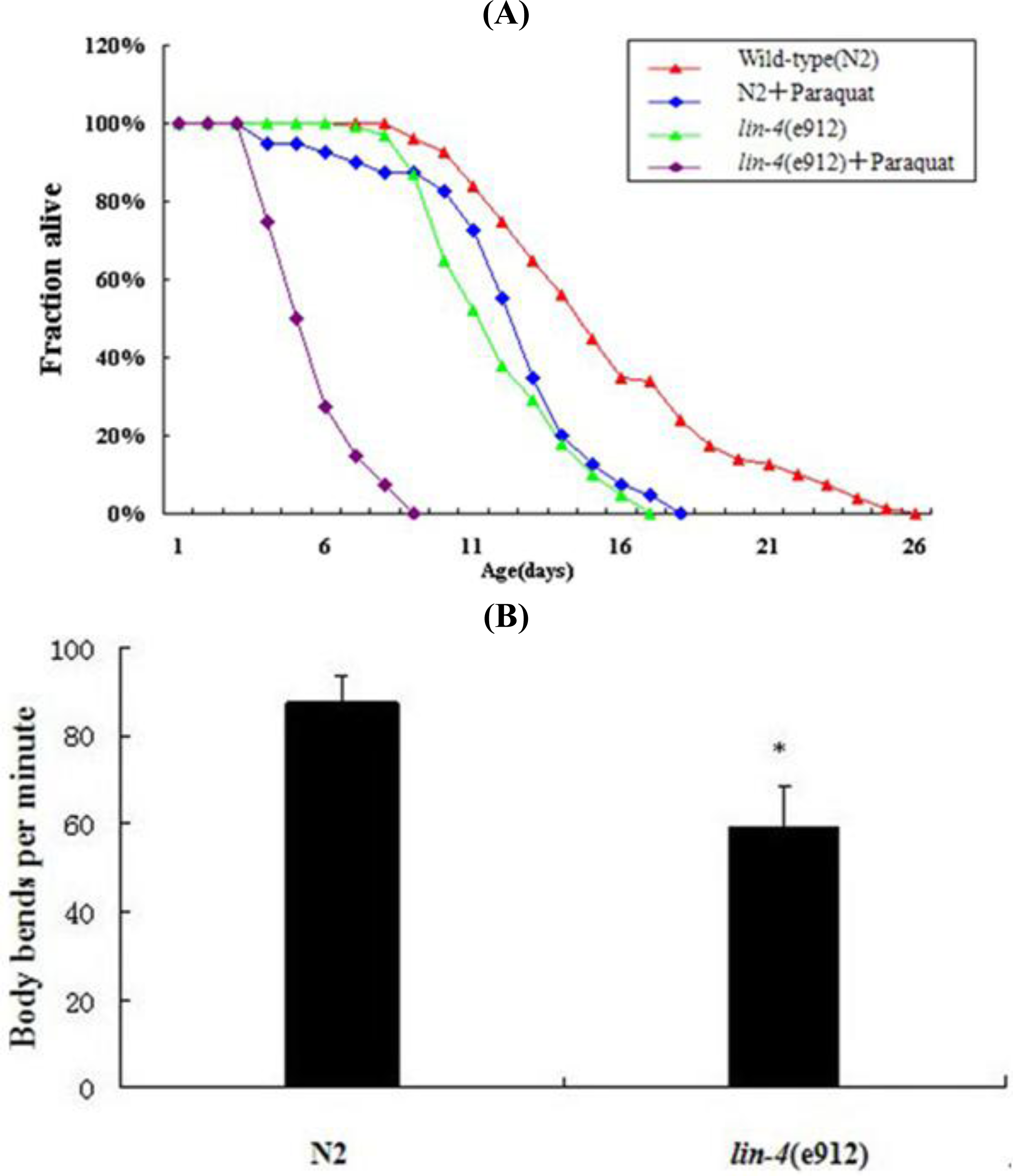
2.3. Effect of lin-4 on the Life Span and Locomotion of C. elegans
2.4. Discussion
3. Experimental Methods
3.1. Worms and Culture
3.2. Nile Red Staining
3.3. Effect of Fatty Acid
3.4. Effect of Glucose
3.5. Effect of Paraquat
3.6. Measurement of Intracellular ROS in C. elegans
3.7. Real-Time PCR of mtDNA
3.8. Life Span and Locomotion Assay
3.9. Quantitative RT-PCR
3.10. Statistical Analysis
4. Conclusion
Acknowledgments
References
- Bartel, DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar]
- Wienholds, E; Plasterk, RH. MicroRNA function in animal development. FEBS Lett 2005, 579, 5911–5922. [Google Scholar]
- Heneghan, HM; Miller, N; Kerin, MJ. Role of microRNAs in obesity and the metabolic syndrome. Obes. Rev 2010, 11, 354–361. [Google Scholar]
- Vella, MC; Slack, FJC. Elegans microRNAs. Worm Book 2005, 21, 1–9. [Google Scholar]
- Olsen, PH; Ambros, V. The lin-4 regulatory RNA controls developmental timing in Caenorhabditis elegans by blocking LIN-14 protein synthesis after the initiation of translation. Dev. Biol 1999, 216, 671–680. [Google Scholar]
- Boehm, M; Slack, F. A developmental timing microRNA and its target regulate life span in C. elegans. Science 2005, 310, 1954–1957. [Google Scholar]
- Singhal, V; Schwenk, WF; Kumar, S. Evaluation and management of childhood and adolescent obesity. Mayo Clin. Proc 2007, 82, 1258–1264. [Google Scholar]
- Speakman, J; Hambly, C; Mitchell, S; Krol, E. The contribution of animal models to the study of obesity. Lab. Anim 2008, 42, 413–432. [Google Scholar]
- McKay, RM; McKay, JP; Avery, L; Graff, JM. C elegans: A model for exploring the genetics of fat storage. Dev. Cell 2003, 4, 131–142. [Google Scholar]
- Brooks, KK; Liang, B; Watts, JL. The influence of bacterial diet on fat storage in C. elegans. PLoS One 2009, 4, e7545. [Google Scholar]
- Nomura, T; Horikawa, M; Shimamura, S; Hashimoto, T; Sakamoto, K. Fat accumulation in Caenorhabditis elegans is mediated by SREBP homolog SBP-1. Genes Nutr 2009, 5, 17–27. [Google Scholar]
- Forsythe, ME; Love, DC; Lazarus, BD; Kim, EJ; Prinz, WA; Ashwell, G; Krause, MW; Hanover, JA. Caenorhabditis elegans ortholog of a diabetes susceptibility locus: oga-1 (O-GlcNAcase) knockout impacts O-GlcNAc cycling, metabolism, and dauer. Proc. Natl. Acad. Sci. USA 2006, 103, 11952–11957. [Google Scholar]
- Eberlé, D; Hegarty, B; Bossard, P; Ferré, P; Foufelle, F. SREBP transcription factors: Master regulators of lipid homeostasis. Biochimie 2004, 86, 839–848. [Google Scholar]
- Yang, F; Vought, BW; Satterlee, JS; Walker, AK; Jim Sun, ZY; Watts, JL; DeBeaumont, R; Saito, RM; Hyberts, SG; Yang, S; Macol, C; lyer, L; Tjian, R; van den Heuvel, S; Hart, AC; Wagner, G; Naar, AM. An ARC/Mediator subunit required for SREBP control of cholesterol and lipid homeostasis. Nature 2006, 442, 700–704. [Google Scholar]
