Calpain Activator Dibucaine Induces Platelet Apoptosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Reagents
2.2. Platelet Preparation
2.3. Platelet Aggregation Assay
2.4. ΔΨm Measurement
2.5. PS Externalization Assay
2.6. Platelet Surface Staining
2.7. Western Blot Analysis
2.8. Confocal Microscopy
2.9. Caspase-3 Activity Assay
3. Results and Discussion
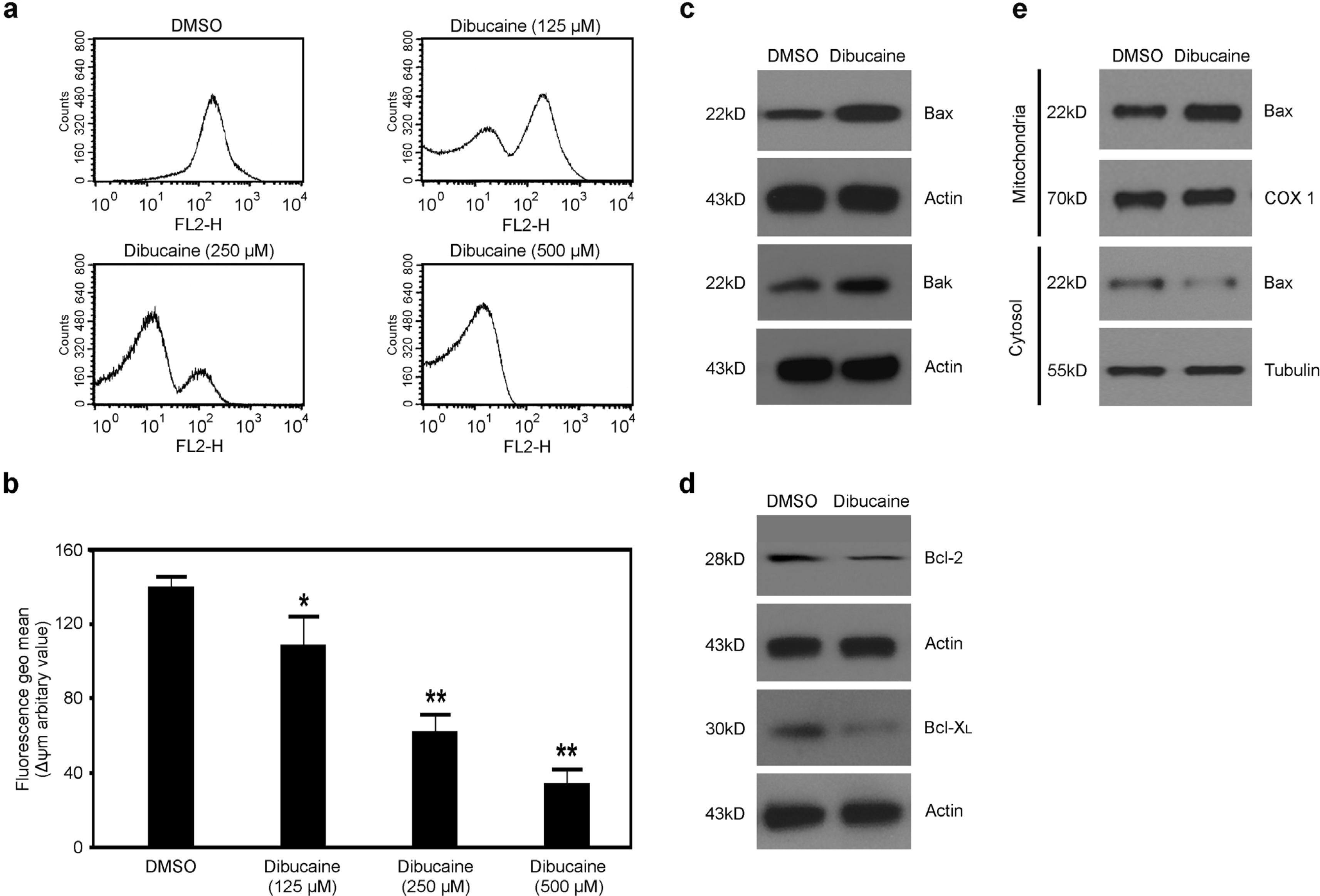
3.1. Dibucaine Dose-Dependently Induces Depolarization of ΔΨm in Platelets
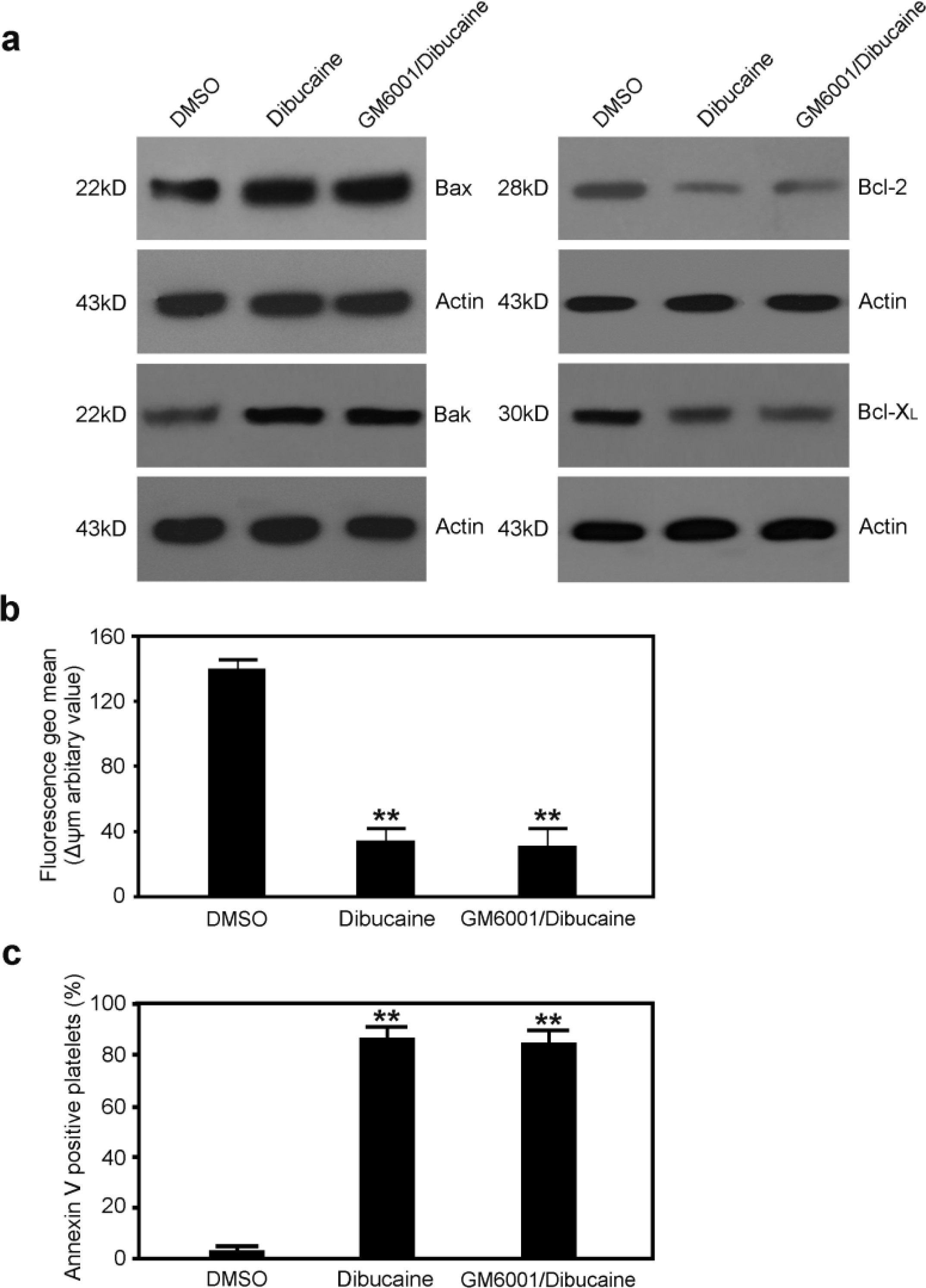
3.2. Dibucaine Elicits Up-regulation of Bax and Bak, and Down-regulation of Bcl-2 and Bcl-XL in Platelets
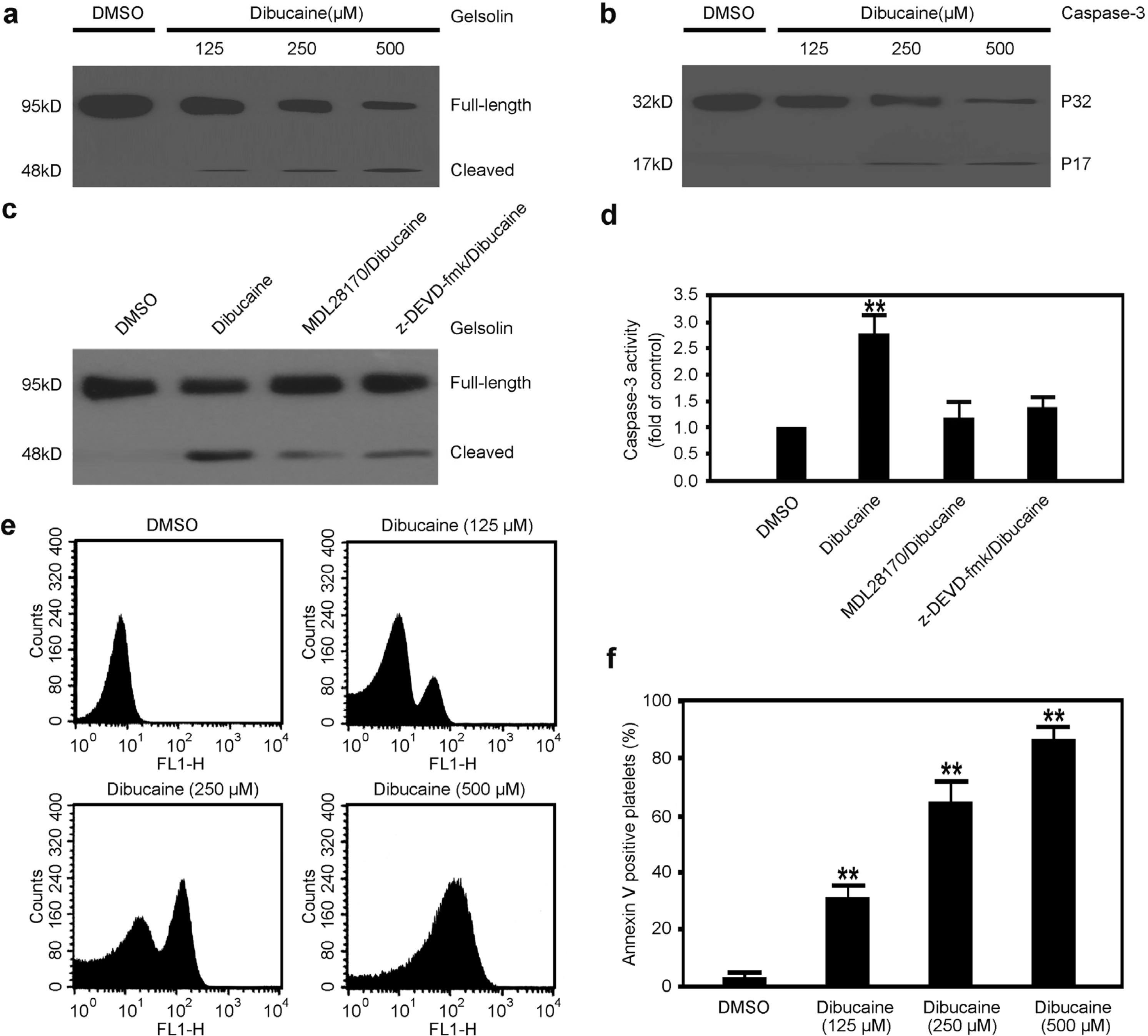
3.3. Caspase-3 Is Activated in Platelets Treated with Dibucaine
3.4. Dibucaine Induces PS Exposure in Platelets
3.5. Dibucaine-Induced Platelet Apoptosis Is Independent of GPIbα Shedding
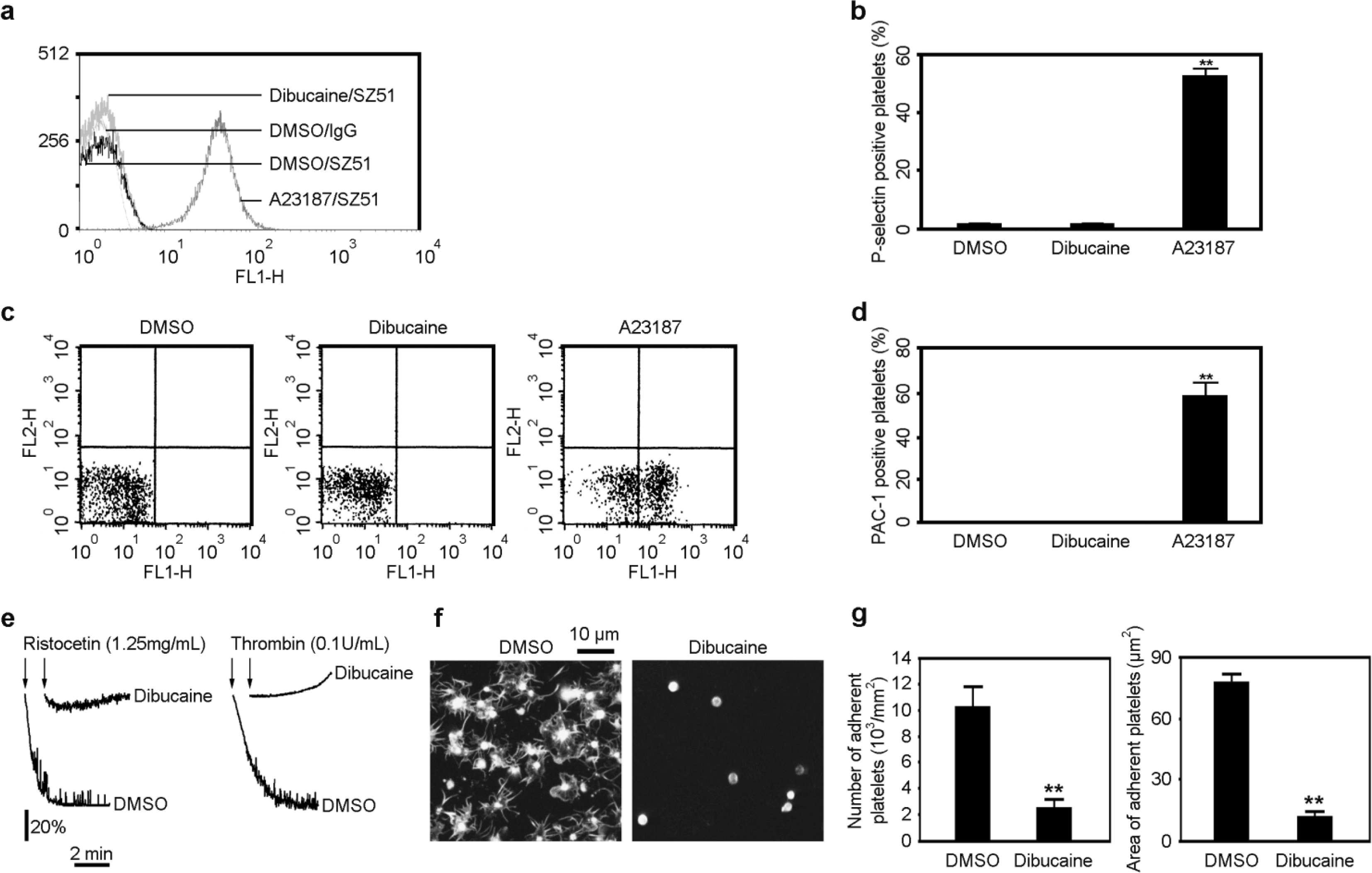
3.6. Dibucaine Does Not Induce Platelet Activation
3.7. Dibucaine Impairs Platelet Function
4. Conclusions
ijms-12-02125-s001.pdf
Acknowledgments
References
- Hato, T; Pampori, N; Shattil, SJ. Complementary roles for receptor clustering and conformational change in the adhesive and signaling functions of integrin αIIbβ3. J. Cell Biol 1998, 141, 1685–1695. [Google Scholar]
