Isolation and Characterization of Microsatellite Loci in the Asian Rice Gall Midge (Orseolia oryzae) (Diptera: Cecidomyiidae)

Abstract
:1. Introduction
2. Results and Discussion
2.1. Microsatellite Loci and Polymorphism
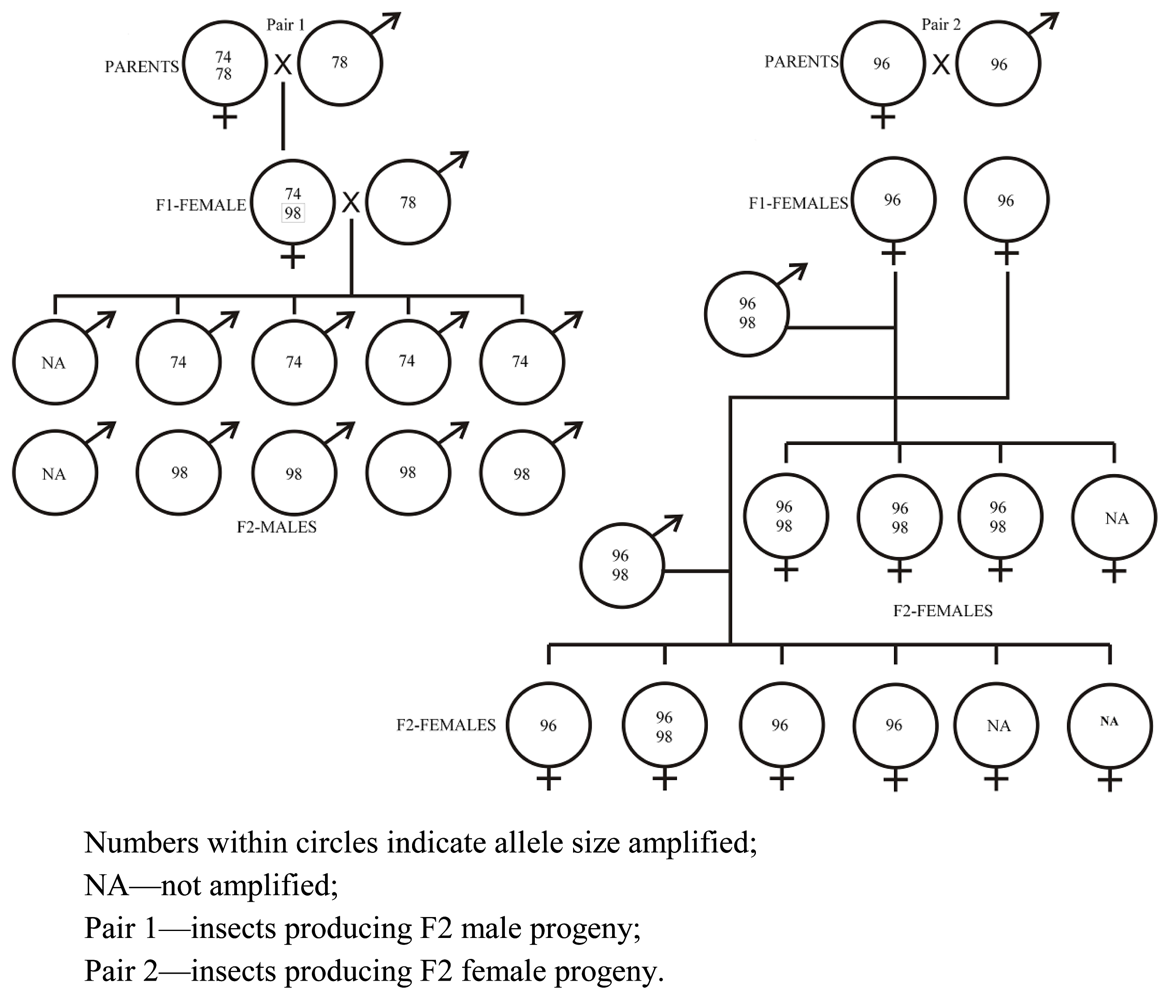
2.2. Inheritance of SSR Markers
3. Experimental Section
3.1. Insect Colonies
3.2. Isolation of Microsatellite Loci
3.3. Detection of Polymorphism and Data Analysis
3.4. Inheritance of SSR Markers
4. Conclusions
Supplementary Material

{kind=link}
{kind=link}
{kind=link}
| Accession Number (GeneBank) | Locus | Repeat Motif | Primer | Ann. temp. | MgCl2 conc. | Alleles Size |
|---|---|---|---|---|---|---|
| HM804497 | Oosat01 | (GA)10 | F:TCATCAAAAGGCAATGAGAAA R:GAAGACAACACACCGCACAT | 59 °C | 1.5 mM | 159 |
| HM804498 | Oosat02 | (TTAAAAT)2 N….(ATTTTA)4 | F:TGCACAAAAATAACGCAGGA R:ATAACCCAACCAAACCACGA | 60 °C | 2.0 mM | 263 |
| HM804500 | Oosat04 | (TC)10…(TC)3 | F:TGCACAAAAATTGCGATTCTAC R:CCCATATTGGGCAGCATCT | 59 °C | 3.0 mM | 340 |
| HM804501 | Oosat05 | (CA)6 | F:ATGATCATGCTGCTGTGCTC R:TGCGCTATTCTCCCCAGTAG | 61 °C | 3.0 mM | 134 |
| HM804502 | Oosat06 | (GT)4…(CG)2...(GT)4...(GT)5 | F:ATCTCAATCTTGGCGCTGTT R:TGCGAGCAATGAAACAAAAG | 59 °C | 1.5 mM | 157 |
| HM804503 | Oosat07 | (GT)27 | F:TGCAGAATTCGGCTTAGTGA R:GGGCAAATTCTCTGTCTCGT | 50 °C | 2.0 mM | 124 |
| HM804504 | Oosat08 | (CA)8 | F:GGCTTACTGCATCAGACTCTTTT R:AGCAGAATCGCTCTTTACGG | 52 °C | 2.0 mM | 109 |
| HM804505 | Oosat09 | (GT)8...(TG)2(CT)2 | F:CGAATTATATGATCGCGGAAA R:AAAATGTGTTTTGGGTCGAGA | 58 °C | 1.5 mM | 152 |
| HM804506 | Oosat10 | (CA)8…(CA)6….(CA)5 | F:TGACGTCAATAGCGACAACG R:GGCTTAGTGAGTGAGTGTGTGAG | 56 °C | 2.0 mM | 164 |
| HM804507 | Oosat11 | (GA)5 | F:TCTCACAGTCGCATGTTATC R:ATGCTGAGAGCATCTGAAAT | 56 °C | 3.0 mM | 110 |
| HM804508 | Oosat12 | (TC)9 | F:TATGCAAAGTGCGCGATATT R:TGTGCAGCCATTTACTGTGC | 56 °C | 2.0 mM | 361 |
| HM804509 | Oosat13 | (GA)23 | F:GGGAAACGATGATGATAATG R:TTCAGTTGGTGAGTATTCATGT | 52 °C | 1.5 mM | 145 |
| HM804510 | Oosat14 | (TA)3...(GT)4...(GT)2 | F:GTTCAGGCACCATGATATTT R:GTAGATTTTTCTTCGCAAGG | 54 °C | 1.5 mM | 284 |
| HM804511 | Oosat15 | (AG)13...(AG)8 | F:TAATAAAAAGGGCTGTCTGC R:GCATACAGACAGAAAAATCAA | 52 °C | 3.0 mM | 114 |
| HM804513 | Oosat17 | (TA)6 | F:CGAAATCGAACACAAACTTC R:GTTGAGCAGTTCAGGAGATT | 55 °C | 1.5 mM | 115 |
| HM804514 | Oosat18 | (CAA)8 | F:GCCAGATAATGAAGCCGAGA R:GCCGAAATGCATTAATGGTC | 52 °C | 1.5 mM | 255 |
| HM804515 | Oosat19 | (CG)4(GC)3 | F:AGGTGGAAGTGTTCGACGAG R:GAAGAACTCACGGTCGATGG | 57 °C | 2.5 mM | 218 |
