Molecular Characterization of Tob1 in Muscle Development in Pigs

Abstract
:1. Introduction
2. Results and Discussion
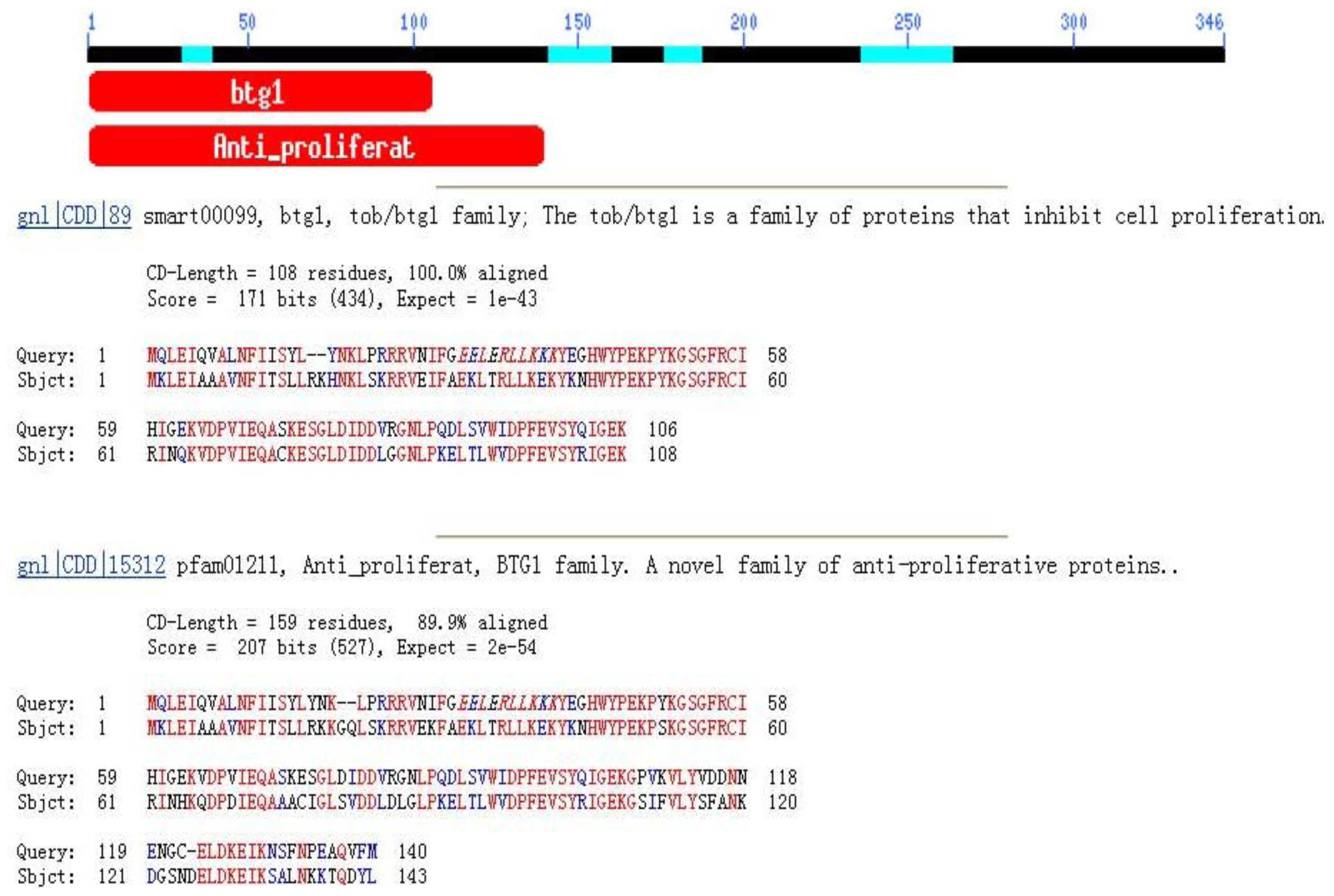
2.1. Tob1 mRNA Sequences Analysis
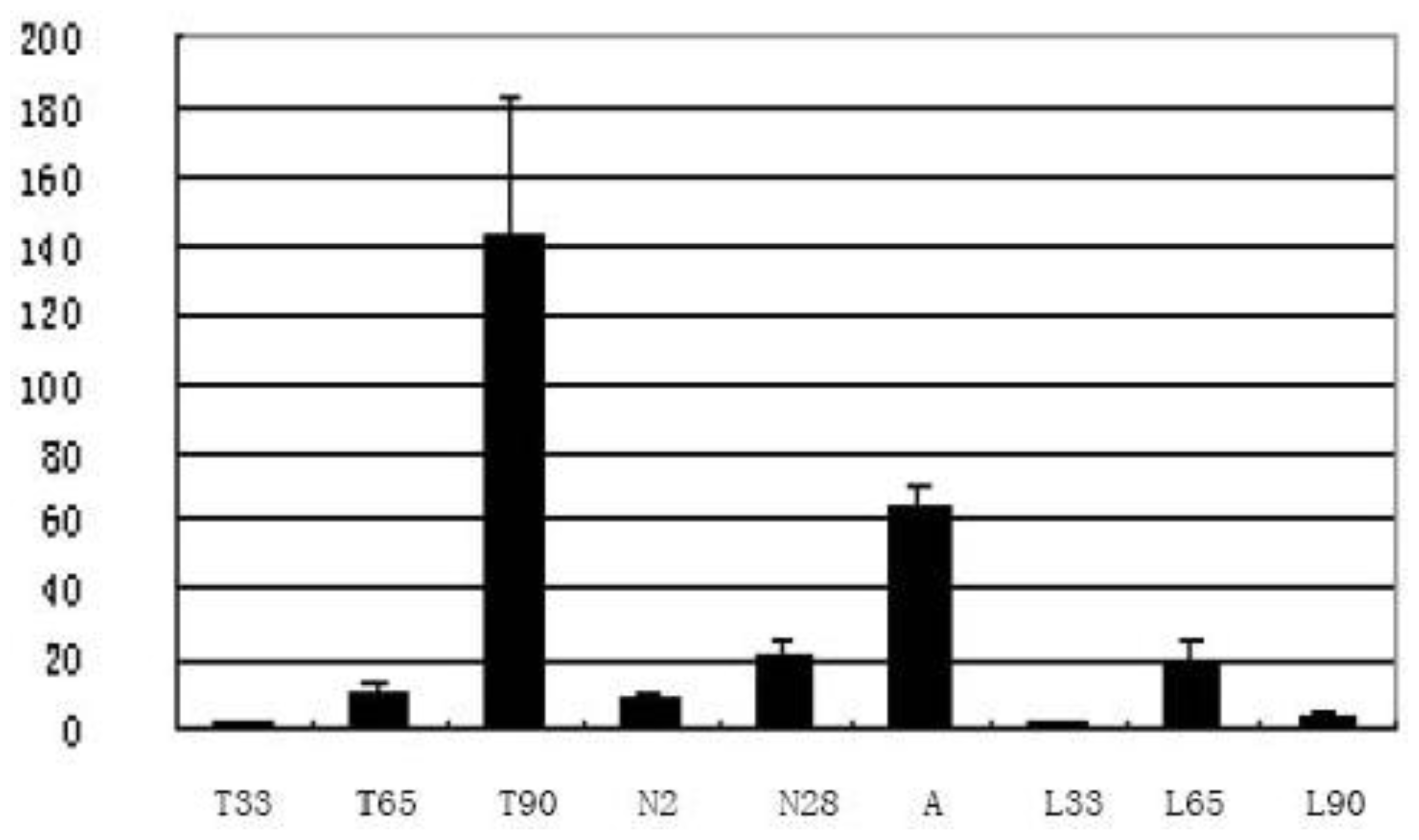
