Diversity of Nonribosomal Peptide Synthetases Involved in the Biosynthesis of Lipopeptide Biosurfactants


Abstract
:1. Introduction
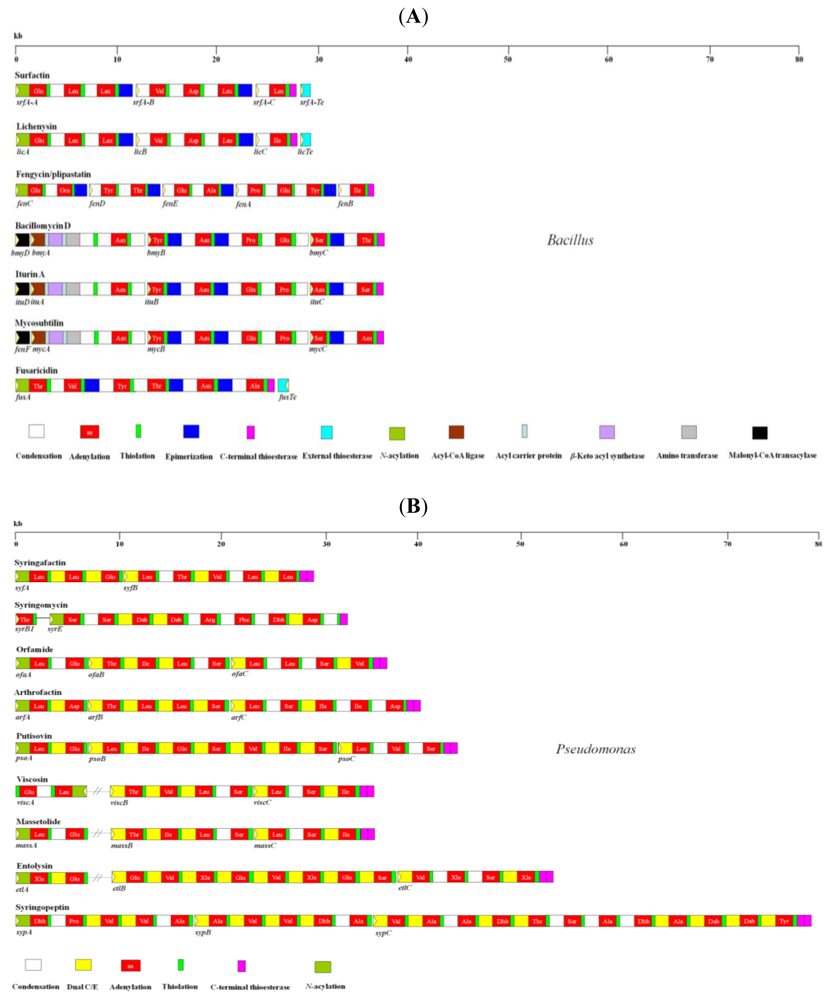
2. Diversity of LPBSs and NRPSs from Bacillus and Pseudomonas
2.1. Bacillus
2.1.1. Surfactin and Lichenysin Synthetases
2.1.2. Fengycin Synthetase
2.1.3. Bacillomycin, Iturin, and Mycosubtilin Synthetases
2.1.4. Fusaricidin Synthetase
2.2. Pseudomonas
2.2.1. Syringomycin Synthetase
2.2.2. Syringopeptin Syntheatase
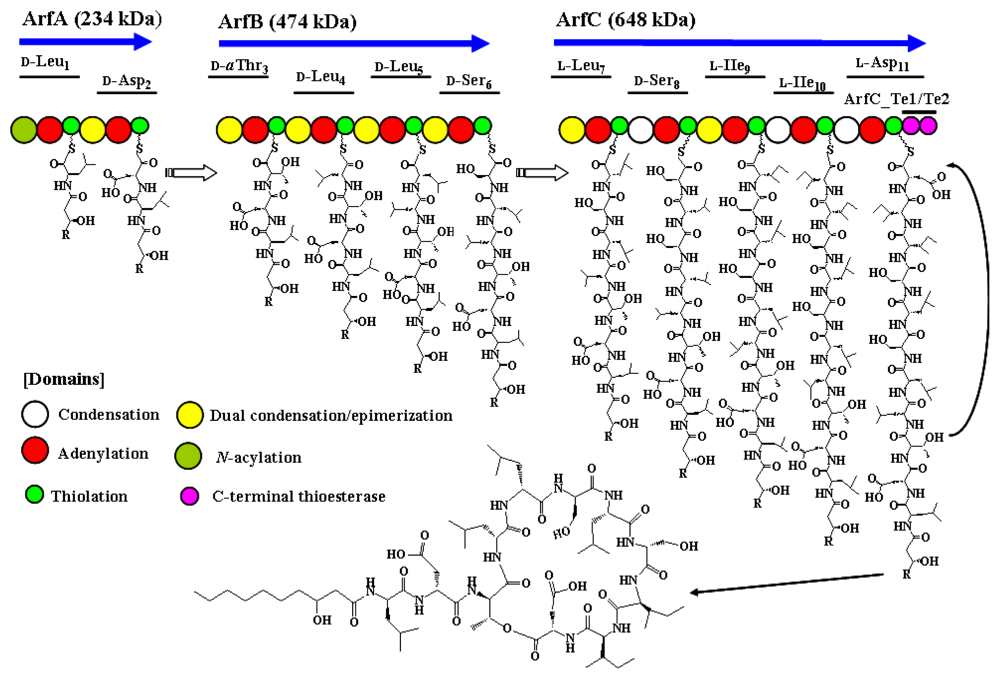
2.2.3. Arthrofactin Synthetase
2.2.4. Viscosin and Massetolide Synthetases
2.2.5. Orfamide Synthetase
2.2.6. Putisolvin Synthetase
2.2.7. Syringafactin Synthetase
2.2.8. Entolysin Synthetase
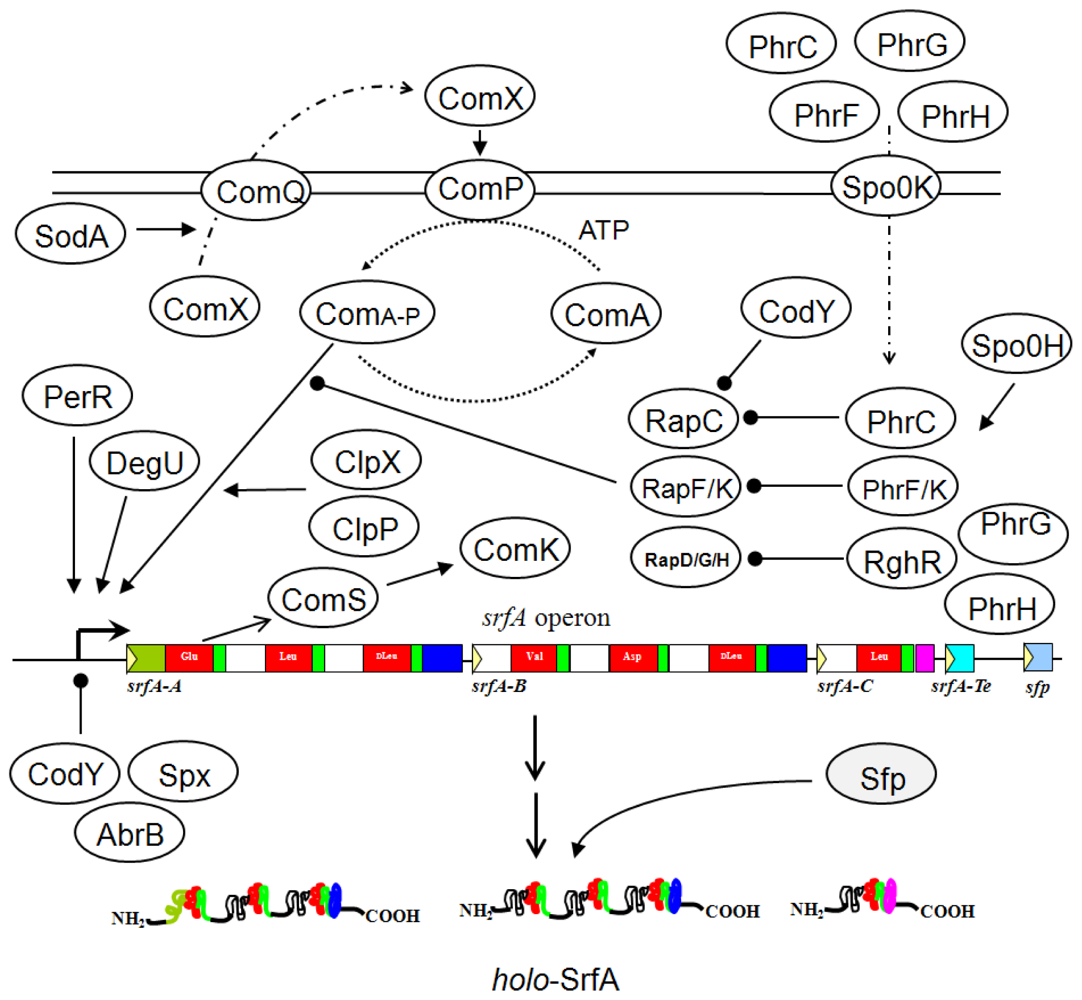
3. Gene Regulation in Bacillus
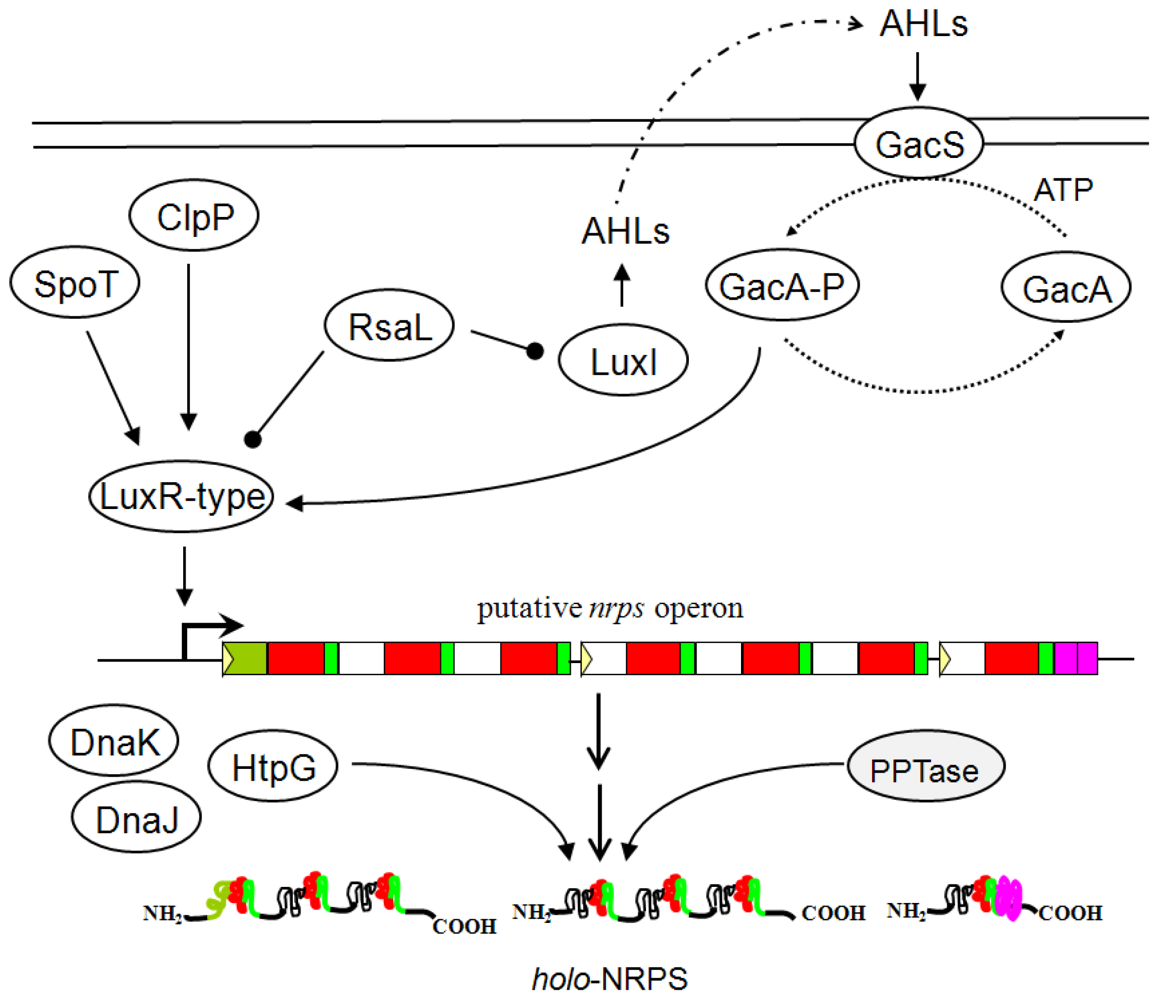
4. Gene Regulation in Pseudomonas
5. Genetic Engineering of NRPS to Create Novel Products
6. Conclusion
Acknowledgements
References
- Georgiou, G; Lin, SC; Sharma, MM. Surface-active compounds from microorganisms. Biotechnology (NY) 1992, 10, 60–65. [Google Scholar]
- Desai, JD; Banat, IM. Microbial production of surfactants and their commercial potential. Microbiol. Mol. Biol. Rev 1997, 61, 47–64. [Google Scholar]
- Arima, K; Kakinuma, A; Tamura, G. Surfactin, a crystalline peptidelipid surfactant produced by Bacillus subtilis: isolation, characterization and its inhibition of fibrin clot formation. Biochem. Biophys. Res. Commun 1968, 31, 488–494. [Google Scholar]
- Nishikiori, T; Naganawa, H; Muraoka, Y; Aoyagi, T; Umezawa, H. Plipastatins: new inhibitors of phospholipase A2, produced by Bacillus cereus BMG302-fF67. III. Structural elucidation of plipastatins. J. Antibiot (Tokyo) 1986, 39, 755–761. [Google Scholar]
- Peypoux, F; Pommier, MT; Das, BC; Besson, F; Delcambe, L; Michel, G. Structures of bacillomycin D and bacillomycin L peptidolipid antibiotics from Bacillus subtilis. J. Antibiot (Tokyo) 1984, 37, 1600–1604. [Google Scholar]
- Grangemard, I; Bonmatin, JM; Bernillon, J; Das, BC; Peypoux, F. Lichenysins G, a novel family of lipopeptide biosurfactants from Bacillus licheniformis IM 1307: production, isolation and structural evaluation by NMR and mass spectrometry. J. Antibiot (Tokyo) 1999, 52, 363–373. [Google Scholar]
- Yakimov, MM; Abraham, WR; Meyer, H; Laura, G; Golyshin, PN. Structural characterization of lichenysin A components by fast atom bombardment tandem mass spectrometry. Biochim. Biophys Acta 1999, 1438, 273–280. [Google Scholar]
- Morikawa, M; Hirata, Y; Imanaka, T. A study on the structure-function relationship of lipopeptide biosurfactants. Biochim. Biophys Acta 2000, 1488, 211–218. [Google Scholar]
- Nielsen, TH; Sorensen, D; Tobiasen, C; Andersen, JB; Christophersen, C; Givskov, M; Sorensen, J. Antibiotic and biosurfactant properties of cyclic lipopeptides produced by fluorescent Pseudomonas spp. from the sugar beet rhizosphere. Appl. Environ. Microbiol 2002, 68, 3416–3423. [Google Scholar]
- Sorensen, D; Nielsen, TH; Christophersen, C; Sorensen, J; Gajhede, M. Cyclic lipoundecapeptide amphisin from Pseudomonas sp. strain DSS73. Acta Crystallogr C 2001, 57, 1123–1124. [Google Scholar]
- Sorensen, D; Nielsen, TH; Sorensen, J; Christophersen, C. Cyclic lipoundecapeptide lokisin from Pseudomonas sp. strain DSS41. Tetrahedron Lett 2002, 43, 4421–4423. [Google Scholar]
- Ui, H; Miyake, T; Iinuma, H; Imoto, M; Naganawa, H; Hattori, S; Hamada, M; Takeuchi, T; Umezawa, S; Umezawa, K. Pholipeptin, a novel cyclic lipoundecapeptide from Pseudomonas fluorescens. J. Org. Chem 1997, 62, 103–108. [Google Scholar]
- Eppelmann, K; Stachelhaus, T; Marahiel, MA. Exploitation of the selectivity-conferring code of nonribosomal peptide synthetases for the rational design of novel peptide antibiotics. Biochemistry 2002, 41, 9718–9726. [Google Scholar]
- Stachelhaus, T; Schneider, A; Marahiel, MA. Rational design of peptide antibiotics by targeted replacement of bacterial and fungal domains. Science 1995, 269, 69–72. [Google Scholar]
- Koglin, A; Walsh, CT. Structural insights into nonribosomal peptide enzymatic assembly lines. Nat. Prod. Rep 2009, 26, 987–1000. [Google Scholar]
- Raaijmakers, JM; de Bruijn, I; Nybroe, O; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol Rev 2010. [Google Scholar] [CrossRef]
- Roongsawang, N; Thaniyavarn, J; Thaniyavarn, S; Kameyama, T; Haruki, M; Imanaka, T; Morikawa, M; Kanaya, S. Isolation and characterization of a halotolerant Bacillus subtilis BBK-1 which produces three kinds of lipopeptides: bacillomycin L, plipastatin, and surfactin. Extremophiles 2002, 6, 499–506. [Google Scholar]
- Koumoutsi, A; Chen, XH; Henne, A; Liesegang, H; Hitzeroth, G; Franke, P; Vater, J; Borriss, R. Structural and functional characterization of gene clusters directing nonribosomal synthesis of bioactive cyclic lipopeptides in Bacillus amyloliquefaciens strain FZB42. J. Bacteriol 2004, 186, 1084–1096. [Google Scholar]
- Romero, D; de Vicente, A; Rakotoaly, RH; Dufour, SE; Veening, JW; Arrebola, E; Cazorla, FM; Kuipers, OP; Paquot, M; Perez-Garcia, A. The iturin and fengycin families of lipopeptides are key factors in antagonism of Bacillus subtilis toward Podosphaera fusca. Mol. Plant Microbe Interact 2007, 20, 430–440. [Google Scholar]
- Kim, PI; Ryu, J; Kim, YH; Chi, YT. Production of biosurfactant lipopeptides iturin A, fengycin and surfactin A from Bacillus subtilis CMB32 for control of Colletotrichum gloeosporioides. J. Microbiol. Biotechnol 2010, 20, 138–145. [Google Scholar]
- Yakimov, MM; Timmis, KN; Wray, V; Fredrickson, HL. Characterization of a new lipopeptide surfactant produced by thermotolerant and halotolerant subsurface Bacillus licheniformis BAS50. Appl. Environ. Microbiol 1995, 61, 1706–1713. [Google Scholar]