| HM804516 | Oosat20 | (CA)27 | F:TTTCGAAACGAAATCGAAAT R:GCTAGCAGAGTGGATGAGCA | 52 °C | 2.0 mM | 111 |
| HM804518 | Oosat22 | (GT)13 | F:TTTGGGCCATGTATGAGAGC R:TGGAAGACTGAAGGGAAGACA | 55 °C | 2.0 mM | 234 |
| HM804519 | Oosat23 | (CA)66 | F:TTTGGGCCATGTATGAGAGC R:TGGAAGACTGAAGGGAAGACA | 59 °C | 2.0 mM | 206 |
| HM804521 | Oosat25 | (GA)6…(GA)3 | F:GTTGGTATCTGGTCGAGACG R:ACGCGCTTACCTGTTCAAAT | 53 °C | 2.5 mM | 131 |
| HM804523 | Oosat27 | (GA)14 | F:TCGAAAATCAGCTGAACGAA R:AACTCTTACACCCACACATATTCA | 52 °C | 3.0 mM | 129 |
| HM804524 | Oosat28 | (GTT)3 ATT (GTT)3 | F:CGCCTTTTTGCAAATTCTCT R:TGGATATTTGGTGTAAGGCAGA | 51 °C | 3.0 mM | 115 |
| HM804525 | Oosat29 | (ATT)3(ATG)3…(CAT)2 | F:CGTTCATGTGAATTGGTTGG R:TGTATACGAAGGTGGCGATG | 51 °C | 3.0 mM | 137 |
| HM804526 | Oosat30 | (GTT)7 | F:GCCGAAATGCATTAATGGTC R:TTCGATTATGGCATGGTTCA | 50 °C | 2.5 mM | 140 |
| HM804527 | Oosat31 | (TCCGTT)2…(GAATT)2 (TCCGTT)2..…(TCCGTT)2 | F:ATCCGCGATCAATTATTCTG R:TCCAAGTCCGTGAAATCAAA | 48 °C | 3.0 mM | 292 |
| HM804528 | Oosat32 | (GA)9 | F:CATGACACCATCCGATGAAT R:CACATAAACAACCAGGCACAA | 50 °C | 1.5 mM | 105 |
| HM804529 | Oosat33 | (AC)6..(AC)6..(AC)3..(AC)7.. (AC)3..(AC)4 | F:CGACACACACGAAACACACA R:TTTCGGGCACCACTTTACTC | 55 °C | 2.5 mM | 233 |
| HM804530 | Oosat34 | (CAA)12 | F:TGAGGCAGAATGAAAGAGCA R:CCATGGCACACGATAACAAT | 51 °C | 1.5 mM | 156 |
| HM804533 | Oosat37 | (GT)10 | F:TTCGACCGACTGACTGAGTG R:GAGACGTCGGTCGTGATTTT | 53 °C | 1.5 mM | 137 |
| HM804534 | Oosat38 | (CTT)6 | F:AACGGTTATAGAGTCGCGATG R:CGTGTGTTTCCTCACTAGAATCG | 53 °C | 1.5 mM | 106 |
| HM804535 | Oosat39 | (GT)3...(TG)5 | F:AACTGGCCACGGTCATTATC R:AATACGTCGACGGAAGAACG | 53 °C | 1.5 mM | 121 |
| HM804536 | Oosat40 | (GT)(AT)(GT)4 | F:GACCCAATCCACTTTGATCCT R:TGTCATCTAAAAGTATGTGCAACTGA | 55 °C | 1.5 mM | 156 |
| HM804537 | Oosat41 | (CT)13 | F:TCGTTGGAATAGCACATTCG R:TGACGTGTCTATGCCATGTG | 54 °C | 1.5 mM | 167 |
| HM804538 | Oosat42 | (TAA)7(TAG)(TAA)3 | F:GAGAGCAATTTTGATTCGACTTG R:GGGCCGAATGAAACAACTAC | 54 °C | 1.5 mM | 150 |
| HM804540 | Oosat44 | (GT)17 | F:GAAAAGCCGTTCGTTGAATC R:TTTCCACCAAATAAGAAAACCA | 50 °C | 2.0 mM | 193 |
| HM804541 | Oosat45 | (GA)8…(GA)6 | F:GAGTGAAAGAAGTCACGCACA R :GGCATCCACAGTCGAAGAT | 50 °C | 2.0 mM | 145 |
| HM804543 | Oosat47 | (GT)21 | F:CGAAGTGAATGTTTAATGGTTT R:CCGGTTTGTATAATTGTGAA | 56 °C | 2.5 mM | 128 |
| HM804544 | Oosat48 | (GA)14…(GA)8 | F:ACGCTGATCAAAAGAGTTCAG R:CCCTTGATAACAGAAAGTGAGAAC | 52 °C | 2.5 mM | 116 |
| HM804545 | Oosat49 | (CT)10GT(CT)2CA(GT)2AT (GT)4GA(GT)20 | F:CAACGTCCCATAGTCTGCATT R:AGCGGCAGTGTTTTCTCTTC | 55 °C | 1.5 mM | 168 |
