Experimental Construction of BMP2 and VEGF Gene Modified Tissue Engineering Bone in Vitro

Abstract
:1. Introduction
2. Results and Discussion
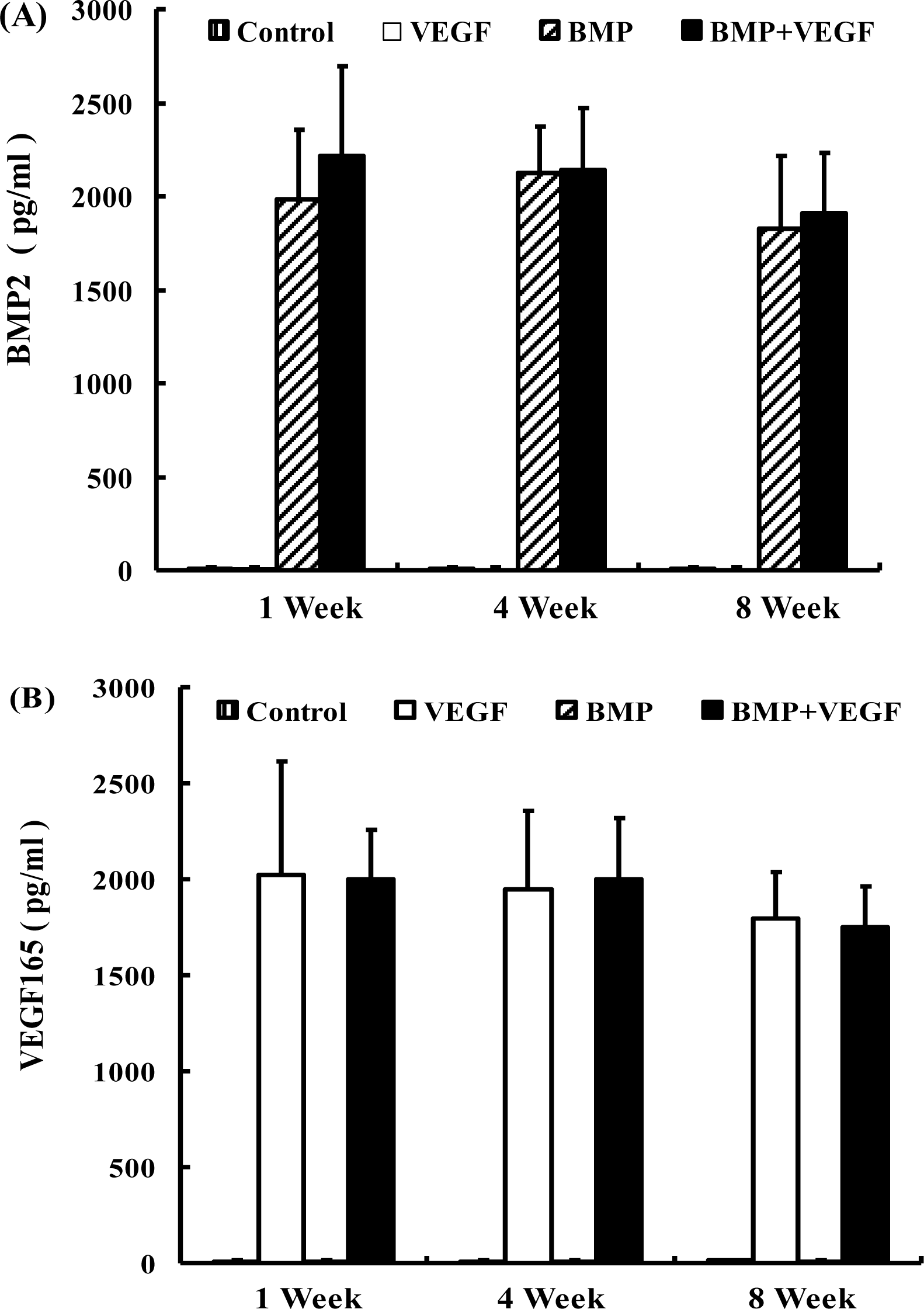
2.1. ELISA
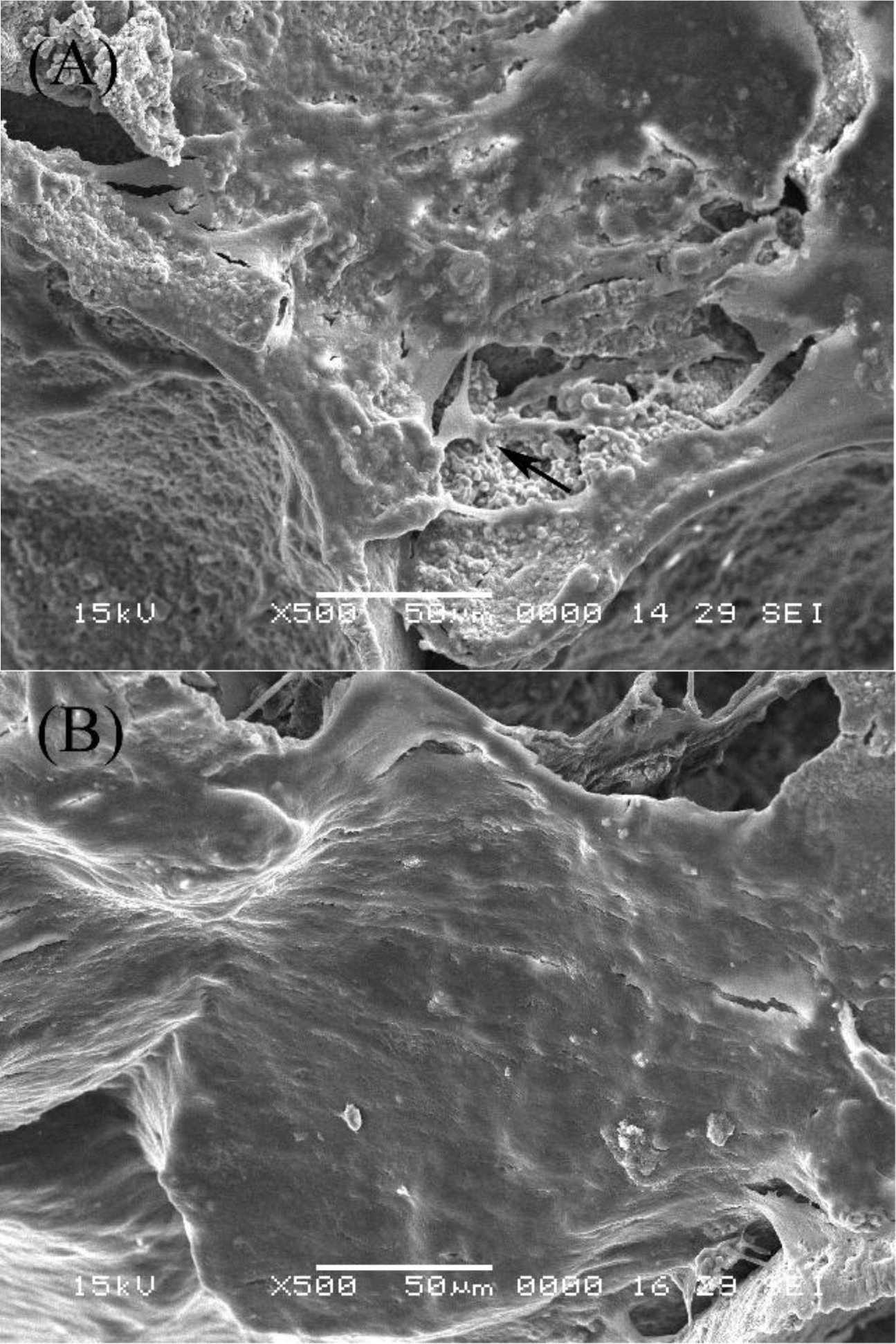
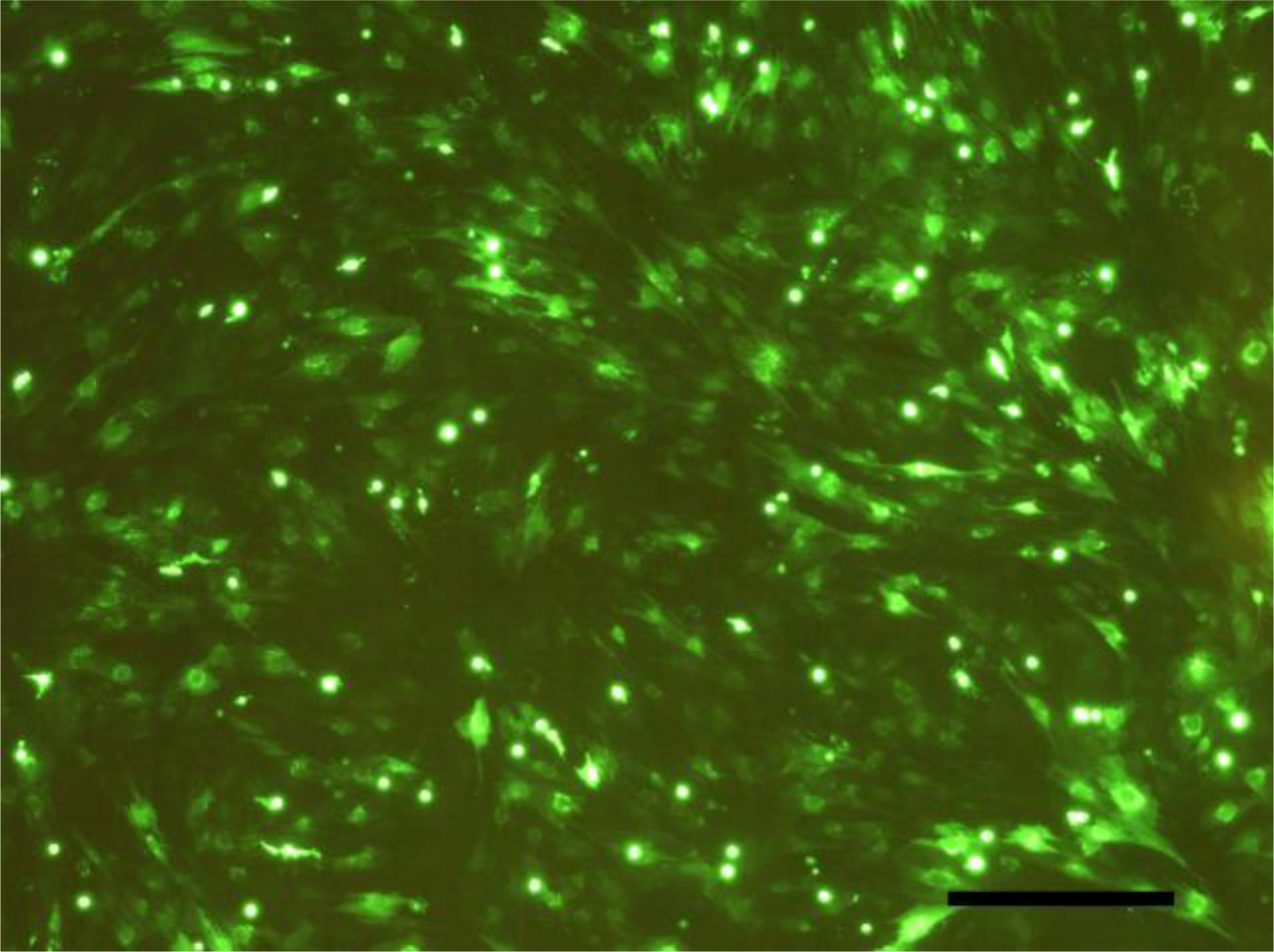
2.2. SEM Examination
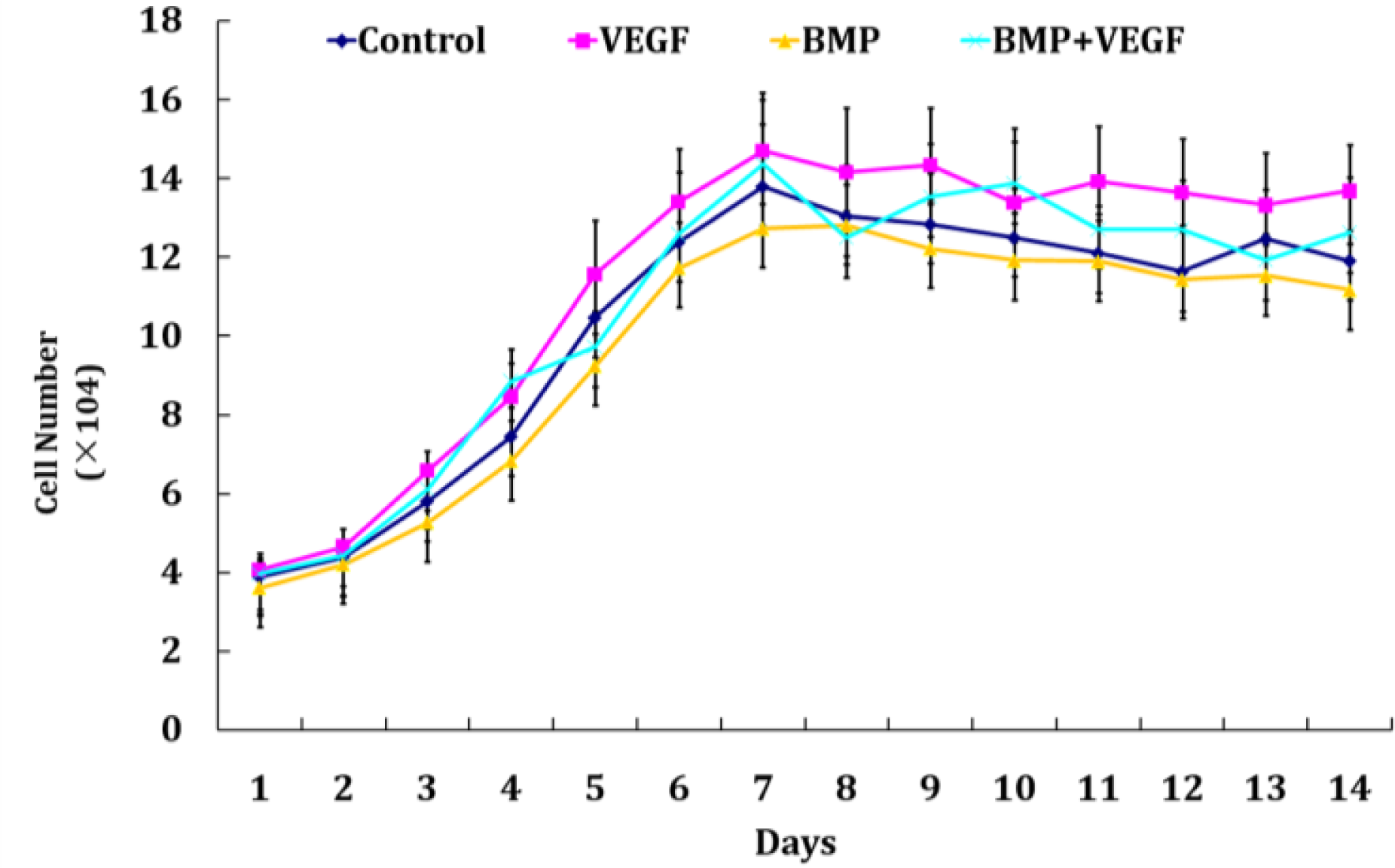
2.3. Hoechst DNA Assay
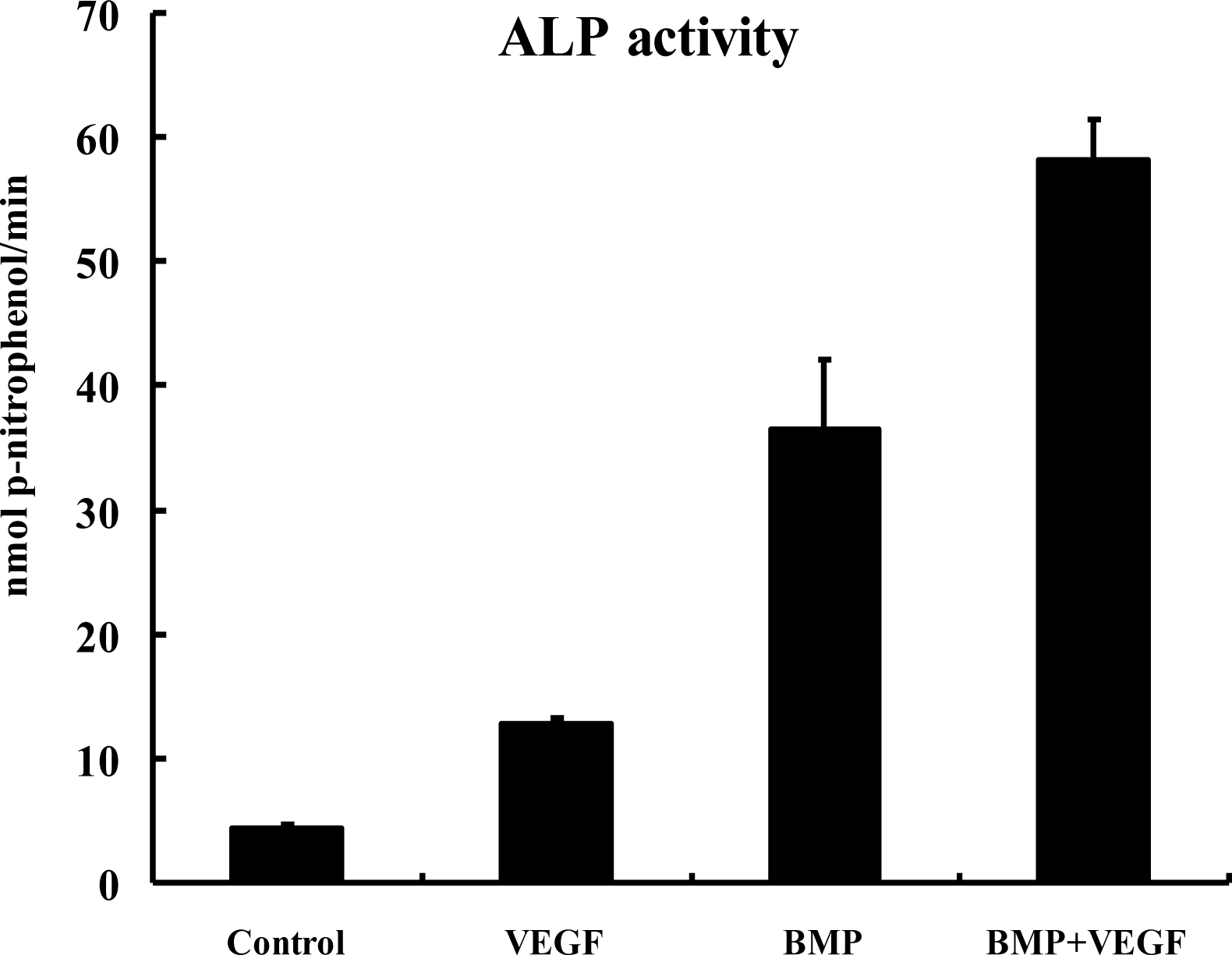
2.4. ALP Activity Assay
3. Experimental Section
3.1. MSCs Isolation and Expansion
3.3. Enzyme Linked Immunoabsorbent Assay (ELISA)
3.4. Preparation of Porous β-TCP Scaffolds and Cell Seeding
3.5. Scanning Electron Microscopy (SEM)
3.6. Hoechst DNA Assay
3.7. ALP Activity Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Jiang, XQ; Chen, JG; Gittens, S; Chen, CJ; Zhang, XL; Zhang, ZY. The ectopic study of tissue-engineered bone with hBMP-4 gene modified bone marrow stromal cells in rabbits. Chin. Med. J 2005, 118, 281–288. [Google Scholar]
- Schek, RM; Wilke, EN; Hollister, SJ; Krebsbach, PH. Combined use of designed scaffolds and adenoviral gene therapy for skeletal tissue engineering. Biomaterials 2006, 27, 1160–1166. [Google Scholar]