- Bonmatin, JM; Laprevote, O; Peypoux, F. Diversity among microbial cyclic lipopeptides: iturins and surfactins. Activity-structure relationships to design new bioactive agents. Comb. Chem High Throughput Screen 2003, 6, 541–556. [Google Scholar]
- Grangemard, I; Wallach, J; Maget-Dana, R; Peypoux, F. Lichenysin: a more efficient cation chelator than surfactin. Appl. Biochem. Biotechnol 2001, 90, 199–210. [Google Scholar]
- Rodrigues, L; Banat, IM; Teixeira, J; Oliveira, R. Biosurfactants: potential applications in medicine. J. Antimicrob. Chemother 2006, 57, 609–618. [Google Scholar] [Green Version]
- Bais, HP; Fall, R; Vivanco, JM. Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiol 2004, 134, 307–319. [Google Scholar]
- Hofemeister, J; Conrad, B; Adler, B; Hofemeister, B; Feesche, J; Kucheryava, N; Steinborn, G; Franke, P; Grammel, N; Zwintscher, A; Leenders, F; Hitzeroth, G; Vater, J. Genetic analysis of the biosynthesis of nonribosomal peptide and polyketide-like antibiotics, iron uptake and biofilm formation by Bacillus subtilis A1/3. Mol. Genet Genomics 2004, 272, 363–378. [Google Scholar]
- Kearns, DB; Losick, R. Swarming motility in undomesticated Bacillus subtilis. Mol. Microbiol 2003, 49, 581–590. [Google Scholar]
- Julkowska, D; Obuchowski, M; Holland, IB; Seror, SJ. Comparative analysis of the development of swarming communities of Bacillus subtilis 168 and a natural wild type: critical effects of surfactin and the composition of the medium. J. Bacteriol 2005, 187, 65–76. [Google Scholar]
- Branda, SS; Gonzalez-Pastor, JE; Ben-Yehuda, S; Losick, R; Kolter, R. Fruiting body formation by Bacillus subtilis. Proc. Natl. Acad. Sci USA 2001, 98, 11621–11626. [Google Scholar]
- Mireles, JR; Toguchi, A; Harshey, RM. Salmonella enterica serovar typhimurium swarming mutants with altered biofilm-forming abilities: surfactin inhibits biofilm formation. J. Bacteriol 2001, 183, 5848–5854. [Google Scholar]
- Kakinuma, A; Ouchida, A; Shima, T; Sugino, H; Isono, M; Tamura, G; Arima, K. Conformation of the structure of surfactin by mass spectrometry. Agric. Biol. Chem 1969, 33, 1669–1671. [Google Scholar]
- Konz, D; Doekel, S; Marahiel, MA. Molecular and biochemical characterization of the protein template controlling biosynthesis of the lipopeptide lichenysin. J. Bacteriol 1999, 181, 133–140. [Google Scholar]
- Lin, SC; Minton, MA; Sharma, MM; Georgiou, G. Structural and immunological characterization of a biosurfactant produced by Bacillus licheniformis JF-2. Appl. Environ. Microbiol 1994, 60, 31–38. [Google Scholar]
- Jenny, K; Kappeli, O; Fiechter, A. Biosurfactants from Bacillus licheniformis: structural analysis and characterization. Appl. Microbiol. Biotechnol 1991, 36, 5–13. [Google Scholar]
- Horowitz, S; Gilbert, JN; Griffin, WM. Isolation and characterization of a surfactant produced by Bacillus licheniformis 86. J. Indus. Microbiol 1990, 6, 243–248. [Google Scholar]
- Naruse, N; Tenmyo, O; Kobaru, S; Kamei, H; Miyaki, T; Konishi, M; Oki, T. Pumilacidin, a complex of new antiviral antibiotics. Production, isolation, chemical properties, structure and biological activity. J Antibiot (Tokyo) 1990, 43, 267–280. [Google Scholar]
- Volpon, L; Besson, F; Lancelin, JM. NMR structure of antibiotics plipastatins A and B from Bacillus subtilis inhibitors of phospholipase A(2). FEBS Lett 2000, 485, 76–80. [Google Scholar]
- Isogai, I; Takayama, S; Murakoshi, S; Suzuki, A. Structure of β-amino acids in antibiotics iturin A. Tetrahedron Lett 1982, 23, 3065–3068. [Google Scholar]
- Volpon, L; Tsan, P; Majer, Z; Vass, E; Hollosi, M; Noguera, V; Lancelin, JM; Besson, F. NMR structure determination of a synthetic analogue of bacillomycin Lc reveals the strategic role of L-Asn1 in the natural iturinic antibiotics. Spectrochim Acta Part A 2007, 67, 1374–1381. [Google Scholar]
- Duitman, EH; Hamoen, LW; Rembold, M; Venema, G; Seitz, H; Saenger, W; Bernhard, F; Reinhardt, R; Schmidt, M; Ullrich, C; Stein, T; Leenders, F; Vater, J. The mycosubtilin synthetase of Bacillus subtilis ATCC6633: a multifunctional hybrid between a peptide synthetase, an amino transferase, and a fatty acid synthase. Proc. Natl. Acad. Sci USA 1999, 96, 13294–13299. [Google Scholar]
- Batrakov, SG; Rodionova, TA; Esipov, SE; Polyakov, NB; Sheichenko, VI; Shekhovtsova, NV; Lukin, SM; Panikov, NS; Nikolaev, YA. A novel lipopeptide, an inhibitor of bacterial adhesion, from the thermophilic and halotolerant subsurface Bacillus licheniformis strain 603. Biochim. Biophys Acta 2003, 1634, 107–115. [Google Scholar]
- Lee, SC; Kim, SH; Park, IH; Chung, SY; Choi, YL. Isolation and structural analysis of bamylocin A, novel lipopeptide from Bacillus amyloliquefaciens LP03 having antagonistic and crude oil-emulsifying activity. Arch. Microbiol 2007, 188, 307–312. [Google Scholar]
- He, H; Shen, B; Korshalla, J; Carter, GT. Circulocins, new antibacterial lipopeptides from Bacillus circulans, J2154. Tetrahedron 2001, 57, 1189–1195. [Google Scholar]
- Li, J; Jensen, SE. Nonribosomal biosynthesis of fusaricidins by Paenibacillus polymyxa PKB1 involves direct activation of a D-amino acid. Chem. Biol 2008, 15, 118–127. [Google Scholar]
- Hathout, Y; Ho, YP; Ryzhov, V; Demirev, P; Fenselau, C. Kurstakins: a new class of lipopeptides isolated from Bacillus thuringiensis. J. Nat. Prod 2000, 63, 1492–1496. [Google Scholar]
- de Bruijn, I; de Kock, MJ; Yang, M; de Waard, P; van Beek, TA; Raaijmakers, JM. Genome-based discovery, structure prediction and functional analysis of cyclic lipopeptide antibiotics in Pseudomonas species. Mol. Microbiol 2007, 63, 417–428. [Google Scholar]
- Nielsen, TH; Christophersen, C; Anthoni, U; Sorensen, J. Viscosinamide, a new cyclic depsipeptide with surfactant and antifungal properties produced by Pseudomonas fluorescens DR54. J. Appl. Microbiol 1999, 87, 80–90. [Google Scholar]
- de Bruijn, I; de Kock, MJ; de Waard, P; van Beek, TA; Raaijmakers, JM. Massetolide A biosynthesis in Pseudomonas fluorescens. J. Bacteriol 2008, 190, 2777–2789. [Google Scholar]