- Azam, M; Andrabi, SS; Sahr, KE; Kamath, L; Kuliopulos, A; Chishti, AH. Disruption of the mouse μ-calpain gene reveals an essential role in platelet function. Mol. Cell Biol 2001, 21, 2213–2220. [Google Scholar]
- Croce, K; Flaumenhaft, R; Rivers, M; Furie, B; Furie, BC; Herman, IM; Potter, DA. Inhibition of calpain blocks platelet secretion, aggregation, and spreading. J. Biol. Chem 1999, 274, 36321–36327. [Google Scholar]
- Fox, JEB; Reynolds, CC; Phillips, DR. Calcium-dependent proteolysis occurs during platelet aggregation. J. Biol. Chem 1983, 258, 9973–9981. [Google Scholar]
- Wang, Z; Shi, Q; Yan, R; Liu, G; Zhang, W; Dai, K. The role of calpain in the regulation of ADAM-17-dependent GPIbα ectodomain shedding. Arch. Biochem. Biophys 2010, 495, 136–143. [Google Scholar]
- Murphy, WG; Moore, J; Kelton, JG. Calcium-dependent cysteine protease activity in the sera of patients with thrombotic thrombocytopenic purpura. Blood 1987, 70, 1683–1687. [Google Scholar]
- Kelton, JG; Warkentin, TE; Hayward, CPM; Murphy, WG; Moore, JC. Calpain activity in patients with thrombotic thromocytopenic purpur is associated with platelet microparticles. Blood 1992, 80, 2246–2251. [Google Scholar]