- Roldán, JC; Detsch, R; Schaefer, S; Chang, E; Kelantan, M; Waiss, W; Reichert, TE; Gurtner, GC; Deisinger, U. Bone formation and degradation of a highly porous biphasic calcium phosphate ceramic in presence of BMP-7, VEGF and mesenchymal stem cells in an ectopic mouse model. J. Craniomaxillofac. Surg 2010, 38(6), 423–430. [Google Scholar]
- Xu, XL; Lou, J; Tang, T; Ng, KW; Zhang, dJ; Yu, C; Dai, K. Evaluation of different scaffolds for BMP-2 genetic orthopedic tissue engineering. J. Biomed. Mater. Res. B. Appl. Biomater 2005, 75, 289–303. [Google Scholar]
- Dai, KR; Xu, XL; Tang, TT; Zhu, ZA; Yu, CF; Lou, JR; Zhang, XL. Repairing of goat tibial bone defects with BMP-2 gene-modified tissue-engineered bone. Calcif. Tissue Int 2005, 77, 55–61. [Google Scholar]
- Jeon, O; Song, SJ; Yang, HS; Bhang, SH; Kang, SW; Lee, JH; Kim, BS. Long-term delivery enhances in vivo osteogenic efficacy of bone morphogenetic protein-2 compared to short-term delivery. Biochem. Biophys. Res. Commun 2008, 369(2), 774–780. [Google Scholar]
- Kiyozuka, Y; Miyazaki, H; Yoshizawa, K; Senzaki, H; Yamamoto, D; Inoue, K; Bessho, K; Okubo, Y; Kusumoto, K; Tsubura, A. An autopsy case of malignant mesothelioma with osseous and cartilaginous differentiation: Bone morphogenetic protein-2 in mesothelial cells and its tumor. Dig. Dis. Sci 1999, 44, 1626–1631. [Google Scholar]