| HM804546 | Oosat50 | AG (AC)2 (TC)2 AC | F:TGAGATGATATGTTCCTTTTTGTC R:GCAGTTCCGAGATGTTTGTG | 53 °C | 1.5 mM | 162 |
| HM804547 | Oosat51 | (AT)3…(TA)5…(AT)4 | F:GGTTTGACGGGCACTGTAT R:CGGCCACTGTATCTATAGGC | 50 °C | 2.0 mM | 269 |
| HM804548 | Oosat52 | (ACT)3(TGT)4 | F:AACTTGGAATGAAGCGTTCG R:CGAGGTCTACCTCTACCCATAGAT | 55 °C | 1.5 mM | 141 |
| HM804549 | Oosat53 | (ATG)4…ATG | F:GAGTTGCTTTGAAACGATTGC R:ATCGTCGGATGAGTGTTTGA | 54 °C | 1.5 mM | 110 |
| HM804550 | Oosat54 | (GT)8 | F:CTTGGCGTTTCGTTTATCTCA R:CCAATAAAGCAAGCACGTGTAA | 54 °C | 1.5 mM | 149 |
| HM804552 | Oosat56 | (GAAC)4…(GAAC)4 (GAAC)4 | F:TGCGAACATTCAACGACCTA R:ACCACGCATACGTCAGGACT | 56 °C | 1.5 mM | 138 |
| HM804553 | Oosat57 | (TTG)8 | F:CGATGTAGGCAACATTTTCG R:CCAATGAACATTCCCATCAA | 52 °C | 1.5 mM | 100 |
| HM804554 | Oosat58 | ACA (CAA)6 ACA | F:GTTCGGTCGGTGTCTTTTTC R:AAAGACCACACGCTGAAAGG | 48 °C | 2.0 mM | 111 |
| HM804556 | Oosat60 | (TA)4TC(TA)2TA TC (TA)2TT(AT)6 | F:CCAGTGATTTGAGCATCGAG R:TCTTGGTTTTACGACCATTTCA | 54 °C | 1.5 mM | 186 |
| HM804557 | Oosat61 | (AGC)2N5(CAG)2ACA ..(GAC)2 | F:GTCTGACTGGCATCACCAGA R:CGGCTATTTCCTGTCGGTAG | 55 °C | 1.5 mM | 152 |
| HM804558 | Oosat62 | (CA)3(CT)2(CA)14 | F:GGAACCATTAAACACTCACTTCG R:GGAAATTCATGGTCCGAAAA | 53 °C | 1.5 mM | 129 |
| HM804559 | Oosat63 | (TG)16N4(CT)6 | F:GAAGCACTGCAACAACCAAA R:TGTTCGCTCACACCGTTTAG | 54 °C | 1.5 mM | 147 |
| HM804560 | Oosat64 | (CA)3 AG (AC)2 | F:TGAGACAGTTTTCGACTCCTTG R:TAGAGGGCTTTTTCGACTGC | 54 °C | 1.5 mM | 131 |
| HM804561 | Oosat65 | (GTT)3Nn(AT)CA(TC)2 (CA)2 | F:TTCCTAGAATGTGGCGTTTG R:TTGAACGCAGGTTTAATTGC | 50 °C | 2.0 mM | 194 |
| HM804562 | Oosat66 | (ACA)3(TC)2CT(TC)13N3(CA)14 | F:TGCATTTCCGACAGGTTTTA R:CACCTATCGTCTTAAAGGAAATGA | 53 °C | 1.5 mM | 167 |
| HM804563 | Oosat67 | (AT)2(TA)3 | F:CTGTGCACACTTTGCCATTC R:ACTCCGTGTATGCGGAAAAG | 55 °C | 1.5 mM | 202 |
| HM804564 | Oosat68 | (TTTC)3 | F:CGCATGAAATTTGGATCAGC R:ATCGTGCCAAAAGTGACTGA | 54 °C | 1.5 mM | 101 |
| HM804565 | Oosat69 | (TC)7CATA(CA)3CTCA | F:CCGGATAGATAGCCGTGTTT R:CTCACTTGGTGGGTGAGTACC | 56 °C | 2.0 mM | 115 |
| HM804566 | Oosat70 | (GA)3TA(GA)3Nn(GA)4 | F:ATTTGGCCATGGCTATTTGA R:GCTGGGGGCTAATCTCTCTC | 54 °C | 1.5 mM | 103 |
| HM804567 | Oosat71 | (AT)2(AC)3 (CCA)3 | F:GTGTGCGCACTTTACTGGTG R:TGTGGTGGATTTGCTTTTTG | 55 °C | 1.5 mM | 199 |
| HM804568 | Oosat72 | (CAT)N2 (CAT)4 | F:TCGTCGGATGAGTGTTTGAT R:CAGAGGAGTTGCTTTGAAACG | 54 °C | 1.5 mM | 114 |
| HM804569 | Oosat73 | (TG)12 | F:GGAAAACATGTCGGCAGAA R:TGCACATGGTGTTGTTGTTG | 62 °C | 2.0 mM | 123 |
| HM804570 | Oosat74 | (AG)9 | F:TGAACATTGATACAGTGCGACA R:TGTGTCCGGGCCAATCTA | 55 °C | 1.5 mM | 123 |
