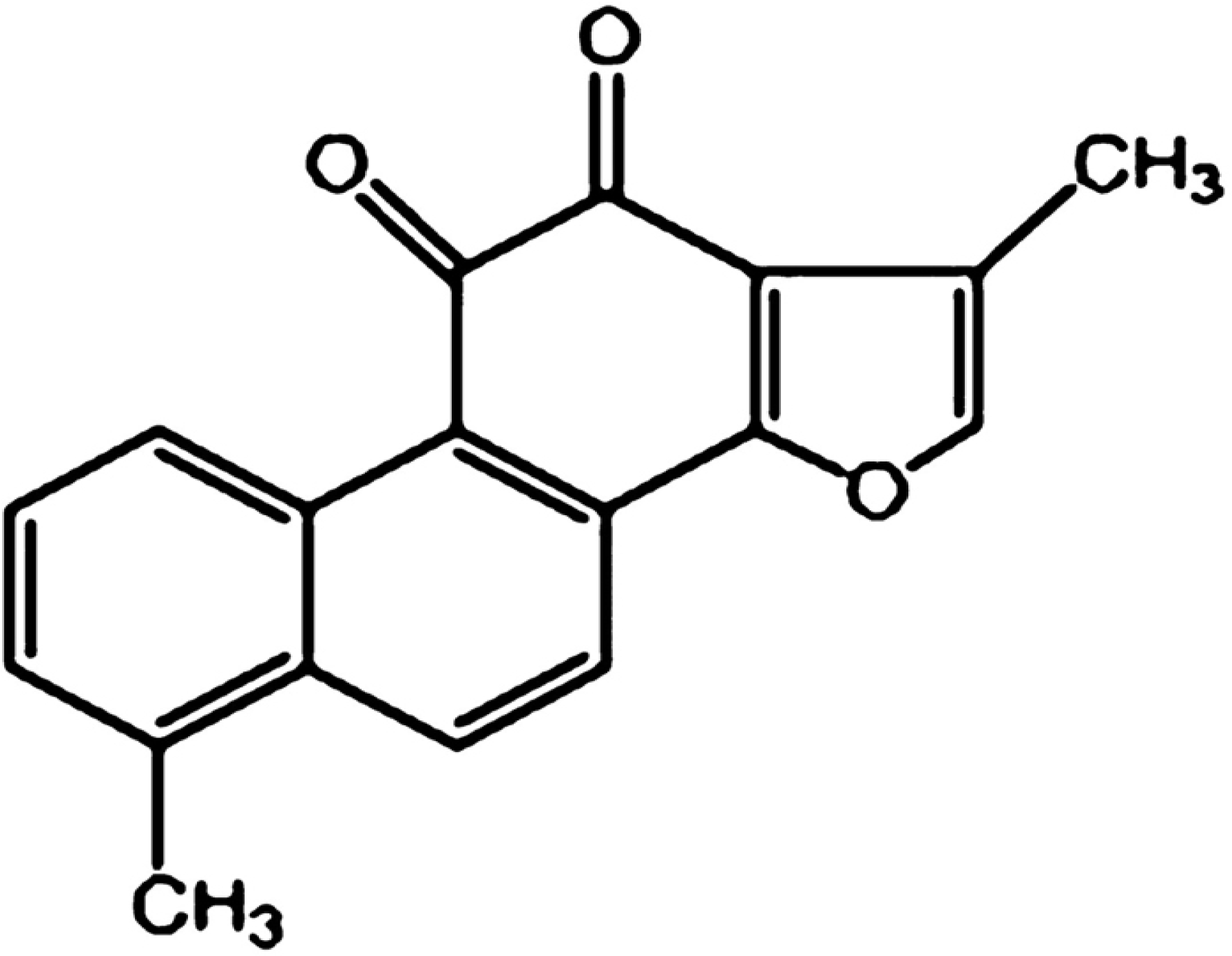
Down-Regulation of Telomerase Activity and Activation of Caspase-3 Are Responsible for Tanshinone I-Induced Apoptosis in Monocyte Leukemia Cells in Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental
2.1. Main Reagents
2.2. Cell Culture
2.3. Cell Growth Inhibitory Rate (MTT Assay)
2.4. Real-Time RT-PCR Detection of Survivin
2.5. Telomerase Activity and hTERT Assay
2.6. Caspase-3 Activity Assay
2.7. Western Blotting Analysis of Caspase-3 and Survivin
2.8. Flow Cytometry (FCM) Detection
2.9. Statistical Analysis
3. Results
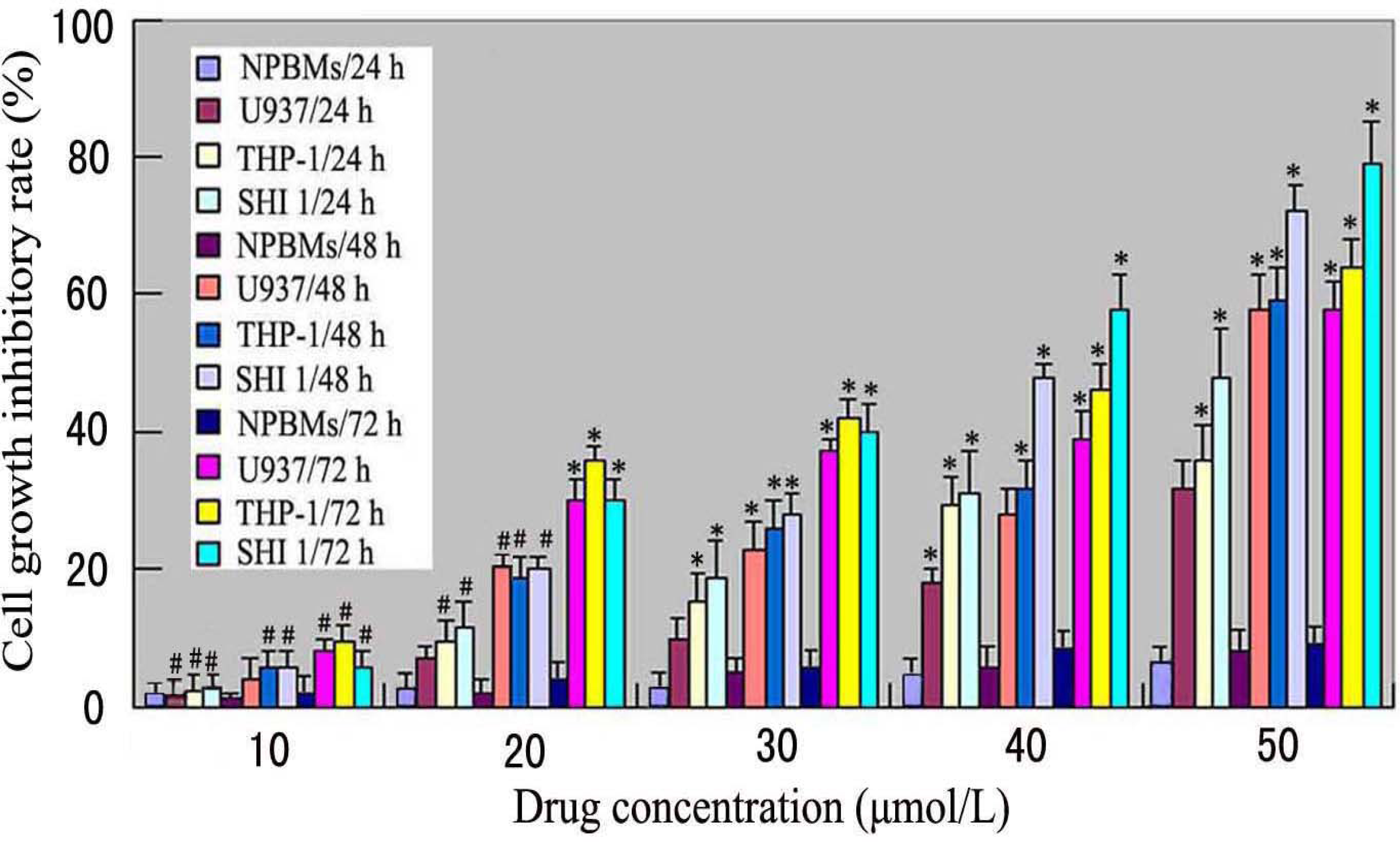
3.1. Tan-I Inhibits Monocytic Leukemia Cell Growth
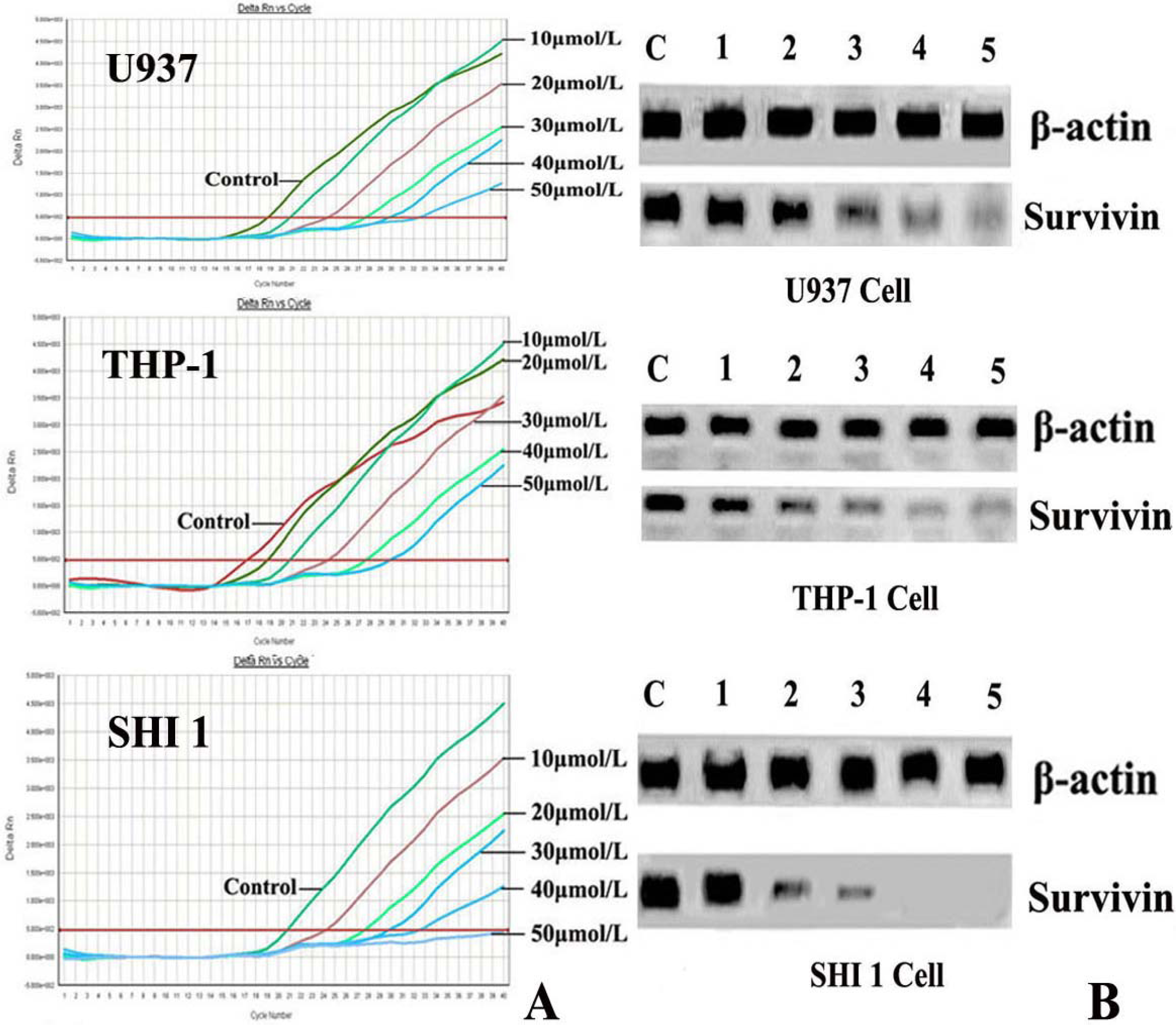
3.2. Tan-I Inhibits Survivin Expression
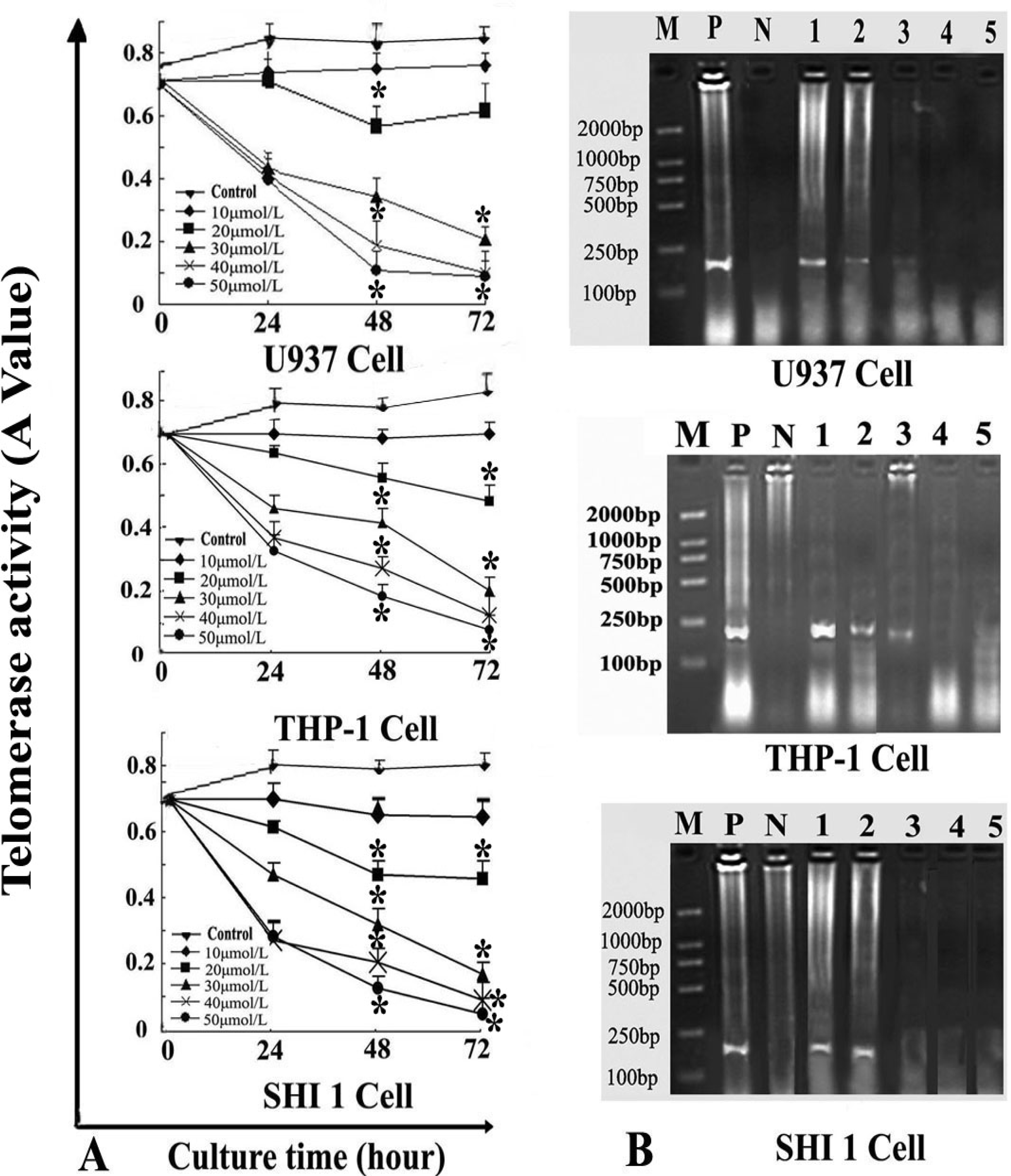
3.3. Tan-I Inhibits Telomerase Activity and hTERT mRNA Expression
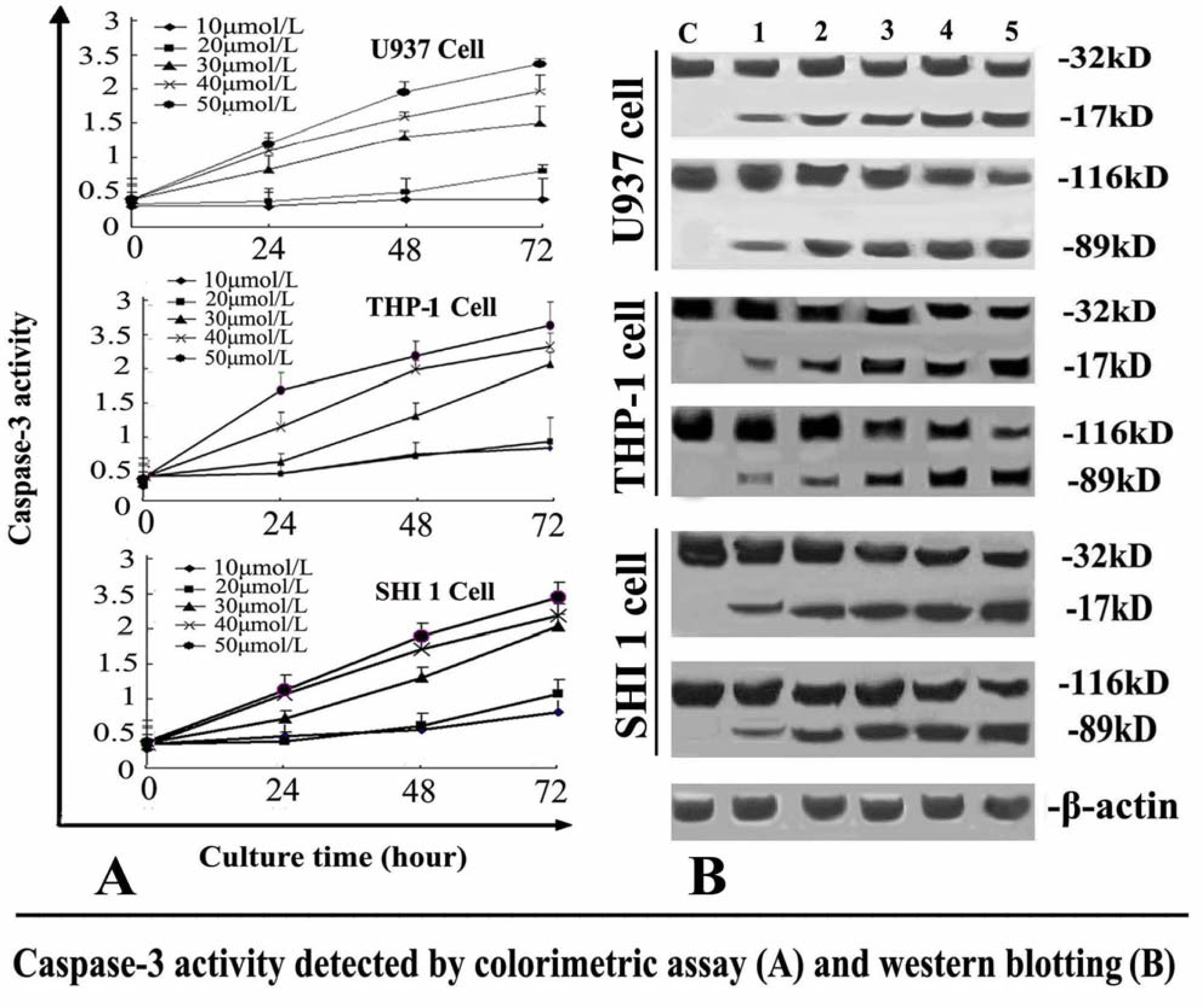
3.4. Tan-I Activates Caspase-3