- Peng, H; Wright, V; Usas, A; Gearhart, B; Shen, HC; Cummins, J; Huard, J. Synergistic enhancement of bone formation and healing by stem cell-expressed VEGF and bone morphogenetic protein-4. J. Clin. Invest 2002, 110, 751–759. [Google Scholar]
- Huang, YC; Kaigler, D; Rice, KG; Krebsbach, PH; Mooney, DJ. Combined angiogenic and osteogenic factor delivery enhances bone marrow stromal cell-driven bone regeneration. J. Bone. Miner. Res 2005, 20, 848–857. [Google Scholar]
- Lieberman, JR; Ghivizzani, SC; Evans, CH. Gene transfer approaches to the healing of bone and cartilage. Mol. Ther 2002, 6, 141–147. [Google Scholar]
- McMahon, JM; Conroy, S; Lyons, M; Greiser, U; O’shea, C; Strappe, P; Howard, L; Murphy, M; Barry, F; O’Brien, T. Gene transfer into rat mesenchymal stem cells: A comparative study of viral and nonviral vectors. Stem Cell. Dev 2006, 15, 87–96. [Google Scholar]
- Naldini, L; Blomer, U; Gage, FH; Trono, D; Verma, IM. Efficient transfer, integration, and sustained long-term expression of the transgene in adult rat brains injected with a lentiviral vector. Proc. Natl. Acad. Sci. USA 1996, 93, 11382–11388. [Google Scholar]
- Lu, W; Zhou, X; Hong, B; Liu, J; Yue, Z. Suppression of invasion in human U87 glioma cells by adenovirus-mediated co-transfer of TIMP-2 and PTEN gene. Cancer Lett 2004, 214, 205–213. [Google Scholar]