| HM804571 | Oosat75 | (CT)4 N3 (TC)7 N2 (TC)9 N2 (CT)7 | F:CAGTTTCGGTTCGTTTTTCA R:CTTGCCATCCATTCATCAGA | 56 °C | 2.0 mM | 127 |
| HM804572 | Oosat76 | (GTT)5 | F:GGAAATTTTATTTCGGGAATTCAT R:ACCAAAGCTTTTCAACAACAG | 52 °C | 1.5 mM | 190 |
| HM804573 | Oosat77 | (GTC)4 GTT (GTC)2 | F:CGAATTCAGCACGAACACTG R:ACGTTTTCGATCACCGTTTC | 55 °C | 1.5 mM | 180 |
| HM804576 | Oosat80 | (TC)16 | F:TTGAAAAGTGAGGCTGATG R:TTAAACGTCCATCAAGTGAG | 55 °C | 1.5 mM | 237 |
| HM804577 | Oosat81 | (AG)15 | F:TAAGCGATGTTGCTTGC R:CGATTTTGTCGTTGTGC | 55 °C | 1.5 mM | 188 |
| HM804578 | Oosat82 | (GT)17 | F:AAATGAAAAGCCGTTCG R:TGCTAGCAGTTTCATTTCC | 52 °C | 1.5 mM | 211 |
| HM804580 | Oosat84 | (GT)16 | F: CAATCGTTTCAGTTCCTTT R:CACCCAAAATTCAATCG | 54 °C | 1.5 mM | 160 |
| HM804581 | Oosat85 | (TC)16 | F: CTAGCAGAATCACATTGA R:CAAATCATGCTCATAGTTCC | 55 °C | 1.5 mM | 358 |
| HM804582 | Oosat86 | (AG)15 | F:TAAGCGATGTTGCTTGC R:GCGATTTGTCGTTGTGC | 55 °C | 1.5 mM | 188 |
| HM804583 | Oosat87 | (AC)15 | F:TCCACCAAATACAGAAAACC R:AAATGAAAAGCCGTTCG | 52 °C | 1.5 mM | 192 |
| HM804585 | Oosat89 | (TG)16 | F:GAAGCACTGCAACAACC R:GATCTGTTCGCTCACACC | 55 °C | 1.5 mM | 151 |
| HM804586 | Oosat90 | (TG)16 | F: GAACATTATATTTTGAAAG R: AATGAAGCCTGAAGAAAGC | 55 °C | 1.5 mM | 206 |
Acknowledgements
References
- Bentur, JS. Insect pests of rice and their management. In Pests and Pathogens: Management Strategies; Reddy, DV, Rao, PN, Rao, KV, Eds.; BS Publications: Hyderabad, India, 2010; pp. 1–42. [Google Scholar]
- Bentur, JS; Pasalu, IC; Sarma, NP; Prasad Rao, U; Mishra, B. Gall Midge Resistance in Rice; Research Paper Series 01; Directorate of Rice Research: Hyderabad, India, 2003; p. 22. [Google Scholar]
- Khush, GS; Panda, N. Host Plant Resistance in Rice; International Rice Research Institute: Manila, Philippines, 1995; p. 240. [Google Scholar]
- Himabindu, K; Suneetha, K; Sama, VSAK; Bentur, JS. A new rice gall midge resistance gene in the breeding line CR57-MR1523, mapping with flanking markers and development of NILs. Euphytica 2009, 174, 179–187. [Google Scholar]
- Vijayalakshmi, P; Amudhan, S; Himabindu, K; Cheralu, C; Bentur, JS. A new biotype of the Asian rice gall midge Orseolia oryzae (Diptera: Cecidomyiidae) characterized from the Warangal population in Andhra Pradesh, India. Int. J. Trop. Ins. Sci 2006, 26, 207–211. [Google Scholar]
- Bentur, JS; Cheralu, C; Rao, PRM. Monitoring virulence in Asian rice gall midge populations in India. Entomol. Exp. Appl 2008, 129, 96–106. [Google Scholar]