- Quail, JW; Ismail, N; Pedras, MS; Boyetchko, SM. Pseudophomins A and B, a class of cyclic lipodepsipeptides isolated from a Pseudomonas species. Acta Crystallogr C 2002, 58, 268–271. [Google Scholar]
- Sinnaeve, D; Hendrickx, PM; Van Hemel, J; Peys, E; Kieffer, B; Martins, JC. The solution structure and self-association properties of the cyclic lipodepsipeptide pseudodesmin A support its pore-forming potential. Chemistry 2009, 15, 12653–12662. [Google Scholar]
- Guenzi, E; Galli, G; Grgurina, I; Gross, DC; Grandi, G. Characterization of the syringomycin synthetase gene cluster. A link between prokaryotic and eukaryotic peptide synthetases. J. Biol. Chem 1998, 273, 32857–32863. [Google Scholar]
- Sorensen, KN; Kim, KH; Takemoto, JY. In vitro antifungal and fungicidal activities and erythrocyte toxicities of cyclic lipodepsinonapeptides produced by Pseudomonas syringae pv. syringae. Antimicrob. Agents Chemother 1996, 40, 2710–2713. [Google Scholar]
- Ballio, A; Bossa, F; Collina, A; Gallo, M; Iacobellis, NS; Paci, M; Pucci, P; Scaloni, A; Segre, A; Simmaco, M. Structure of syringotoxin, a bioactive metabolite of Pseudomonas syringae pv. syringae. FEBS Lett 1990, 269, 377–380. [Google Scholar]
- Harrison, L; Teplow, DB; Rinaldi, M; Strobel, G. Pseudomycins, a family of novel peptides from Pseudomonas syringae possessing broad-spectrum antifungal activity. J. Gen. Microbiol 1991, 137, 2857–2865. [Google Scholar]
- Scaloni, A; Dalla Serra, M; Amodeo, P; Mannina, L; Vitale, RM; Segre, AL; Cruciani, O; Lodovichetti, F; Greco, ML; Fiore, A; Gallo, M; D’Ambrosio, C; Coraiola, M; Menestrina, G; Graniti, A; Fogliano, V. Structure, conformation and biological activity of a novel lipodepsipeptide from Pseudomonas corrugata: cormycin A. Biochem. J 2004, 384, 25–36. [Google Scholar]
- Morikawa, M; Daido, H; Takao, T; Murata, S; Shimonishi, Y; Imanaka, T. A new lipopeptide biosurfactant produced by Arthrobacter sp. strain MIS38. J. Bacteriol 1993, 175, 6459–6466. [Google Scholar]
- Henriksen, A; Anthoni, U; Nielsen, TH; Sorensen, J; Christophersen, C; Gajhede, M. Cyclic lipoundecapeptide tensin from Pseudomonas fluorescens strain 96.578. Acta Crystallogr C 2000, 56 Pt 1, 113–115. [Google Scholar]
- Kuiper, I; Lagendijk, EL; Pickford, R; Derrick, JP; Lamers, GE; Thomas-Oates, JE; Lugtenberg, BJ; Bloemberg, GV. Characterization of two Pseudomonas putida lipopeptide biosurfactants, putisolvin I and II, which inhibit biofilm formation and break down existing biofilms. Mol. Microbiol 2004, 51, 97–113. [Google Scholar]
- Bassarello, C; Lazzaroni, S; Bifulco, G; Lo Cantore, P; Iacobellis, NS; Riccio, R; Gomez- Paloma, L; Evidente, A. Tolaasins A-E, five new lipodepsipeptides produced by Pseudomonas tolaasii. J. Nat. Prod 2004, 67, 811–816. [Google Scholar]
- Ballio, A; Bossa, F; Camoni, L; Di Giorgio, D; Flamand, MC; Maraite, H; Nitti, G; Pucci, P; Scaloni, A. Structure of fuscopeptins, phytotoxic metabolites of Pseudomonas fuscovaginae. FEBS Lett 1996, 381, 213–216. [Google Scholar]
- Emanuele, MC; Scaloni, A; Lavermicocca, P; Jacobellis, NS; Camoni, L; Di Giorgio, D; Pucci, P; Paci, M; Segre, A; Ballio, A. Corpeptins, new bioactive lipodepsipeptides from cultures of Pseudomonas corrugata. FEBS Lett 1998, 433, 317–320. [Google Scholar]
- Ballio, A; Barra, D; Bossa, F; Collina, A; Grgurina, I; Marino, G; Moneti, G; Paci, M; Pucci, P; Segre, A; et al. Syringopeptins, new phytotoxic lipodepsipeptides of Pseudomonas syringae pv. syringae. FEBS Lett 1991, 291, 109–112. [Google Scholar]
- Ballio, A; Bossa, F; Di Giorgio, D; Di Nola, A; Manetti, C; Paci, M; Scaloni, A; Segre, AL. Solution conformation of the Pseudomonas syringae pv. syringae phytotoxic lipodepsipeptide syringopeptin 25-A. Two-dimensional NMR, distance geometry and molecular dynamics. Eur. J. Biochem 1995, 234, 747–758. [Google Scholar]
- Scaloni, A; Camoni, L; di Giorgio, D; Scortichini, M; Cozzolino, R; Ballio, A. A new syringopeptin produced by a Pseudomonas syringae pv syringae strain isolated from diseased twigs of laurel. Physiol. Mol. Plant Pathol 1997, 51, 259–264. [Google Scholar]
- Vallet-Gely, I; Novikov, A; Augusto, L; Liehl, P; Bolbach, G; Pechy-Tarr, M; Cosson, P; Keel, C; Caroff, M; Lemaitre, B. Association of hemolytic activity of Pseudomonas entomophila, a versatile soil bacterium, with cyclic lipopeptide production. Appl. Environ. Microbiol 2010, 76, 910–921. [Google Scholar]
- Gross, H; Stockwell, VO; Henkels, MD; Nowak-Thompson, B; Loper, JE; Gerwick, WH. The genomisotopic approach: a systematic method to isolate products of orphan biosynthetic gene clusters. Chem. Biol 2007, 14, 53–63. [Google Scholar]
- Janek, T; Lukaszewicz, M; Rezanka, T; Krasowska, A. Isolation and characterization of two new lipopeptide biosurfactants produced by Pseudomonas fluorescens BD5 isolated from water from the Arctic Archipelago of Svalbard. Bioresour. Technol 2010, 101, 6118–6123. [Google Scholar]
- Berti, AD; Greve, NJ; Christensen, QH; Thomas, MG. Identification of a biosynthetic gene cluster and the six associated lipopeptides involved in swarming motility of Pseudomonas syringae pv. tomato DC3000. J. Bacteriol 2007, 189, 6312–6323. [Google Scholar]
- Cosmina, P; Rodriguez, F; de Ferra, F; Grandi, G; Perego, M; Venema, G; van Sinderen, D. Sequence and analysis of the genetic locus responsible for surfactin synthesis in Bacillus subtilis. Mol. Microbiol 1993, 8, 821–831. [Google Scholar]
- Roongsawang, N; Lim, SP; Washio, K; Takano, K; Kanaya, S; Morikawa, M. Phylogenetic analysis of condensation domains in the nonribosomal peptide synthetases. FEMS Microbiol. Lett 2005, 252, 143–151. [Google Scholar]
- Kraas, FI; Helmetag, V; Wittmann, M; Strieker, M; Marahiel, MA. Functional dissection of surfactin synthetase initiation module reveals insights into the mechanism of lipoinitiation. Chem. Biol 2010, 17, 872–880. [Google Scholar]