- Hiwasa, T. Induction of apoptosis by a calpain stimulator, ONO-3403. Apoptosis 1996, 1, 75–80. [Google Scholar]
- Gil-Parrado, S; Fernandez-Montalvan, A; Assfalg-Machleidt, I; Popp, O; Bestvater, F; Holloschi, A; Knoch, TA; Auerswald, EA; Welsh, K; Reed, JC; et al. Ionomycin-activated calpain triggers apoptosis. J. Biol. Chem 2002, 277, 27217–27226. [Google Scholar]
- Squier, MK; Miller, AC; Malkinson, AM; Cohen, JJ. Calpain activation in apoptosis. J. Cell Physiol 1994, 159, 229–237. [Google Scholar]
- Leytin, V; Allen, DJ; Mykhaylov, S; Lyubimov, E; Freedman, J. Thrombin-triggered platelet apoptosis. J. Thromb. Haemost 2006, 4, 2656–2663. [Google Scholar]
- Leytin, V; Freedman, J. Platelet apoptosis in stored platelet concentrations and other models. Transfus. Apher. Sci 2003, 28, 285–295. [Google Scholar]
- Vanags, DM; Orrenius, S; Aguilar-Santelises, M. Alterations in Bcl-2/Bax protein levels in platelets form part of an ionomycin-induced process that resembles apoptosis. Br. J. Haematol 1997, 99, 824–831. [Google Scholar]
- Leytin, V; Allen, DJ; Mykhaylov, S; Mis, L; Lyubimov, EV; Garvey, B; Freedman, J. Pathologic high shear stress induces apoptosis events in human platelets. Biochem. Biophys. Res. Commun 2004, 320, 303–310. [Google Scholar]