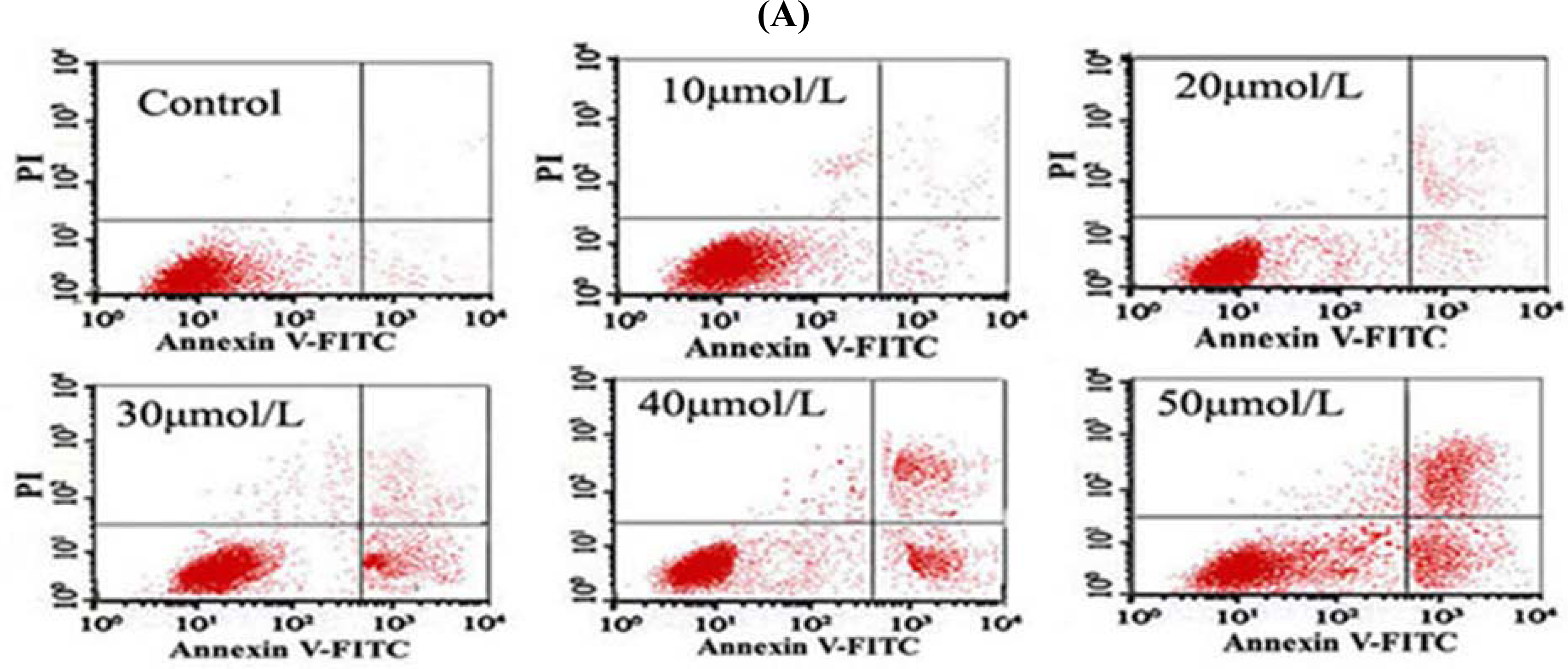
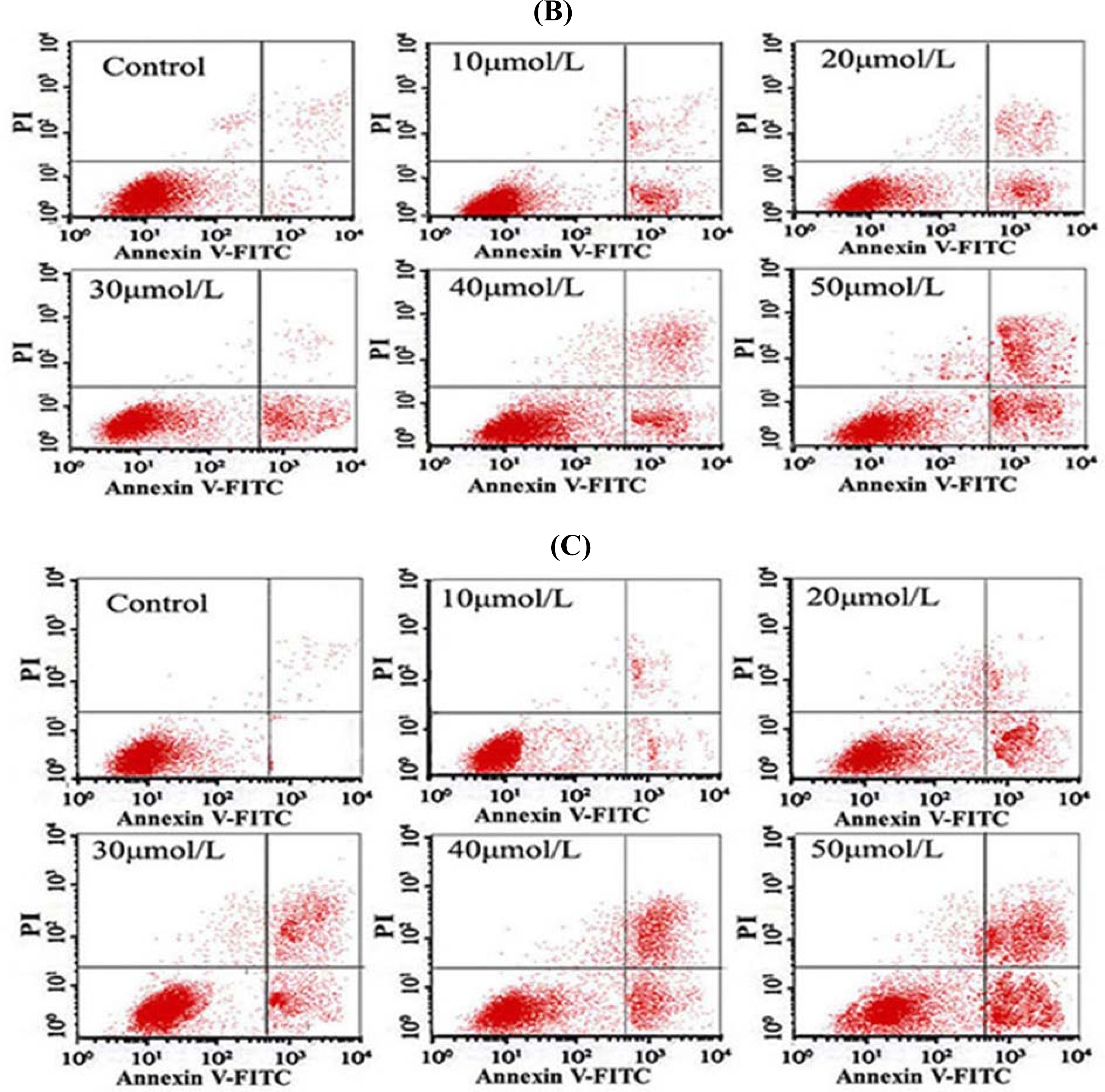
3.5. Tan-I Promotes Apoptosis in Leukemia Cells
4. Discussion
5. Conclusion
Acknowledgments
References
- Boon, H; Wong, J. Botanical medicine and cancer: A review of the safety and efficacy. Exp. Opin. Pharmacother 2004, 5, 2485–2501. [Google Scholar]
- Zhou, L; Zuo, Z; Chow, MS. Danshen: An overview of its chemistry, pharmacology, pharmacokinetics, and clinical use. J. Clin. Pharmacol 2005, 45, 1345–1359. [Google Scholar]
- Wang, AM; Sha, SH; Lesniak, W; Schacht, J. Tanshinone (Salviae miltiorrhizae extract) preparations attenuate aminoglycoside-induced free radical formation in vitro and ototoxicity in vivo. Antimicrob. Agents Chemother 2003, 47, 1836–1841. [Google Scholar]
- Xu, WX; Li, JF; Dong, C. Spectral Properties of Tanshinone I and pH Effect on Its Molecular Structure. Chin. J. Spectro. Lab 2003, 20, 319–323. [Google Scholar]