- Schneider, A; Marahiel, MA. Genetic evidence for a role of thioesterase domains, integrated in or associated with peptide synthetases, in non-ribosomal peptide biosynthesis in Bacillus subtilis. Arch. Microbiol 1998, 169, 404–410. [Google Scholar]
- Schwarzer, D; Mootz, HD; Linne, U; Marahiel, MA. Regeneration of misprimed nonribosomal peptide synthetases by type II thioesterases. Proc. Natl. Acad. Sci USA 2002, 99, 14083–14088. [Google Scholar]
- Yeh, E; Kohli, RM; Bruner, SD; Walsh, CT. Type II thioesterase restores activity of a NRPS module stalled with an aminoacyl-S-enzyme that cannot be elongated. ChemBioChem 2004, 5, 1290–1293. [Google Scholar]
- Menkhaus, M; Ullrich, C; Kluge, B; Vater, J; Vollenbroich, D; Kamp, RM. Structural and functional organization of the surfactin synthetase multienzyme system. J. Biol. Chem 1993, 268, 7678–7684. [Google Scholar]
- Steller, S; Sokoll, A; Wilde, C; Bernhard, F; Franke, P; Vater, J. Initiation of surfactin biosynthesis and the role of the SrfD-thioesterase protein. Biochemistry 2004, 43, 11331–11343. [Google Scholar]
- Vanittanakom, N; Loeffler, W; Koch, U; Jung, G. Fengycin-a novel antifungal lipopeptide antibiotic produced by Bacillus subtilis F-29-3. J. Antibiot (Tokyo) 1986, 39, 888–901. [Google Scholar]
- Rivardo, F; Turner, RJ; Allegrone, G; Ceri, H; Martinotti, MG. Anti-adhesion activity of two biosurfactants produced by Bacillus spp. prevents biofilm formation of human bacterial pathogens. Appl. Microbiol. Biotechnol 2009, 83, 541–553. [Google Scholar]
- Thaniyavarn, J; Roongsawang, N; Kameyama, T; Haruki, M; Imanaka, T; Morikawa, M; Kanaya, S. Production and characterization of biosurfactants from Bacillus licheniformis F2.2. Biosci. Biotechnol. Biochem 2003, 67, 1239–1244. [Google Scholar]
- Wu, CY; Chen, CL; Lee, YH; Cheng, YC; Wu, YC; Shu, HY; Gotz, F; Liu, ST. Nonribosomal synthesis of fengycin on an enzyme complex formed by fengycin synthetases. J. Biol. Chem 2007, 282, 5608–5616. [Google Scholar]
- Leclere, V; Marti, R; Bechet, M; Fickers, P; Jacques, P. The lipopeptides mycosubtilin and surfactin enhance spreading of Bacillus subtilis strains by their surface-active properties. Arch. Microbiol 2006, 186, 475–483. [Google Scholar]
- Tsuge, K; Akiyama, T; Shoda, M. Cloning, sequencing, and characterization of the iturin A operon. J. Bacteriol 2001, 183, 6265–6273. [Google Scholar]
- Moyne, AL; Cleveland, TE; Tuzun, S. Molecular characterization and analysis of the operon encoding the antifungal lipopeptide bacillomycin D. FEMS Microbiol. Lett 2004, 234, 43–49. [Google Scholar]
- Hansen, DB; Bumpus, SB; Aron, ZD; Kelleher, NL; Walsh, CT. The loading module of mycosubtilin: an adenylation domain with fatty acid selectivity. J. Am. Chem. Soc 2007, 129, 6366–6367. [Google Scholar]
- Aron, ZD; Fortin, PD; Calderone, CT; Walsh, CT. FenF: servicing the mycosubtilin synthetase assembly line in trans. ChemBioChem 2007, 8, 613–616. [Google Scholar]
- Aron, ZD; Dorrestein, PC; Blackhall, JR; Kelleher, NL; Walsh, CT. Characterization of a new tailoring domain in polyketide biogenesis: the amine transferase domain of MycA in the mycosubtilin gene cluster. J. Am. Chem. Soc 2005, 127, 14986–14987. [Google Scholar]
- Dittmann, J; Wenger, RM; Kleinkauf, H; Lawen, A. Mechanism of cyclosporin A biosynthesis. Evidence for synthesis via a single linear undecapeptide precursor. J. Biol. Chem 1994, 269, 2841–2846. [Google Scholar]
- Walton, JD. Two enzymes involved in biosynthesis of the host-selective phytotoxin HC-toxin. Proc. Natl. Acad. Sci USA 1987, 84, 8444–8447. [Google Scholar]
- Balibar, CJ; Vaillancourt, FH; Walsh, CT. Generation of D amino acid residues in assembly of arthrofactin by dual condensation/epimerization domains. Chem. Biol 2005, 12, 1189–1200. [Google Scholar]
- Gross, H; Loper, JE. Genomics of secondary metabolite production by Pseudomonas spp. Nat. Prod. Rep 2009, 26, 1408–1446. [Google Scholar]
- Hutchison, ML; Gross, DC. Lipopeptide phytotoxins produced by Pseudomonas syringae pv. syringae: comparison of the biosurfactant and ion channel-forming activities of syringopeptin and syringomycin. Mol. Plant Microbe Interact 1997, 10, 347–354. [Google Scholar]
- Guenzi, E; Galli, G; Grgurina, I; Pace, E; Ferranti, P; Grandi, G. Coordinate transcription and physical linkage of domains in surfactin synthetase are not essential for proper assembly and activity of the multienzyme complex. J. Biol. Chem 1998, 273, 14403–14410. [Google Scholar]
- Vaillancourt, FH; Yin, J; Walsh, CT. SyrB2 in syringomycin E biosynthesis is a nonheme FeII alpha-ketoglutarate- and O2-dependent halogenase. Proc. Natl. Acad. Sci USA 2005, 102, 10111–10116. [Google Scholar]
- Singh, GM; Vaillancourt, FH; Yin, J; Walsh, CT. Characterization of SyrC, an aminoacyltransferase shuttling threonyl and chlorothreonyl residues in the syringomycin biosynthetic assembly line. Chem. Biol 2007, 14, 31–40. [Google Scholar]
- Singh, GM; Fortin, PD; Koglin, A; Walsh, CT. beta-Hydroxylation of the aspartyl residue in the phytotoxin syringomycin E: characterization of two candidate hydroxylases AspH and SyrP in Pseudomonas syringae. Biochemistry 2008, 47, 11310–11320. [Google Scholar]
- Scholz-Schroeder, BK; Soule, JD; Gross, DC. The sypA, sypB, and sypC synthetase genes encode twenty-two modules involved in the nonribosomal peptide synthesis of syringopeptin by Pseudomonas syringae pv. syringae B301D. Mol. Plant Microbe Interact 2003, 16, 271–280. [Google Scholar]
- Ikegami, T. Personal communication, Osaka University: Osaka, Japan, 2004.
- Roongsawang, N; Hase, K; Haruki, M; Imanaka, T; Morikawa, M; Kanaya, S. Cloning and characterization of the gene cluster encoding arthrofactin synthetase from Pseudomonas sp. MIS38. Chem. Biol 2003, 10, 869–880. [Google Scholar]