- Wells, L; Whelan, SA; Hart, GW. O-GlcNAc: A regulatory post-translational modification. Biochem. Biophys. Res. Commun 2003, 302, 435–441. [Google Scholar]
- González-Polo, RA; Rodríguez-Martín, A; Morán, JM; Niso, M; Soler, G; Fuentes, JM. Paraquat-induced apoptotic cell death in cerebellar granule cells. Brain Res 2004, 1011, 170–176. [Google Scholar]
- Hekimi, S; Guarente, L. Genetics and the specificity of the aging process. Science 2003, 299, 1351–1354. [Google Scholar]
- Braeckman, BP; Houthoofd, K; Brys, K; Lenaerts, I; de Vreese, A; van Eygen, S; Raes, H; Vanfleteren, JR. No reduction of energy metabolism in Clk mutants. Mech. Ageing Dev 2003, 123, 1447–1456. [Google Scholar]
- Ide, T; Tsutsui, H; Hayashidani, S; Kang, D; Suematsu, N; Nakamura, K; Utsumi, H; Hamasaki, N; Takeshita, A. Mitochondrial DNA damage and dysfunction associated with oxidative stress in failing hearts after myocardial infarction. Circ. Res 2001, 88, 529–535. [Google Scholar]
- Beckman, KB; Ames, BN. The free radical theory of aging matures. Physiol. Rev 1998, 78, 547–581. [Google Scholar]
- Boyle, JH; Cohen, N. Caenorhabditis elegans body wall muscles are simple actuators. Biosyststems 2008, 94, 170–181. [Google Scholar]
- Finkel, T; Holbrook, NJ. Oxidants, oxidative stress and the biology of aging. Nature 2000, 408, 239–247. [Google Scholar]
- Murakami, S; Murakami, H. The effects of aging and oxidative stress on learning behavior in C. elegans. Neurobiol. Aging 2005, 26, 899–905. [Google Scholar]
- Kam, PCA; Ferch, NI. Apoptosis: Mechanisms and clinical implications. Anaesthesia 2000, 55, 1081–1093. [Google Scholar]
- Libina, N; Berman, JR; Kenyon, C. Tissue-specific activities of C. elegans DAF-16 in the regulation of lifespan. Cell 2006, 115, 489–502. [Google Scholar]
- Hashimoto, T; Horikawa, M; Nomura, T; Sakamoto, K. Nicotinamide adenine dinucleotide extends the lifespan of Caenorhabditis elegans mediated by sir-2.1 and daf-16. Biogerontology 2010, 11, 31–43. [Google Scholar]
- Horikawa, M; Nomura, T; Hashimoto, T; Sakamoto, K. Elongation and desaturation of fatty acids are critical in growth, lipid metabolism, and ontogeny of Caenorhabditis elegans. J. Biochem 2008, 144, 149–158. [Google Scholar]
- Lee, SJ; Murphy, CT; Kenyon, C. Glucose shortens the life span of C. elegans by downregulating DAF-16/FOXO activity and aquaporin gene expression. Cell Metab 2009, 10, 379–391. [Google Scholar]
- Keaney, M; Matthijssens, F; Sharpe, M; Vanfleteren, J; Gems, D. Superoxide dismutase mimetics elevate superoxide dismutase activity in vivo but do not retard aging in the nematode Caenorhabditis elegans. Free Radic. Biol. Med 2004, 37, 239–250. [Google Scholar]