- Su, CC; Chen, GW; Lin, JG. Growth inhibition and apoptosis induction by tanshinone I in human colon cancer Colo 205 cells. Int. J. Mol. Med 2008, 22, 613–618. [Google Scholar]
- Lee, CY; Sher, HF; Chen, HW; Liu, CC; Chen, CH; Lin, CS; Yang, PC; Tsay, HS; Chen, JJ. Anticancer effects of tanshinone I in human non-small cell lung cancer. Mol. Cancer Ther 2008, 7, 3527–3538. [Google Scholar]
- Nizamutdinova, IT; Lee, GW; Son, KH; Jeon, SJ; Kang, SS; Kim, YS; Lee, JH; Seo, HG; Chang, KC; Kim, HJ. Tanshinone I effectively induces apoptosis in estrogen receptor-positive (MCF-7) and estrogen receptor-negative (MDA-MB-231) breast cancer cells. Int. J. Oncol 2008, 33, 485–491. [Google Scholar]
- Debatin, KM. Apoptosis pathways in cancer and cancer therapy. Cancer. Immunol. Immunother 2004, 53, 153–197. [Google Scholar]
- Rossi, D; Gaidano, G. Messengers of cell death: apoptotic signaling in health and disease. Haematologica 2003, 88, 212–218. [Google Scholar]
- Tsuruo, T; Naito, M; Tomida, A; Fujita, N; Mashima, T; Sakamoto, H; Haga, N. Molecular targeting therapy of cancer: drug resistance, apoptosis and survival signal. Cancer Sci 2003, 94, 15–21. [Google Scholar]
- Ryan, BM; O’Donovan, N; Duffy, MJ. Survivin: A new target for anti-cancer therapy. Cancer Treat Rev 2009, 35, 553–562. [Google Scholar]
- Kanwa, RK; Cheung, CH; Chang, JY; Kanwar, JR. Recent advances in anti-survivin treatments for cancer. Curr. Med. Chem 2010, 17, 1509–1515. [Google Scholar]
- Oto, OA; Paydas, S; Tanriverdi, K; Seydaoglu, G; Yavuz, S; Disel, U. Survivin and EPR-1 expression in acute leukemias: prognostic significance and review of the literature. Leuk. Res 2007, 31, 1495–1501. [Google Scholar]
- Lin, KW; Yan, J. The telomere length dynamic and methods of its assessment. J. Cell Mol. Med 2005, 9, 977–989. [Google Scholar]