- Andersen, JB; Koch, B; Nielsen, TH; Sorensen, D; Hansen, M; Nybroe, O; Christophersen, C; Sorensen, J; Molin, S; Givskov, M. Surface motility in Pseudomonas sp. DSS73 is required for efficient biological containment of the root-pathogenic microfungi Rhizoctonia solani and Pythium ultimum. Microbiology 2003, 149, 37–46. [Google Scholar]
- Roongsawang, N; Washio, K; Morikawa, M. Hokkaido University: Hokkaido, Japan; Unpublished work; 2007. [Google Scholar]
- Roongsawang, N; Washio, K; Morikawa, M. In vivo characterization of tandem C-terminal thioesterase domains in arthrofactin synthetase. ChemBioChem 2007, 8, 501–512. [Google Scholar]
- Lim, SP; Roongsawang, N; Washio, K; Morikawa, M. Functional analysis of a pyoverdine synthetase from Pseudomonas sp. MIS38. Biosci. Biotechnol. Biochem 2007, 71, 2002–2009. [Google Scholar]
- Saini, HS; Barragan-Huerta, BE; Lebron-Paler, A; Pemberton, JE; Vazquez, RR; Burns, AM; Marron, MT; Seliga, CJ; Gunatilaka, AA; Maier, RM. Efficient purification of the biosurfactant viscosin from Pseudomonas libanensis strain M9-3 and its physicochemical and biological properties. J. Nat. Prod 2008, 71, 1011–1015. [Google Scholar]
- Kruijt, M; Tran, H; Raaijmakers, JM. Functional, genetic and chemical characterization of biosurfactants produced by plant growth-promoting Pseudomonas putida 267. J. Appl. Microbiol 2009, 107, 546–556. [Google Scholar]
- Dubern, JF; Coppoolse, ER; Stiekema, WJ; Bloemberg, GV. Genetic and functional characterization of the gene cluster directing the biosynthesis of putisolvin I and II in Pseudomonas putida strain PCL1445. Microbiology 2008, 154, 2070–2083. [Google Scholar]
- Magnuson, R; Solomon, J; Grossman, AD. Biochemical and genetic characterization of a competence pheromone from Bacillus subtilis. Cell 1994, 77, 207–216. [Google Scholar]
- Auchtung, JM; Lee, CA; Grossman, AD. Modulation of the ComA-dependent quorum response in Bacillus subtilis by multiple Rap proteins and Phr peptides. J. Bacteriol 2006, 188, 5273–5285. [Google Scholar]
- Roggiani, M; Dubnau, D. ComA, a phosphorylated response regulator protein of Bacillus subtilis, binds to the promoter region of srfA. J. Bacteriol 1993, 175, 3182–3187. [Google Scholar]
- Yakimov, MM; Golyshin, PN. ComA-dependent transcriptional activation of lichenysin A synthetase promoter in Bacillus subtilis cells. Biotechnol. Prog 1997, 13, 757–761. [Google Scholar]
- Griffith, KL; Grossman, AD. A degenerate tripartite DNA-binding site required for activation of ComA-dependent quorum response gene expression in Bacillus subtilis. J. Mol. Biol 2008, 381, 261–275. [Google Scholar]
- Wang, X; Luo, C; Liu, Y; Nie, Y; Zhang, R; Chen, Z. Three non-aspartate amino acid mutations in the ComA Response regulator receiver motif severely decrease surfactin production, competence development and spore formation in Bacillus subtilis. J. Microbiol. Biotechnol 2010, 20, 301–310. [Google Scholar]
- Nakano, MM; Zuber, P. Cloning and characterization of srfB, a regulatory gene involved in surfactin production and competence in Bacillus subtilis. J. Bacteriol 1989, 171, 5347–5353. [Google Scholar]
- Kim, SB; Shin, BS; Choi, SK; Kim, CK; Park, SH. Involvement of acetyl phosphate in the in vivo activation of the response regulator ComA in Bacillus subtilis. FEMS Microbiol. Lett 2001, 195, 179–183. [Google Scholar]
- Cosby, WM; Vollenbroich, D; Lee, OH; Zuber, P. Altered srf expression in Bacillus subtilis resulting from changes in culture pH is dependent on the Spo0K oligopeptide permease and the ComQX system of extracellular control. J. Bacteriol 1998, 180, 1438–1445. [Google Scholar]
- Hayashi, K; Ohsawa, T; Kobayashi, K; Ogasawara, N; Ogura, M. The H2O2 stress-responsive regulator PerR positively regulates srfA expression in Bacillus subtilis. J. Bacteriol 2005, 187, 6659–6667. [Google Scholar]
- Nakano, MM; Zhu, Y; Liu, J; Reyes, DY; Yoshikawa, H; Zuber, P. Mutations conferring amino acid residue substitutions in the carboxy-terminal domain of RNA polymerase alpha can suppress clpX and clpP with respect to developmentally regulated transcription in Bacillus subtilis. Mol. Microbiol 2000, 37, 869–884. [Google Scholar]
- Mader, U; Antelmann, H; Buder, T; Dahl, MK; Hecker, M; Homuth, G. Bacillus subtilis functional genomics: genome-wide analysis of the DegS-DegU regulon by transcriptomics and proteomics. Mol. Genet Genomics 2002, 268, 455–467. [Google Scholar]
- Hayashi, K; Kensuke, T; Kobayashi, K; Ogasawara, N; Ogura, M. Bacillus subtilis RghR (YvaN) represses rapG and rapH, which encode inhibitors of expression of the srfA operon. Mol. Microbiol 2006, 59, 1714–1729. [Google Scholar]
- Ogura, M; Fujita, Y. Bacillus subtilis rapD, a direct target of transcription repression by RghR, negatively regulates srfA expression. FEMS Microbiol. Lett 2007, 268, 73–80. [Google Scholar]
- Ohsawa, T; Tsukahara, K; Sato, T; Ogura, M. Superoxide stress decreases expression of srfA through inhibition of transcription of the comQXP quorum-sensing locus in Bacillus subtilis. J. Biochem 2006, 139, 203–211. [Google Scholar]
- Serror, P; Sonenshein, AL. CodY is required for nutritional repression of Bacillus subtilis genetic competence. J. Bacteriol 1996, 178, 5910–5915. [Google Scholar]
- Nakano, S; Nakano, MM; Zhang, Y; Leelakriangsak, M; Zuber, P. A regulatory protein that interferes with activator-stimulated transcription in bacteria. Proc. Natl. Acad. Sci USA 2003, 100, 4233–4238. [Google Scholar]
- Reyes, DY; Zuber, P. Activation of transcription initiation by Spx: formation of transcription complex and identification of a cis-acting element required for transcriptional activation. Mol. Microbiol 2008, 69, 765–779. [Google Scholar]
- Quadri, LE; Weinreb, PH; Lei, M; Nakano, MM; Zuber, P; Walsh, CT. Characterization of Sfp, a Bacillus subtilis phosphopantetheinyl transferase for peptidyl carrier protein domains in peptide synthetases. Biochemistry 1998, 37, 1585–1595. [Google Scholar]