- Behura, SK; Sahu, SC; Rajamani, S; Devi, A; Mago, R; Nair, S; Mohan, M. Differentiation of Asian rice gall midge, Orseolia oryzae (Wood-Mason), biotypes by sequence characterized amplified regions (SCARs). Insect Mol. Biol 1999, 8, 391–397. [Google Scholar]
- Katiyar, SK; Chandel, G; Tan, Y; Zhang, Y; Huang, B; Nugaliyadde, L; Fernando, K; Bentur, JS; Inthavong, S; Constantino, S; Bennett, J. Biodiversity of Asian rice gall midge (Orseolia oryzae Wood-Mason) from five countries examined by AFLP analysis. Genome 2000, 43, 322–332. [Google Scholar]
- Rozen, S; Skaletsky, H. Primer3 on the WWW for general users and for biologist programMers. In Bioinformatics Methods and Protocols: Methods in Molecular Biology; Krawetz, S, Misener, S, Eds.; Humana Press: Totowa, NJ, USA, 2000; pp. 365–386. [Google Scholar]
- Schemerhorn, BJ; Crane, YM; Morton, PK. Development of polymorphic microsatellite markers in Hessian fly, Mayetiola destructor (Say). Mol. Ecol Notes 2008, 8, 1360–1362. [Google Scholar]
- Marshal, TC; Slate, J; Kruuk, LEB; Pemberton, JM. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol 1998, 7, 639–655. [Google Scholar]
- Guo, S; Thompson, E. Performing the exact test of Hardy-Weinberg proportion for multiple alleles. Biometrics 1992, 48, 361–372. [Google Scholar]
- Oosterhout, CV; Hutchinson, WF; Wills, DPM; Shipley, P. Micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol Notes 2004, 4, 535–538. [Google Scholar]
- Interthal, H; Pouliot, JJ; Champoux, JJ. The tyrosyl-DNA phosphodiesterase Tdp1 is a member of the phospholipase D superfamily. Proc. Nat. Acad. Sci USA 2001, 98, 12009–12014. [Google Scholar]
- Harris, MO; Stuart, JJ; Mohan, M; Nair, S; Lamb, RJ; Rohfritsch, O. Grasses and gall midges: Plant defense and insect adaptation. Annu. Rev. Entomol 2000, 48, 549–577. [Google Scholar]
- Stuart, JJ; Hatchett, JH. Cytogenetics of the Hessian fly: II. Inheritance and behavior of somatic and germ-line-limited chromosomes. J. Hered 1988, 79, 190–199. [Google Scholar]
- Andow, DA; Bentur, JS. Pedigreed crosses to estimate recessive virulence allele frequencies in natural populations of gall midges. Entomol. Exp. Appl 2010, 135, 18–36. [Google Scholar]
- Behura, SK; Nair, S; Sahu, SC; Mohan, M. An AFLP marker that differentiates biotypes of the Asian rice gall midge (Orseolia oryzae, Wood-Mason) is sex linked and also linked to avirulence. Mol. Gen. Genet 2000, 263, 328–334. [Google Scholar]
- Glenn, TC; Schable, NA. Isolating microsatellite DNA loci. Methods Enzymol 2005, 395, 202–222. [Google Scholar]