- Ojima, T; Iwahashi, M; Nakamura, M; Matsuda, K; Naka, T; Nakamori, M; Ueda, K; Ishida, K; Yamaue, H. The boosting effect of co-transduction with cytokine genes on cancer vaccine therapy using genetically modified dendritic cells expressing tumor-associated antigen. Int. J. Oncol 2006, 28, 947–953. [Google Scholar]
- Tsuda, H; Wada, T; Yamashita, T; Hamada, H. Enhanced osteoinduction by mesenchymal stem cells transfected with a fiber-mutant adenoviral BMP2 gene. J. Gene. Med 2005, 7, 1322–1334. [Google Scholar]
- Wang, WG; Lou, SQ; Ju, XD; Xia, K; Xia, JH. In vitro chondrogenesis of human bone marrow-derived mesenchymal progenitor cells in monolayer culture: Activation by transfection with TGF-beta2. Tissue Cell 2003, 35, 69–77. [Google Scholar]
- Sugiyama, O; An, DS; Kung, SP; Feeley, BT; Gamradt, S; Liu, NQ; Chen, IS; Lieberman, JR. Lentivirus-mediated gene transfer induces long-term transgene expression of BMP-2 in vitro and new bone formation in vivo. Mol. Ther 2005, 11, 390–398. [Google Scholar]
- Pereboeva, L; Komarova, S; Mikheeva, G; Krasnykh, V; Curiel, DT. Approaches to utilize mesenchymal progenitor cells as cellular vehicles. Stem Cells 2003, 21, 389–404. [Google Scholar]