- Tsuge, K; Ohata, Y; Shoda, M. Gene yerP, involved in surfactin self-resistance in Bacillus subtilis. Antimicrob. Agents Chemother 2001, 45, 3566–3573. [Google Scholar]
- Tsuge, K; Ano, T; Hirai, M; Nakamura, Y; Shoda, M. The genes degQ, pps, and lpa-8 (sfp) are responsible for conversion of Bacillus subtilis 168 to plipastatin production. Antimicrob. Agents Chemother 1999, 43, 2183–2192. [Google Scholar]
- Msadek, T; Kunst, F; Klier, A; Rapoport, G. DegS-DegU and ComP-ComA modulator-effector pairs control expression of the Bacillus subtilis pleiotropic regulatory gene degQ. J. Bacteriol 1991, 173, 2366–2377. [Google Scholar]
- Tsuge, K; Ano, T; Shoda, M. Isolation of a gene essential for biosynthesis of the lipopeptide antibiotics plipastatin B1 and surfactin in Bacillus subtilis YB8. Arch. Microbiol 1996, 165, 243–251. [Google Scholar]
- Ke, WJ; Chang, BY; Lin, TP; Liu, ST. Activation of the promoter of the fengycin synthetase operon by the UP element. J. Bacteriol 2009, 191, 4615–4623. [Google Scholar]
- Duitman, EH; Wyczawski, D; Boven, LG; Venema, G; Kuipers, OP; Hamoen, LW. Novel methods for genetic transformation of natural Bacillus subtilis isolates used to study the regulation of the mycosubtilin and surfactin synthetases. Appl. Environ. Microbiol 2007, 73, 3490–3496. [Google Scholar]
- Koumoutsi, A; Chen, XH; Vater, J; Borriss, R. DegU and YczE positively regulate the synthesis of bacillomycin D by Bacillus amyloliquefaciens strain FZB42. Appl. Environ. Microbiol 2007, 73, 6953–6964. [Google Scholar]
- Huang, CC; Ano, T; Shoda, M. Nucleotide sequence and characteristics of the gene, lpa-14, responsible for biosynthesis of the lipopeptide antibiotics iturinA and surfactin from Bacillus subtilis RB14. J. Ferment. Bioeng 1993, 76, 445–450. [Google Scholar]
- Heeb, S; Haas, D. Regulatory roles of the GacS/GacA two-component system in plant-associated and other gram-negative bacteria. Mol. Plant Microbe Interact 2001, 14, 1351–1363. [Google Scholar]
- Workentine, ML; Chang, L; Ceri, H; Turner, RJ. The GacS-GacA two-component regulatory system of Pseudomonas fluorescens: a bacterial two-hybrid analysis. FEMS Microbiol. Lett 2009, 292, 50–56. [Google Scholar]
- Willis, DK; Holmstadt, JJ; Kinscherf, TG. Genetic evidence that loss of virulence associated with gacS or gacA mutations in Pseudomonas syringae B728a does not result from effects on alginate production. Appl. Environ. Microbiol 2001, 67, 1400–1403. [Google Scholar]
- Koch, B; Nielsen, TH; Sorensen, D; Andersen, JB; Christophersen, C; Molin, S; Givskov, M; Sorensen, J; Nybroe, O. Lipopeptide production in Pseudomonas sp. strain DSS73 is regulated by components of sugar beet seed exudate via the Gac two-component regulatory system. Appl. Environ. Microbiol 2002, 68, 4509–4516. [Google Scholar]
- Nielsen, TH; Nybroe, O; Koch, B; Hansen, M; Sorensen, J. Genes involved in cyclic lipopeptide production are important for seed and straw colonization by Pseudomonas sp. strain DSS73. Appl. Environ. Microbiol 2005, 71, 4112–4116. [Google Scholar]
- Dubern, JF; Bloemberg, GV. Influence of environmental conditions on putisolvins I and II production in Pseudomonas putida strain PCL1445. FEMS Microbiol. Lett 2006, 263, 169–175. [Google Scholar]
- Dubern, JF; Lugtenberg, BJ; Bloemberg, GV. The ppuI-rsaL-ppuR quorum-sensing system regulates biofilm formation of Pseudomonas putida PCL1445 by controlling biosynthesis of the cyclic lipopeptides putisolvins I and II. J. Bacteriol 2006, 188, 2898–2906. [Google Scholar]
- Washio, K; Lim, SP; Roongsawang, N; Morikawa, M. Identification and characterization of the genes responsible for the production of the cyclic lipopeptide arthrofactin by Pseudomonas sp. MIS38. Biosci. Biotechnol. Biochem 2010, 74, 992–999. [Google Scholar]
- Dubern, JF; Lagendijk, EL; Lugtenberg, BJ; Bloemberg, GV. The heat shock genes dnaK, dnaJ, and grpE are involved in regulation of putisolvin biosynthesis in Pseudomonas putida PCL1445. J. Bacteriol 2005, 187, 5967–5976. [Google Scholar]
- Wang, N; Lu, SE; Records, AR; Gross, DC. Characterization of the transcriptional activators SalA and SyrF, Which are required for syringomycin and syringopeptin production by Pseudomonas syringae pv. syringae. J. Bacteriol 2006, 188, 3290–3298. [Google Scholar]
- de Bruijn, I; Raaijmakers, JM. Diversity and functional analysis of LuxR-type transcriptional regulators of cyclic lipopeptide biosynthesis in Pseudomonas fluorescens. Appl. Environ. Microbiol 2009, 75, 4753–4761. [Google Scholar]
- de Bruijn, I; Raaijmakers, JM. Regulation of cyclic lipopeptide biosynthesis in Pseudomonas fluorescens by the ClpP protease. J. Bacteriol 2009, 191, 1910–1923. [Google Scholar]
- Srivatsan, A; Wang, JD. Control of bacterial transcription, translation and replication by (p)ppGpp. Curr. Opin. Microbiol 2008, 11, 100–105. [Google Scholar]
- Finking, R; Solsbacher, J; Konz, D; Schobert, M; Schafer, A; Jahn, D; Marahiel, MA. Characterization of a new type of phosphopantetheinyl transferase for fatty acid and siderophore synthesis in Pseudomonas aeruginosa. J. Biol. Chem 2002, 277, 50293–50302. [Google Scholar]
- Barekzi, N; Joshi, S; Irwin, S; Ontl, T; Schweizer, HP. Genetic characterization of pcpS, encoding the multifunctional phosphopantetheinyl transferase of Pseudomonas aeruginosa. Microbiology 2004, 150, 795–803. [Google Scholar]
- Seidle, HF; Couch, RD; Parry, RJ. Characterization of a nonspecific phosphopantetheinyl transferase from Pseudomonas syringae pv. syringae FF5. Arch. Biochem. Biophys 2006, 446, 167–174. [Google Scholar]
- Quigley, NB; Mo, YY; Gross, DC. SyrD is required for syringomycin production by Pseudomonas syringae pv. syringae and is related to a family of ATP-binding secretion proteins. Mol. Microbiol 1993, 9, 787–801. [Google Scholar]