- Mason, KD; Carpinelli, MR; Fletcher, JI; Collinge, JE; Hilton, AA; Ellis, S; Kelly, PN; Ekert, PG; Metcalf, D; Roberts, AW; Huang, DCS; Kile, BT. Programmed anuclear cell death delimits platelet life span. Cell 2007, 128, 1173–1186. [Google Scholar]
- Leytin, V; Allen, DJ; Mutlu, A; Mykhaylov, S; Lyubimov, E; Freedman, J. Platelet activation and apoptosis are different phenomena: Evidence from the sequential dynamics and the magnitude of responses during platelet storage. Br. J. Haematol 2008, 142, 480–501. [Google Scholar]
- Li, S; Wang, Z; Liao, Y; Zhang, W; Shi, Q; Yan, R; Ruan, C; Dai, K. The glycoprotein Ib alpha-von Willebrand factor interaction induces platelet apoptosis. J. Thromb. Haemost 2010, 8, 341–350. [Google Scholar]
- Wang, Z; Li, S; Shi, Q; Yan, R; Liu, G; Dai, K. Calmodulin antagonists induce platelet apoptosis. Thromb. Res 2010, 125, 340–350. [Google Scholar]
- Wang, Z; Shi, Q; Li, S; Du, J; Liu, J; Dai, K. Hyperthermia induces platelet apoptosis and glycoprotein Ib alpha ectodomain shedding. Platelets 2010, 21, 229–237. [Google Scholar]