- Kampkötter, A; Nkwonkam, CG; Zurawski, RF; Timpel, C; Chovolou, Y; Watjen, W; Kahl, R. Investigations of protective effects of the flavonoids quercetin and rutin on stress resistance in the model organism Caenorhabditis elegans. Toxicology 2007, 234, 113–123. [Google Scholar]
- Miller, KG; Alfonso, A; Nguyen, M; Crowell, JA; Johnson, CD; Rand, JB. A genetic selection for Caenorhabditis elegans synaptic transmission mutants. Proc. Natl. Acad. Sci. USA 1996, 93, 12593–12598. [Google Scholar]
- Ye, K; Ji, CB; Lu, XW; Ni, YH; Gao, CL; Chen, XH; Zhao, YP; Gu, GX; Guo, XR. Resveratrol attenuates radiation damage in Caenorhabditis elegans by preventing oxidative stress. J. Radiat. Res 2010, 51, 473–479. [Google Scholar]
- Dillin, A; Crawford, DK; Kenyon, C. Timing requirements for insulin/IGF-1 signaling in C. elegans. Science 2002, 298, 830–834. [Google Scholar]
- Taubert, S; van Gilst, MR; Hansen, M; Yamamoto, KR. A Mediator subunit, MDT-15, integrates regulation of fatty acid metabolism by NHR-49-dependent and independent pathways in C. elegans. Genes Dev 2006, 20, 1137–1149. [Google Scholar]
- Livak, KJ; Schmittgen, TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Wormbase | Primers |
|---|---|---|
| GDP2 (nuclear) | K10B3.8.1 | Probe:ACATAGTCGGCTCCAGAGGCTCC |
| Sense: CGCAGCACAAGATCAAGGTAG | ||
| Anti: AAGACTCCGGTGGACTCAAC | ||
| CYTB (mitochondria) | Mitochondrial | Probe:GGGTCAGCTTCAATAAACATCTCTGC |
| Sense: GGTTATATTGCCACGGTGATTATG | ||
| Anti:AATATCACTCTGGAACAATATGAACTG |
| Symbol | Wormbase | Primers |
|---|---|---|
| Sbp-1 | Y47D3B.7 | Probe:CTTCGCCGCCTTCTCCTTACTCCTCG |
| Sense:CCACCACCTCATCACCACCAT | ||
| Oga-1 | T20B5.3a | Anti: CCTTCCGCTGTCTCCTGAATCA |
| Probe:CAGCCGAGTATTCACAGCCTATGGA | ||
| actin | T04C12.5 | Sense:GAGCCTGTGCCTTCTGAGTTA |
| Anti: CAGTGATAGTCTTTGATTTCTTATTTCCT | ||
| Probe:CATCCTCCGTCTTGACTTGGCTGG | ||
| Sense:CAATCTACGAAGGATATGCCCTC | ||
| Anti: CTCAGCGGTGGTGGTGAA |
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhu, C.; Ji, C.-B.; Zhang, C.-M.; Gao, C.-L.; Zhu, J.-G.; Qin, D.-N.; Kou, C.-Z.; Zhu, G.-Z.; Shi, C.-M.; Guo, X.-R. The lin-4 Gene Controls Fat Accumulation and Longevity in Caenorhabditis elegans . Int. J. Mol. Sci. 2010, 11, 4814-4825. https://doi.org/10.3390/ijms11124814
Zhu C, Ji C-B, Zhang C-M, Gao C-L, Zhu J-G, Qin D-N, Kou C-Z, Zhu G-Z, Shi C-M, Guo X-R. The lin-4 Gene Controls Fat Accumulation and Longevity in Caenorhabditis elegans . International Journal of Molecular Sciences. 2010; 11(12):4814-4825. https://doi.org/10.3390/ijms11124814
Chicago/Turabian StyleZhu, Chun, Chen-Bo Ji, Chun-Mei Zhang, Chun-Lin Gao, Jin-Gai Zhu, Da-Ni Qin, Chun-Zhao Kou, Guan-Zhong Zhu, Chun-Mei Shi, and Xi-Rong Guo. 2010. "The lin-4 Gene Controls Fat Accumulation and Longevity in Caenorhabditis elegans " International Journal of Molecular Sciences 11, no. 12: 4814-4825. https://doi.org/10.3390/ijms11124814