- Wilson, JM. Adenoviruses as gene-delivery vehicles. N. Engl. J. Med 1996, 334, 1185–1187. [Google Scholar]
- Hsu, WK; Sugiyama, O; Park, SH; Conduah, A; Feeley, BT; Liu, NQ; Krenek, L; Virk, MS; An, DS; Chen, IS; Lieberman, JR. Lentiviral-mediated BMP-2 gene transfer enhances healing of segmental femoral defects in rats. Bone 2007, 40, 931–938. [Google Scholar]
- Mao, X; Chu, CL; Mao, Z; Wang, JJ. The development and identification of constructing tissue engineered bone by seeding osteoblasts from differentiated rat marrow stromal stem cells onto three-dimensional porous nano-hydroxylapatite bone matrix in vitro. Tissue Cell 2005, 37, 349–357. [Google Scholar]
- Boden, SD; Zdeblick, T; Sandhu, HS; Heim, S. The use of rhBMP-2 in interbody fusion cages: Definitive evidence of osteoinduction in humans—A preliminary report. Spine 2000, 5, 376–381. [Google Scholar]
- Banwart, JC; Asher, MA; Hassanein, RS. Iliac crest bone graft harvest donor site morbidity: A statistical evaluation. Spine 1995, 20, 1055–1060. [Google Scholar]
- Rose, FR; Oreffo, RO. Bone tissue engineering: Hope vs hype. Biochem. Biophys. Res. Commun 2002, 292, 1–7. [Google Scholar]