- Truett, GE. Preparation of genomic DNA from animal tissues. In DNA Sequencing II: Optimizing Preparation and Cleanup; Keileczawa, J, Ed.; Jones and Barlett Publisher: Sudbury, MA, USA, 2006; pp. 33–38. [Google Scholar]
- Excoffier, L; Laval, G; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform Online 2005, 1, 47–50. [Google Scholar]
- Raymond, M; Rousset, F. GenePop (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered 1995, 86, 248–249. [Google Scholar]



| Accession Number (GeneBank) | Locus | Repeat | Primer | Ann. Temp (°C) | MgCl2 (mM) | Size Range | n | Biotype | He | Ho | PIC | HWE (P) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HM804499 | Oosat03 | (TG)12 | *F:TTGATTGTCCCAAGGAGCAT | 60 | 1.5 | 135–148 4 | 4 | GMB1 | 0.583 | 0.350 | 0.475 | 0.0126 |
| GMB4 | 0.450 | 0.350 | 0.401 | 0.0020 | ||||||||
| R:ATTCGCGTTGTGGATTGTTT | GMB4M | 0.640 | 0.500 | 0.572 | 0.0550 | |||||||
| HM804512 | Oosat16 | (TG)15 (GAGT)6 | *F:TGTTCAGCTTGTTCAGC | 55 | 1.5 | 153–162 | 8 | GMB1 | 0.803 | 0.200 | 0.754 | 0.0000 |
| GMB4 | 0.560 | 0.100 | 0.478 | 0.0000 | ||||||||
| R:CATTGGAACGAAATTAGTGG | GMB4M | 0.810 | 0.500 | 0.770 | 0.0000 | |||||||
| HM804517 | Oosat21 | (TA)6 (TG)18 | F:CCGATTTCACTCGATGTTGTT | 53 | 3.0 | 136–150 | 2 | GMB1 | 0.518 | 0.000 | 0.365 | 0.0000 |
| GMB4 | 0.515 | 0.357 | 0.374 | 0.3150 | ||||||||
| R:TTCTAACTTGAACTCCTCATTCG | GMB4M | 0.508 | 0.286 | 0.374 | 0.1320 | |||||||
| HM804520 | Oosat24 | (AC)11(CA)5 | F:CCTCGGTCGCATCTCATATT | 52 | 3.0 | 160–200 | 4 | GMB1 | 0.634 | 1.000 | 0.501 | 0.0040 |
| GMB4 | 0.518 | 1.000 | 0.445 | 0.0000 | ||||||||
| R:CCATTCAACAGATTTGGCGTA | GMB4M | 0.645 | 1.000 | 0.548 | 0.0010 | |||||||
| HM804522 | Oosat26 | (GT)15 | *F:TGTCAGGTGGAACAGTAAATTG | 53 | 3.0 | 214–236 | 9 | GMB1 | 0.800 | 0.200 | 0.764 | 0.0000 |
| GMB4 | 0.750 | 0.450 | 0.686 | 0.0020 | ||||||||
| R:GCCTGAAGAAAGCTGAATGAA | GMB4M | 0.700 | 0.200 | 0.648 | 0.0000 | |||||||
| HM804531 | Oosat35 | (CA)11 (GA) (GA)2 | F:GCCCGTTGATTGCTTTGTAT | 51 | 1.5 | 185–220 | 5 | GMB1 | 0.796 | 0.928 | 0.704 | 0.0200 |
| GMB4 | 0.735 | 0.714 | 0.674 | 0.0000 | ||||||||
| R:TATCGTTGTCGTCGTCTTCG | GMB4M | 0.304 | 0.357 | 0.247 | 1.0000 | |||||||
| HM804532 | Oosat36 | (GT)14 | *F:CAGTTCCTTTTGTATATGCGTGAG | 51 | 1.5 | 145–174 | 10 | GMB1 | 0.780 | 0.800 | 0.738 | 0.0100 |
| GMB4 | 0.760 | 0.600 | 0.696 | 0.0070 | ||||||||
| R:GCACCCAAAATTCAATCGTT | GMB4M | 0.810 | 0.450 | 0.769 | 0.0001 | |||||||
| HM804539 | Oosat43 | (CT)9 | *F:TCGTTGGAATAGCACATTCG | 54 | 1.5 | 165–188 | 7 | GMB1 | 0.700 | 0.450 | 0.633 | 0.0200 |