- Jobe, SM; Wilson, KM; Leo, L; Raimondi, A; Molkentin, JD; Lentz, SR; Paola, JD. Critical role for the mitochondrial permeability transition pore and cyclophilin D in platelet activation and thrombosis. Blood 2008, 111, 1257–1265. [Google Scholar]
- Schoenwaelder, SM; Yuan, Y; Josefsson, EC; White, MJ; Yao, Y; Mason, KD; O’Reilly, LA; Henley, KJ; Ono, A; Hsiao, S; et al. Two distinct pathways regulate platelet phosphatidylserine exposure and procoagulant function. Blood 2009, 114, 663–666. [Google Scholar]
- Shcherbina, A; Remold-O’Donnell, E. Role of caspase in a subset of human platelet activation responses. Blood 1999, 93, 4222–4231. [Google Scholar]
- Wolf, BB; Green, DR. Suicidal tendencies: Apoptotic cell death by caspase family proteinases. J. Biol. Chem 1999, 274, 20049–20052. [Google Scholar]
- Wolf, BB; Goldstein, JC; Stennicke, HR; Beere, H; Amarante-Mendes, GP; Salvesen, GS; Green, DR. Calpain functions in a caspase-independent manner to promote apoptosis-like events during platelet activation. Blood 1999, 94, 1683–1692. [Google Scholar]
- Lin, KH; Hsiao, G; Shih, CM; Chou, DS; Sheu, JR. Mechanisms of resveratrol-induced platelet apoptosis. Cardiovasc. Res 2009, 83, 575–585. [Google Scholar]