- Grgurina, I; Gross, DC; Iacobellis, NS; Lavermicocca, P; Takemoto, JY; Benincasa, M. Phytotoxin production by Pseudomonas syringae pv. syringae: Syringopeptin production by syr mutants defective in biosynthesis or secretion of syringomycin. FEMS Microbiol. Lett 1996, 138, 35–39. [Google Scholar]
- Kang, H; Gross, DC. Characterization of a resistance-nodulation-cell division transporter system associated with the syr-syp genomic island of Pseudomonas syringae pv. syringae. Appl. Environ. Microbiol 2005, 71, 5056–5065. [Google Scholar]
- Lim, SP; Roongsawang, N; Washio, K; Morikawa, M. Flexible exportation mechanisms of arthrofactin in Pseudomonas sp. MIS38. J. Appl. Microbiol 2009, 107, 157–166. [Google Scholar]
- Kobayashi, N; Nishino, K; Yamaguchi, A. Novel macrolide-specific ABC-type efflux transporter in Escherichia coli. J. Bacteriol 2001, 183, 5639–5644. [Google Scholar]
- Schneider, A; Stachelhaus, T; Marahiel, MA. Targeted alteration of the substrate specificity of peptide synthetases by rational module swapping. Mol. Gen. Genet 1998, 257, 308–318. [Google Scholar]
- Yakimov, MM; Giuliano, L; Timmis, KN; Golyshin, PN. Recombinant acylheptapeptide lichenysin: high level of production by Bacillus subtilis cells. J. Mol. Microbiol. Biotechnol 2000, 2, 217–224. [Google Scholar]
- de Ferra, F; Rodriguez, F; Tortora, O; Tosi, C; Grandi, G. Engineering of peptide synthetases. Key role of the thioesterase-like domain for efficient production of recombinant peptides. J. Biol. Chem 1997, 272, 25304–25309. [Google Scholar]
- Trauger, JW; Kohli, RM; Mootz, HD; Marahiel, MA; Walsh, CT. Peptide cyclization catalysed by the thioesterase domain of tyrocidine synthetase. Nature 2000, 407, 215–218. [Google Scholar]
- Kohli, RM; Trauger, JW; Schwarzer, D; Marahiel, MA; Walsh, CT. Generality of peptide cyclization catalyzed by isolated thioesterase domains of nonribosomal peptide synthetases. Biochemistry 2001, 40, 7099–7108. [Google Scholar]
- Stachelhaus, T; Mootz, HD; Marahiel, MA. The specificity-conferring code of adenylation domains in nonribosomal peptide synthetases. Chem. Biol 1999, 6, 493–505. [Google Scholar]
- Uguru, GC; Milne, C; Borg, M; Flett, F; Smith, CP; Micklefield, J. Active-site modifications of adenylation domains lead to hydrolysis of upstream nonribosomal peptidyl thioester intermediates. J. Am. Chem. Soc 2004, 126, 5032–5033. [Google Scholar]
- Mootz, HD; Kessler, N; Linne, U; Eppelmann, K; Schwarzer, D; Marahiel, MA. Decreasing the ring size of a cyclic nonribosomal peptide antibiotic by in-frame module deletion in the biosynthetic genes. J. Am. Chem. Soc 2002, 124, 10980–10981. [Google Scholar]
- Symmank, H; Franke, P; Saenger, W; Bernhard, F. Modification of biologically active peptides: production of a novel lipohexapeptide after engineering of Bacillus subtilis surfactin synthetase. Protein Eng 2002, 15, 913–921. [Google Scholar]
- Butz, D; Schmiederer, T; Hadatsch, B; Wohlleben, W; Weber, T; Sussmuth, RD. Module extension of a non-ribosomal peptide synthetase of the glycopeptide antibiotic balhimycin produced by Amycolatopsis balhimycina. ChemBioChem 2008, 9, 1195–1200. [Google Scholar]
- Hahn, M; Stachelhaus, T. Harnessing the potential of communication-mediating domains for the biocombinatorial synthesis of nonribosomal peptides. Proc. Natl. Acad. Sci USA 2006, 103, 275–280. [Google Scholar]
- Chiocchini, C; Linne, U; Stachelhaus, T. In vivo biocombinatorial synthesis of lipopeptides by COM domain-mediated reprogramming of the surfactin biosynthetic complex. Chem. Biol 2006, 13, 899–908. [Google Scholar]
- Vater, J; Wilde, C; Kell, H. In situ detection of the intermediates in the biosynthesis of surfactin, a lipoheptapeptide from Bacillus subtilis OKB 105, by whole-cell cell matrix-assisted laser desorption/ionization time-of-flight mass spectrometry in combination with mutant analysis. Rapid. Commun. Mass Spectrom 2009, 23, 1493–1498. [Google Scholar]
- Ackerley, DF; Lamont, IL. Characterization and genetic manipulation of peptide synthetases in Pseudomonas aeruginosa PAO1 in order to generate novel pyoverdines. Chem. Biol 2004, 11, 971–980. [Google Scholar]
- Baltz, RH. Biosynthesis and genetic engineering of lipopeptides in Streptomyces roseosporus. Methods Enzymol 2009, 458, 511–531. [Google Scholar]
- Nguyen, KT; He, X; Alexander, DC; Li, C; Gu, JQ; Mascio, C; Van Praagh, A; Mortin, L; Chu, M; Silverman, JA; Brian, P; Baltz, RH. Genetically engineered lipopeptide antibiotics related to A54145 and daptomycin with improved properties. Antimicrob. Agents Chemother 2010, 54, 1404–1413. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Structure | Ref. |
|---|---|---|
| Surfactin family | ||
| Surfactin | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Val-l-Asp-d-Leu-l-Leu | [31] |
| Lichenysin | A/D FA-β-OH-l-Gln-l-Leu-d-Leu-l-Val-l-Asp-d-Leu-l-Ile | [7,32] |
| (Lichenysin B) | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Val-l-Asp-d-Leu-l-Leu | [33] |
| (Lichenysin C) | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Val-l-Asp-d-Leu-l-Ile | [34] |
| Lichenysin | G FA-β-OH-l-Gln-l-[A2]-d-Leu-l-[A4]-l-Asp-d-Leu-l-[A7] A2 = Leu/Ile, A4 = Val/Ile, A7 = Ile/Val | [6] |
| Surfactant BL86 | FA-β-OH-l-Glx-l-Leu-d-Leu-l-Val-l-Asx-d-Leu-l-[A7] A7 = Ile/Val | [35] |
| Pumilacidin | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Leu-l-Asp-d-Leu-l-[A7] A7 = Ile/Val | [36] |
| Fengycin family | ||
| Fengycin | FA-β-OH-l-Glu-d-Orn-d-Tyr-d-aThr-l-Glu-d-[A6]-l-Pro-l-Gln-l-Tyr-l-Ile A6 = Ala/Val | [37] |
| Plipastatin | FA-β-OH-l-Glu-D-Orn-l-Tyr-D-aThr-l-Glu-D-[A6]-l-Pro-l-Gln-D-Tyr-l-Ile A6 = Ala/Val | [37] |
| Iturin family | ||
| Iturin A | FA-β-NH2-l-Asn-d-Tyr-d-Asn-l-Gln-l-Pro-d-Asn-l-Ser | [38] |
| Iturin C | FA-β-NH2-l-Asp-d-Tyr-d-Asn-l-Gln-l-Pro-d-Asn-l-Ser | [37] |