- Parkinson, EK. Telomerase as a novel and potentially selective target for cancer chemotherapy. Ann. Med 2003, 35, 466–475. [Google Scholar]
- Mokbel, K. The evolving role of telomerase inhibitors in the treatment of cancer. Curr. Med. Res. Opin 2003, 19, 470–472. [Google Scholar]
- Cosme-Blanco, W; Chang, S. Dual roles of telomere dysfunction in initiation and suppression of tumorigenesis. Exp. Cell Res 2008, 314, 1973–1979. [Google Scholar]
- Tauchi, T; Shin-ya, K; Sashida, G; Sumi, M; Okabe, S; Ohyashiki, JH; Ohyashiki, K. Telomerase inhibition with a novel G-quadruplex-interactive agent, telomestatin: In vitro and in vivo studies in acute leukemia. Oncogene 2006, 25, 5719–5725. [Google Scholar]
- Nakajima, A; Tauchi, T; Sashida, G; Sumi, M; Abe, K; Yamamoto, K; Ohyashiki, JH; Ohyashiki, K. Telomerase inhibition enhances apoptosis in human acute leukemia cells: Possibility of antitelomerase therapy. Leukemia 2003, 17, 560–567. [Google Scholar]
- Shammas, MA; Koley, H; Bertheau, RC; Neri, P; Fulciniti, M; Tassone, P; Blotta, S; Protopopov, A; Mitsiades, C; Batchu, RB; Anderson, KC; Chin, A; Gryaznov, S; Munshi, NC. Telomerase inhibitor GRN163L inhibits myeloma cell growth in vitro and in vivo. Leukemia 2008, 22, 1410–1418. [Google Scholar]
- Parsch, D; Brassat, U; Brummendorf, TH; Fellenberg, J. Consequences of telomerase inhibition by BIBR1532 on proliferation and chemosensitivity of chondrosarcoma cell lines. Cancer Invest 2008, 26, 590–596. [Google Scholar]
- Yang, SM; Fang, DC; Yang, JL; Chen, L; Luo, YH; Liang, GP. Antisense human telomerase reverse transcriptase could partially reverse malignant phenotypes of gastric carcinoma cell line in vitro. Eur. J. Cancer Prev 2008, 17, 209–217. [Google Scholar]