| GMB4 | 0.380 | 0.400 | 0.305 | 1.0000 | ||||||||
| R:TGACGTGTCTATGCCATGTG | GMB4M | 0.620 | 0.500 | 0.557 | 0.0040 | |||||||
| HM804542 | Oosat46 | (GA)19 | F:AAATTGGCAGAGCGGAAGTA | 44 | 2 | 185–250 | 3 | GMB1 | 0.494 | 0.785 | 0.359 | 0.0360 |
| GMB4 | 0.648 | 1.000 | 0.553 | 0.0000 | ||||||||
| R:TTTCACGGCCATCACATAAG | GMB4M | 0.645 | 1.000 | 0.548 | 0.0010 | |||||||
| HM804551 | Oosat55 | (CA)2 (CA)20 | *F:CGTCGCCTTGTTGTAATATGTAAG | 55 | 1.5 | 103–135 | 10 | GMB1 | 0.780 | 0.500 | 0.744 | 0.0000 |
| GMB4 | 0.790 | 0.400 | 0.751 | 0.0000 | ||||||||
| R:ACAGCCAATTGTGTTGCTTG | GMB4M | 0.900 | 0.650 | 0.868 | 0.0000 | |||||||
| HM804555 | Oosat59 | (CA)20 | *F:CGTCGCCTTGTTTAATATG | 55 | 1.5 | 78–107 | 11 | GMB1 | 0.880 | 0.300 | 0.852 | 0.0000 |
| GMB4 | 0.580 | 0.150 | 0.534 | 0.0000 | ||||||||
| R:CCAATTGTGTTGCTTGA | GMB4M | 0.910 | 0.300 | 0.877 | 0.0000 | |||||||
| HM804574 | Oosat78 | CAG (CAA)2 (CAG)6 CAA | F:CCCAGCTCTTCGAATTCTATTG | 56 | 2 | 190–200 | 3 | GMB1 | 0.476 | 0.000 | 0.305 | 0.0000 |
| GMB4 | 0.677 | 0.000 | 0.548 | 0.0000 | ||||||||
| R:CCCGAATCATTTTGCATTGT | GMB4M | 0.349 | 0.000 | 0.305 | 0.0000 | |||||||
| HM804575 | Oosat79 | (TG)11 | *F:CGCCCTAAAGAGTCGTGAAG | 55 | 1.5 | 118–128 | 5 | GMB1 | 0.350 | 0.400 | 0.329 | 1.0000 |
| GMB4 | 0.600 | 0.550 | 0.511 | 0.5940 | ||||||||
| R:GAACCGGATGATTTGAATGG | GMB4M | 0.680 | 0.600 | 0.611 | 0.8100 | |||||||
| HM804579 | Oosat83 | (AG)15 | *F:GCGAGTCAAAACACACG | 55 | 1.5 | 105–120 | 9 | GMB1 | 0.840 | 0.600 | 0.803 | 0.0000 |
| GMB4 | 0.710 | 0.500 | 0.646 | 0.0008 | ||||||||
| R:ACACACACATATGCTCTTCC | GMB4M | 0.790 | 0.250 | 0.742 | 0.0000 | |||||||
| HM804584 | Oosat88 | (TC)15 | *F:ACAGAAGGTAGAAGGAGAGC | 55 | 1.5 | 184–192 | 6 | GMB1 | 0.770 | 0.700 | 0.709 | 0.0200 |
| GMB4 | 0.710 | 0.650 | 0.665 | 0.2210 | ||||||||
| R:AGTTGGCGATTGAGTGAG | GMB4M | 0.760 | 0.600 | 0.693 | 0.0500 |
| S/No. | Loci | Frequency of Most Frequent Alleles * (Size, bp) and Number of Alleles in Each Biotype | |||||
|---|---|---|---|---|---|---|---|
| GMB1 | GMB4 | GMB4M | |||||
| Frequency (Size) | No. of Alleles | Frequency (Size) | No. of Alleles | Frequency (Size) | No. of Alleles | ||
| 1 | Oosat03 | 0.475 (148) | 4 | 0.674 (148) | 4 | 0.525 (148) | 4 |
| 2 | Oosat16 | 0.350 (153) | 6 | 0.600 (155) | 5 | 0.325 (155) | 8 |
| 3 | Oosat21 | 0.600 (136) | 2 | 0.525 (150) | 2 | 0.525 (136) | 2 |
| 4 | Oosat24 | 0.500 (180) | 4 | 0.475 (160,180) | 4 | 0.500 (190) | 3 |
| 5 | Oosat26 | 0.350 (220) | 9 | 0.350 (219) | 6 | 0.500 (222) | 6 |
| 6 | Oosat35 | 0.375 (190) | 5 | 0.375 (200) | 4 | 0.825 (200) | 2 |