- Liu, GP; Zhao, L; Cui, L; Liu, W; Cao, YL. Tissue-engineered bone formation using human bone marrow stromal cells and novel β-tricalcium phosphate. Biomed. Mater 2007, 2, 78–86. [Google Scholar]
- Vindigni, V; Cortivo, R; Iacobellis, L; Abatangelo, G; ZavanInt, B. Hyaluronan benzyl ester as a scaffold for tissue engineering. J. Mol. Sci 2009, 10, 2972–2985. [Google Scholar]
- Dull, T; Zufferey, R; Kelly, M; Mandel, RJ; Nguyen, M; Trono, D; Naldini, L. A third-generation lentivirus vector with a conditional packaging system. J. Virol 1998, 72, 8463–8471. [Google Scholar]
- Zufferey, R; Dull, T; Mandel, RJ; Bukovsky, A; Quiroz, D; Naldini, L; Trono, D. Self-inactivating lentivirus vector for safe and efficient in vivo gene delivery. J. Virol 1998, 72, 9873–9880. [Google Scholar]
- Spinella-Jaegle, S; Roman-Roman, S; Faucheu, C; Dunn, FW; Kawai, S; Gallea, S; Stiot, V; Blanchet, AM; Courtois, B; Baron, R; Rawadi, G. Opposite effects of bone morphogenetic protein-2 and transforming growth factor-beta1 on osteoblast differentiation. Bone 2001, 29, 323–330. [Google Scholar]
- Wu, G; He, X; Yang, Z; Guo, L. Influence on the osteogenic activity of the human bone marrow mesenchymal stem cells transfected by liposome-mediated recombinant plasmid pIRES-hBMP2-hVEGF165in vitro. Ann. Plast. Surg 2010, 65(1), 80–84. [Google Scholar]