- Moorhouse, AD; Haider, S; Gunaratnam, M; Munnur, D; Neidle, S; Moses, JE. Targeting telomerase and telomeres: A click chemistry approach towards highly selective G-quadruplex ligands. Mol. Biosyst 2008, 4, 629–642. [Google Scholar]







© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, X.-D.; Fan, R.-F.; Zhang, Y.; Yang, H.-Z.; Fang, Z.-G.; Guan, W.-B.; Lin, D.-J.; Xiao, R.-Z.; Huang, R.-W.; Huang, H.-Q.; et al. Down-Regulation of Telomerase Activity and Activation of Caspase-3 Are Responsible for Tanshinone I-Induced Apoptosis in Monocyte Leukemia Cells in Vitro. Int. J. Mol. Sci. 2010, 11, 2267-2280. https://doi.org/10.3390/ijms11062267
Liu X-D, Fan R-F, Zhang Y, Yang H-Z, Fang Z-G, Guan W-B, Lin D-J, Xiao R-Z, Huang R-W, Huang H-Q, et al. Down-Regulation of Telomerase Activity and Activation of Caspase-3 Are Responsible for Tanshinone I-Induced Apoptosis in Monocyte Leukemia Cells in Vitro. International Journal of Molecular Sciences. 2010; 11(6):2267-2280. https://doi.org/10.3390/ijms11062267
Chicago/Turabian StyleLiu, Xiao-Dan, Rui-Fang Fan, Yong Zhang, Hong-Zhi Yang, Zhi-Gang Fang, Wei-Bing Guan, Dong-Jun Lin, Ruo-Zhi Xiao, Ren-Wei Huang, He-Qing Huang, and et al. 2010. "Down-Regulation of Telomerase Activity and Activation of Caspase-3 Are Responsible for Tanshinone I-Induced Apoptosis in Monocyte Leukemia Cells in Vitro" International Journal of Molecular Sciences 11, no. 6: 2267-2280. https://doi.org/10.3390/ijms11062267
APA StyleLiu, X.-D., Fan, R.-F., Zhang, Y., Yang, H.-Z., Fang, Z.-G., Guan, W.-B., Lin, D.-J., Xiao, R.-Z., Huang, R.-W., Huang, H.-Q., Liu, P.-Q., & Liu, J.-J. (2010). Down-Regulation of Telomerase Activity and Activation of Caspase-3 Are Responsible for Tanshinone I-Induced Apoptosis in Monocyte Leukemia Cells in Vitro. International Journal of Molecular Sciences, 11(6), 2267-2280. https://doi.org/10.3390/ijms11062267