| 7 | Oosat36 | 0.350 (150) | 8 | 0.350 (153) | 5 | 0.375 (152) | 10 |
| 8 | Oosat43 | 0.425 (188) | 7 | 0.750 (168) | 2 | 0.550 (166) | 5 |
| 9 | Oosat46 | 0.625 (185) | 2 | 0.500 (250) | 3 | 0.500 (185) | 3 |
| 10 | Oosat55 | 0.400 (112) | 8 | 0.400 (125) | 9 | 0.175 (125) | 10 |
| 11 | Oosat59 | 0.200 (83,84) | 11 | 0.625 (97) | 6 | 0.150 (79,84) | 11 |
| 12 | Oosat78 | 0.750 (200) | 2 | 0.500 (210) | 3 | 0.750 (200) | 2 |
| 13 | Oosat79 | 0.800 (122) | 5 | 0.500 (122) | 3 | 0.425 (122) | 5 |
| 14 | Oosat83 | 0.250 (115,116) | 9 | 0.450 (116) | 5 | 0.350 (116) | 6 |
| 15 | Oosat88 | 0.325 (192) | 5 | 0.475 (184) | 6 | 0.300 (184) | 4 |
| S/No. | Locus | FIS | FIT | FST | Nm |
|---|---|---|---|---|---|
| 1 | Oosat03 | 0.2666 | 0.3055 | 0.0531 | 4.4625 |
| 2 | Oosat16 | 0.6238 | 0.6597 | 0.0955 | 2.3669 |
| 3 | Oosat21 | 0.4585 | 0.4643 | 0.0106 | 23.3289 |
| 4 | Oosat24 | −0.7118 | −0.3986 | 0.1830 | 1.1162 |
| 5 | Oosat26 | 0.6147 | 0.6618 | 0.1222 | 1.7961 |
| 6 | Oosat35 | −0.1095 | 0.0625 | 0.1550 | 1.3624 |
| 7 | Oosat36 | 0.1965 | 0.2851 | 0.1103 | 2.0168 |
| 8 | Oosat43 | 0.1922 | 0.3466 | 0.1912 | 1.0578 |
| 9 | Oosat46 | −0.6058 | −0.4808 | 0.0779 | 2.9611 |
| 10 | Oosat55 | 0.3608 | 0.4178 | 0.0892 | 2.5526 |
| 11 | Oosat59 | 0.6769 | 0.7164 | 0.1223 | 1.7948 |
| 12 | Oosat78 | 1.0000 | 1.0000 | 0.1772 | 1.1610 |
| 13 | Oosat79 | 0.0380 | 0.1296 | 0.0952 | 2.3753 |
| 14 | Oosat83 | 0.4114 | 0.4348 | 0.0396 | 6.0628 |
| 15 | Oosat88 | 0.1116 | 0.1551 | 0.0489 | 4.8598 |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bentur, J.S.; Sinha, D.K.; Padmavathy, C.; Revathy, C.; Muthulakshmi, M.; Nagaraju, J. Isolation and Characterization of Microsatellite Loci in the Asian Rice Gall Midge (Orseolia oryzae) (Diptera: Cecidomyiidae). Int. J. Mol. Sci. 2011, 12, 755-772. https://doi.org/10.3390/ijms12010755
Bentur JS, Sinha DK, Padmavathy C, Revathy C, Muthulakshmi M, Nagaraju J. Isolation and Characterization of Microsatellite Loci in the Asian Rice Gall Midge (Orseolia oryzae) (Diptera: Cecidomyiidae). International Journal of Molecular Sciences. 2011; 12(1):755-772. https://doi.org/10.3390/ijms12010755
Chicago/Turabian StyleBentur, Jagadish S., Deepak Kumar Sinha, Ch. Padmavathy, Charagonda Revathy, Mayandi Muthulakshmi, and Javaregowda Nagaraju. 2011. "Isolation and Characterization of Microsatellite Loci in the Asian Rice Gall Midge (Orseolia oryzae) (Diptera: Cecidomyiidae)" International Journal of Molecular Sciences 12, no. 1: 755-772. https://doi.org/10.3390/ijms12010755