- Lee, YS; Choi, CW; Kim, JJ; Ganapathi, A; Udayakumar, R; Kim, SC. Determination of mineral content in methanolic safflower (Carthamus tinctorius L.) seed extract and its effect on osteoblast markers. Int. J. Mol. Sci 2009, 10, 292–305. [Google Scholar]
- Musgrave, DS; Pruchnic, R; Bosch, P; Ziran, BH; Whalen, J; Huard, J. Human skeletal muscle cells in ex vivo gene therapy to deliver bone morphogenetic protein-2. J. Bone Joint Surg. Br 2002, 84, 120–127. [Google Scholar]
- Street, J; Bao, M; deGuzman, L; Bunting, S; Peale, FV, Jr; Ferrara, N; Steinmetz, H; Hoeffel, J; Cleland, JL; Daugherty, A; van Bruggen, N; Redmond, HP; Carano, RA; Filvaroff, EH. Vascular endothelial growth factor stimulates bone repair by promoting angiogenesis and bone turnover. Proc. Natl. Acad. Sci. USA 2002, 99, 9656–9661. [Google Scholar]
- Midy, V; Plouet, J. Vasculotropin/vascular endothelial growth factor induces differentiationin cultured osteoblasts. Biochem. Biophys. Res. Commun 1994, 199, 380–386. [Google Scholar]
- Deckers, MM; Karperien, M; van der Bent, C; Yamashita, T; Papapoulos, SE; Lowik, CW. Expression of vascular endothelial growth factors and their receptors during osteoblast differentiation. Endocrinology 2000, 141, 1667–1674. [Google Scholar]