| Bacillomycin L | FA-β-NH2-l-Asn-d-Tyr-d-Asn-l-Ser-l-Glu-d-Ser-l-Thr | [39] |
| Bacillomycin D | FA-β-NH2-l-Asn-d-Tyr-d-Asn-l-Pro-l-Glu-d-Ser-l-Thr | [37] |
| Bacillomycin F | FA-β-NH2-l-Asn-d-Tyr-d-Asn-l-Gln-l-Pro-d-Asn-l-Thr | [37] |
| Mycosubtilin | FA-β-NH2-l-Asn-d-Tyr-d-Asn-l-Gln-l-Pro-d-Ser-l-Asn | [40] |
| Other | ||
| Antiadhesin | FA-β-OH-l-Asp-l-Leu-l-Leu-l-Val-l-Val-l-Glu-l-Leu | [41] |
| Bamylocin A | FA-β-OH-x-Glu-x-Leu-x-Met-x-Leu-x-Pro-x-Leu-x-Leu | [42] |
| Circulocin 1 | gFA-β-OH-x-Thr-x-Phe-x-Ile-x-Dba-x-Asp | [43] |
| Circulocin 3 | gFA-β-OH-x-Thr-x-Leu-x-Ile-x-Thr-x-Asn-x-Ala | [43] |
| Fusaricidin | gFA-β-OH-l-Thr-d-Val-l-Tyr-d-aThr-d-Asn-d-Ala | [44] |
| Kurstakins | FA-x-Thr-x-Gly-x-Ala-x-Ser-x-His-x-Gln-x-Gln | [45] |
| Group/Name | Structure | Ref. |
|---|---|---|
| ViscosinA | ||
| Viscosin | FA-β-OH-l-Leu-d-Glu-d-aThr-d-Val-l-Leu-d-Ser-l-Leu-d-Ser-l-Ile | [46] |
| Viscosinamide | FA-β-OH-l-Leu-d-Gln-d-aThr-d-Val-l-Leu-d-Ser-l-Leu-d-Ser-l-Ile | [47] |
| Massetolide A | FA-β-OH-l-Leu-d-Glu-d-aThr-d-aIle-l-Leu-d-Ser-l-Leu-d-Ser-l-Ile | [48] |
| Pseudophomin A | FA-β-OH-l-Leu-d-Glu-d-aThr-d- Ile-d-Leu-d-Ser-l-Leu-d-Ser-l-Ile | [49] |
| Pseudodesmin A | FA-β-OH-l-Leu-d-Gln-d-aThr-d-Val-d-Leu-d-Ser-l-Leu-d-Ser-l-Ile | [50] |
| SyringomycinB | ||
| Syringomycin A | FA-β-OH-l-Ser-d-Ser-d-Dab-l-Dab-l-Arg-l-Phe-z-Dhb-l-Asp(3-OH)-l-Thr(4-Cl) | [51] |
| Syringostatin A | FA-β-OH-l-Ser-d-Dab-l-Dab-d-Hse-l-Orn-l-aThr-z-Dhb-l-Asp(3-OH)-l-Thr(4-Cl) | [52] |
| Syringotoxin B | FA-β-OH-l-Ser-d-Dab-l-Gly-d-Hse-l-Orn-l-aThr-z-Dhb-l-Asp(3-OH)-l-Thr(4-Cl) | [53] |
| Pseudomycin A | FA-β-OH-l-Ser-d-Dab-l-Asp-d-Lys-l-Dab-l-aThr-z-Dhb-l-Asp(3-OH)-l-Thr(4-Cl) | [54] |
| Cormycin A | FA-β-OH-l-Ser-d-Orn-l-Asn-d-Hse-l-His-l-aThr-z-Dhb-l-Asp(3-OH)-l-Thr(4-Cl) | [55] |
| AmphisinA | ||
| Arthrofactin | FA-β-OH-d-Leu-d-Asp-d-aThr-d-Leu-d-Leu-d-Ser-l-Leu-d-Ser-l-Ile-l-Ile-l-Asp | [56] |
| Amphisin | FA-β-OH-d-Leu-d-Asp-d-aThr-d-Leu-d-Leu-d-Ser-l-Leu-d-Gln-l-Leu-l-Ile-l-Asp | [10] |
| Lokisin | FA-β-OH-d-Leu-d-Asp-d-aThr-d-Leu-d-Leu-d-Ser-l-Leu-d-Ser-l-Leu-l-Ile-l-Asp | [11] |
| Pholipeptin | FA-β-OH-d-Leu-l-Asp-l- Thr-d-Leu-d-Leu-d-Ser-D-leu-d-Ser-d-Leu-l-Ile-d-Asp | [12] |
| Tensin | FA-β-OH-d-Leu-d-Asp-d-aThr-d-Leu-d-Leu-d-Ser-l-Leu-d-Gln-l-Leu-l-Ile-l-Glu | [57] |
| PutisolvinC | ||
| Putisolvin I | CH3(CH)4CO-d-Leu-d-Glu-d-Leu-d-Ile-d-Gln-d-Ser-d-Val-d-Ile-d-Ser-l-Leu-l-Val-x-Ser | [58] |
| Putisolvin II | CH3(CH)4CO-d-Leu-d-Glu-d-Leu-d-Ile-d-Gln-d-Ser-d-Val-d-Ile-d-Ser-l-Leu-l-Xle-x-Ser | [58] |
| TolaasinA | ||
| Tolaasin I | FA-β-OH-Dhb-Pro-Ser-Leu-Val-Ser-Leu-Val-Val-Gln-Leu - - - - Val-Dhb-aThr-Ile-Hse-Dab-Lys | [59] |
| Fuscopeptin | FA-β-OH-Dhb-Pro-Leu-Ala-Ala-Ala-Ala-Val-Gly-Ala-Val-Ala - - - Val-Dhb-aThr-Ala-Dab-Dab-Phe | [60] |
| Corpeptin | FA-β-OH-Dhb-Pro-Ala-Ala-Ala-Val-Val-Dhb-Hse-Val-aIle-Dhb-Ala-Ala-Ala-Val-Dhb-aThr-Ala-Dab-Ser-Ile | [61] |
| SyringopeptinA | ||
| SP22 | FA-β-OH-Dhb-Pro-Val-Val-Ala-Ala-Val - - - Val-Dhb-Ala-Val-Ala-Ala-Dhb-aThr-Ser-Ala-Dhb-Ala-Dab-Dab-Tyr | [62] |
| SP25 | FA-β-OH-Dhb-Pro-Val-Ala-Ala-Val-Leu-Ala-Ala-Dhb-Val-Dhb-Ala-Val-Ala-Ala-Dhb-aThr-Ser-Ala-Val-Ala-Dab-Dab-Tyr | [63] |
| SP25[Phe25] | FA-β-OH-Dhb-Pro-Val-Ala-Ala-Val-Leu-Ala-Ala-Dhb-Val-Dhb-Ala-Val-Ala-Ala-Dhb-aThr-Ser-Ala-Val-Ala-Dab-Dab-Phe | [64] |
| Other | ||
| EntolysinD | FA-β-OH-d-Xle-d-Glu-d-Gln-d-Val-d-Xle-d-Gln-d-Val-d-Xle-d-Gln-d-Ser-l-Val-l-Xle-d-Ser-x-Xle | [65] |
| OfamideA | FA-β-OH-l-Leu-d-Glu-d-aThr-d-aIle-l-Leu-d-Ser-l-Leu-l-Leu-d-Ser-l-Val | [66] |
| PseudofactinA | CH3(CH)14CO-x-Gly-x-Ser-x-Thr-x-Leu-x-Leu-x-Ser-x-Leu-x-Leu/Val | [67] |
| SyringafactinE | FA-β-OH-d-Leu-d-Leu-d-Gln-l-Leu-d-Thr-l-Val-d-Leu-l-Leu | [68] |
| Name | Structure | Ref. |
|---|---|---|
| Cyclic products | ||
| Surfactin[Val7] | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Val-l-Asp-d-Leu-l-Val | [14] |
| Surfactin[Phe7] | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Val-l-Asp-d-Leu-l-Phe | [14] |
| Surfactin[Orn7] | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Val-l-Asp-d-Leu-l-Orn | [14] |
| Surfactin[Cys7] | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Val-l-Asp-d-Leu-l-Cys | [14] |
| Surfactin[Gln1] | FA-β-OH-l-Gln-l-Leu-d-Leu-l-Val-l-Asp-d-Leu-l-Leu | [155] |
| Surfactin[Asn5] | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Val-l-Asn-d-Leu-l-Leu | [13] |
| Surfactin[Orn2] | FA-β-OH-l-Glu-l-Orn-d-Leu-l-Val-l-Asp-d-Leu-l-Leu | [154] |
| Surfactin[Orn2ΔLeu3/Val4] | FA-β-OH-l-Glu-l-Orn-l-Asp-d-Leu-l-Leu | [154] |
| Surfactin[Orn2ΔLeu6/7] | FA-β-OH-l-Glu-l-Orn-d-Leu-l-Val-l-Asp | [154] |
| Surfactin[ΔLeu2] | FA-β-OH-l-Glu-d-Leu-l-Val-l-Asp-d-Leu-l-Leu | [161,162] |
| Linear products | ||
| Surfactin[ΔLeu7] | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Val-l-Asp-d-Leu | [14] |
| Surfactin[ΔLeu6/7] | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Val-l-Asp | [156] |
| Surfactin[ΔAsp5Leu6/7] | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Val | [156] |
| Surfactin[ΔVal4Asp5Leu6] | FA-β-OH-l-Glu-l-Leu-d-Leu-l-Leu | [165] |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Roongsawang, N.; Washio, K.; Morikawa, M. Diversity of Nonribosomal Peptide Synthetases Involved in the Biosynthesis of Lipopeptide Biosurfactants. Int. J. Mol. Sci. 2011, 12, 141-172. https://doi.org/10.3390/ijms12010141
Roongsawang N, Washio K, Morikawa M. Diversity of Nonribosomal Peptide Synthetases Involved in the Biosynthesis of Lipopeptide Biosurfactants. International Journal of Molecular Sciences. 2011; 12(1):141-172. https://doi.org/10.3390/ijms12010141
Chicago/Turabian StyleRoongsawang, Niran, Kenji Washio, and Masaaki Morikawa. 2011. "Diversity of Nonribosomal Peptide Synthetases Involved in the Biosynthesis of Lipopeptide Biosurfactants" International Journal of Molecular Sciences 12, no. 1: 141-172. https://doi.org/10.3390/ijms12010141