- Wang, Q; Luo, W; Zhang, W; Dai, Z; Chen, Y; Chen, J. Iron supplementation protects against lead-induced apoptosis through MAPK pathway in weanling rat cortex. Neurotoxicology 2007, 28, 850–859. [Google Scholar]
- Oda, A; Druker, BJ; Ariyoshi, H; Smith, M; Salzman, EW. pp60src is an endogenous substrate for calpain in human blood platelets. J. Biol. Chem 1993, 268, 12603–12608. [Google Scholar]
- Nakagawa, T; Yuan, J. Cross-talk between two cysteine protease families. J. Cell Biol 2000, 150, 887–894. [Google Scholar]




© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, W.; Liu, J.; Sun, R.; Zhao, L.; Du, J.; Ruan, C.; Dai, K. Calpain Activator Dibucaine Induces Platelet Apoptosis. Int. J. Mol. Sci. 2011, 12, 2125-2137. https://doi.org/10.3390/ijms12042125
Zhang W, Liu J, Sun R, Zhao L, Du J, Ruan C, Dai K. Calpain Activator Dibucaine Induces Platelet Apoptosis. International Journal of Molecular Sciences. 2011; 12(4):2125-2137. https://doi.org/10.3390/ijms12042125
Chicago/Turabian StyleZhang, Weilin, Jun Liu, Ruichen Sun, Lili Zhao, Juan Du, Changgeng Ruan, and Kesheng Dai. 2011. "Calpain Activator Dibucaine Induces Platelet Apoptosis" International Journal of Molecular Sciences 12, no. 4: 2125-2137. https://doi.org/10.3390/ijms12042125