- Yeh, LC; Lee, JC. Co-transfection with the osteogenic protein (OP)-1 gene and the insulin-like growth factor (IGF)-I gene enhanced osteoblastic cell differentiation. Biochim. Biophys. Acta 2006, 1763, 57–63. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Primers |
|---|
| VEGF165 |
| 5′-CGGAATTCGCCACCATGAACTTTCTGCTGTCTTGGGTGC-3′ (forward) |
| 5′-CGCGGATCCTCACCGCCTCGGCTTGTCACATC-3′ (reverse) |
| BMP2 |
| 5′-GGAATTCGCCACCTGCGGTCTCCTAAAGGTC-3′ (forward) |
| 5′-CGGGATCCTTGCTGTACTAGCGACACCCAC-3′ (reverse) |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jiang, J.; Fan, C.-Y.; Zeng, B.-F. Experimental Construction of BMP2 and VEGF Gene Modified Tissue Engineering Bone in Vitro. Int. J. Mol. Sci. 2011, 12, 1744-1755. https://doi.org/10.3390/ijms12031744
Jiang J, Fan C-Y, Zeng B-F. Experimental Construction of BMP2 and VEGF Gene Modified Tissue Engineering Bone in Vitro. International Journal of Molecular Sciences. 2011; 12(3):1744-1755. https://doi.org/10.3390/ijms12031744
Chicago/Turabian StyleJiang, Jia, Cun-Yi Fan, and Bing-Fang Zeng. 2011. "Experimental Construction of BMP2 and VEGF Gene Modified Tissue Engineering Bone in Vitro" International Journal of Molecular Sciences 12, no. 3: 1744-1755. https://doi.org/10.3390/ijms12031744