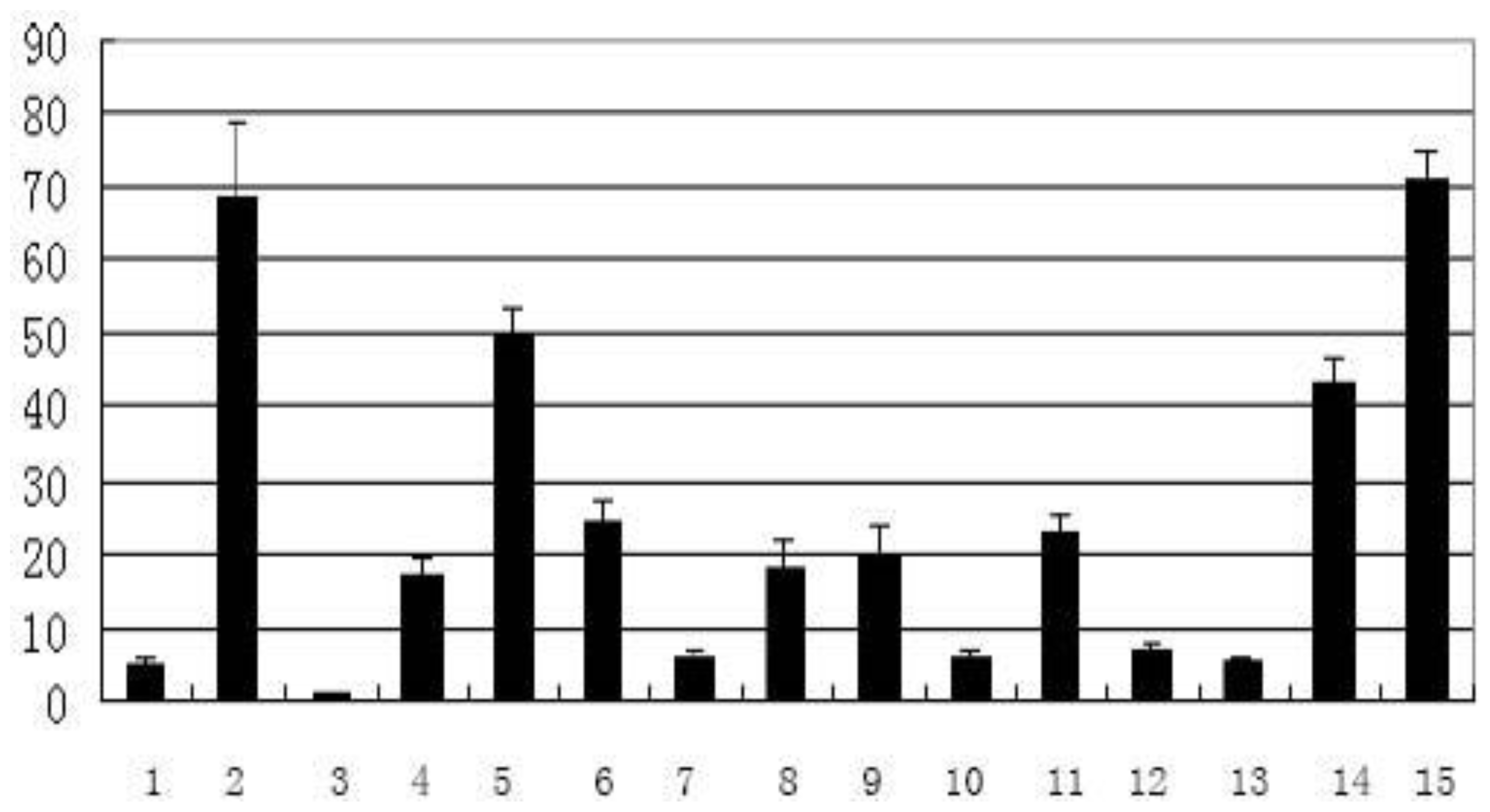
2.2. Temporal and Spatial Expression
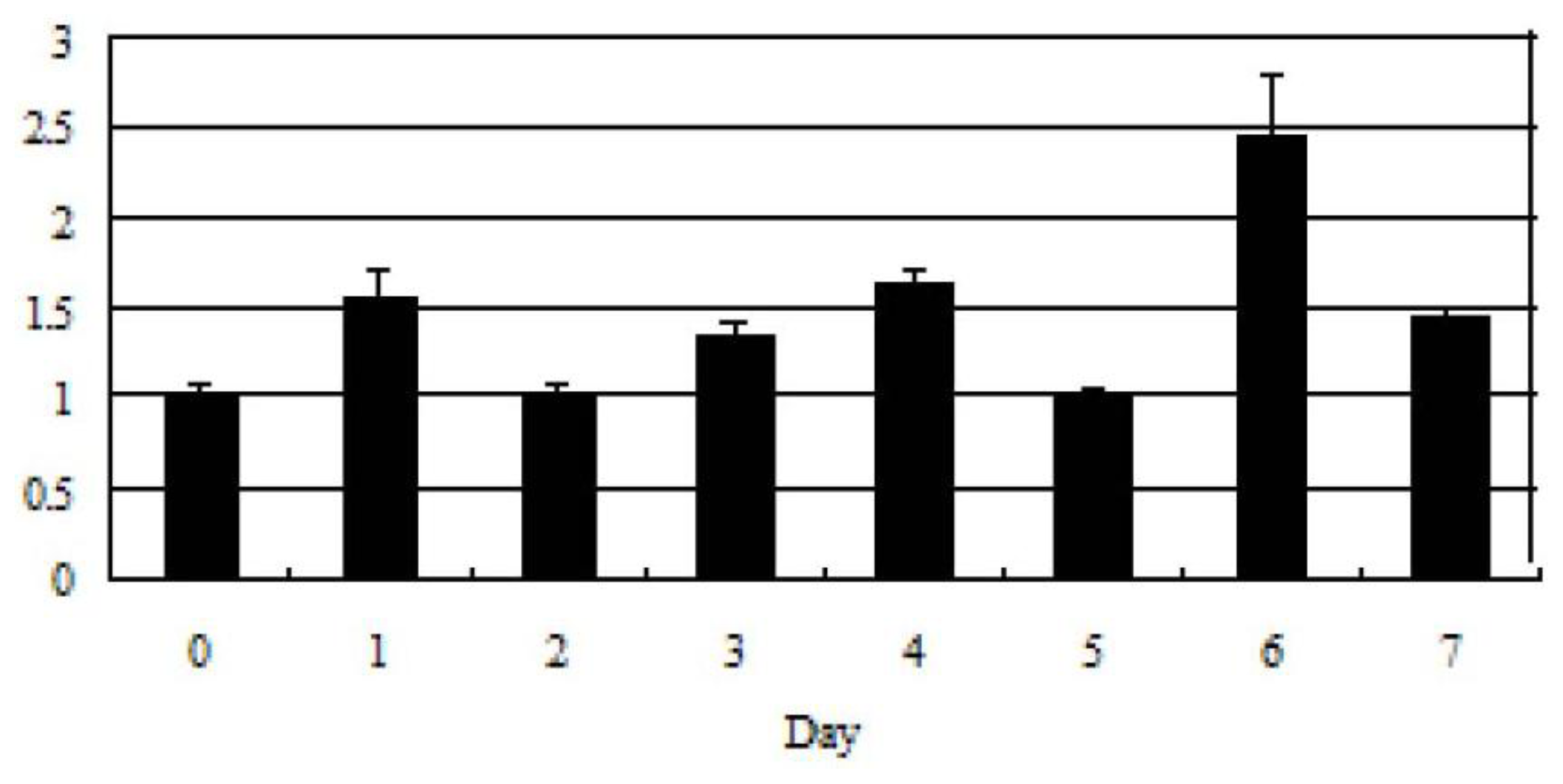
2.3. Tob1 Expression During C2C12 Differentiation Time
2.4. Chromosome Mapping Using IMpRH
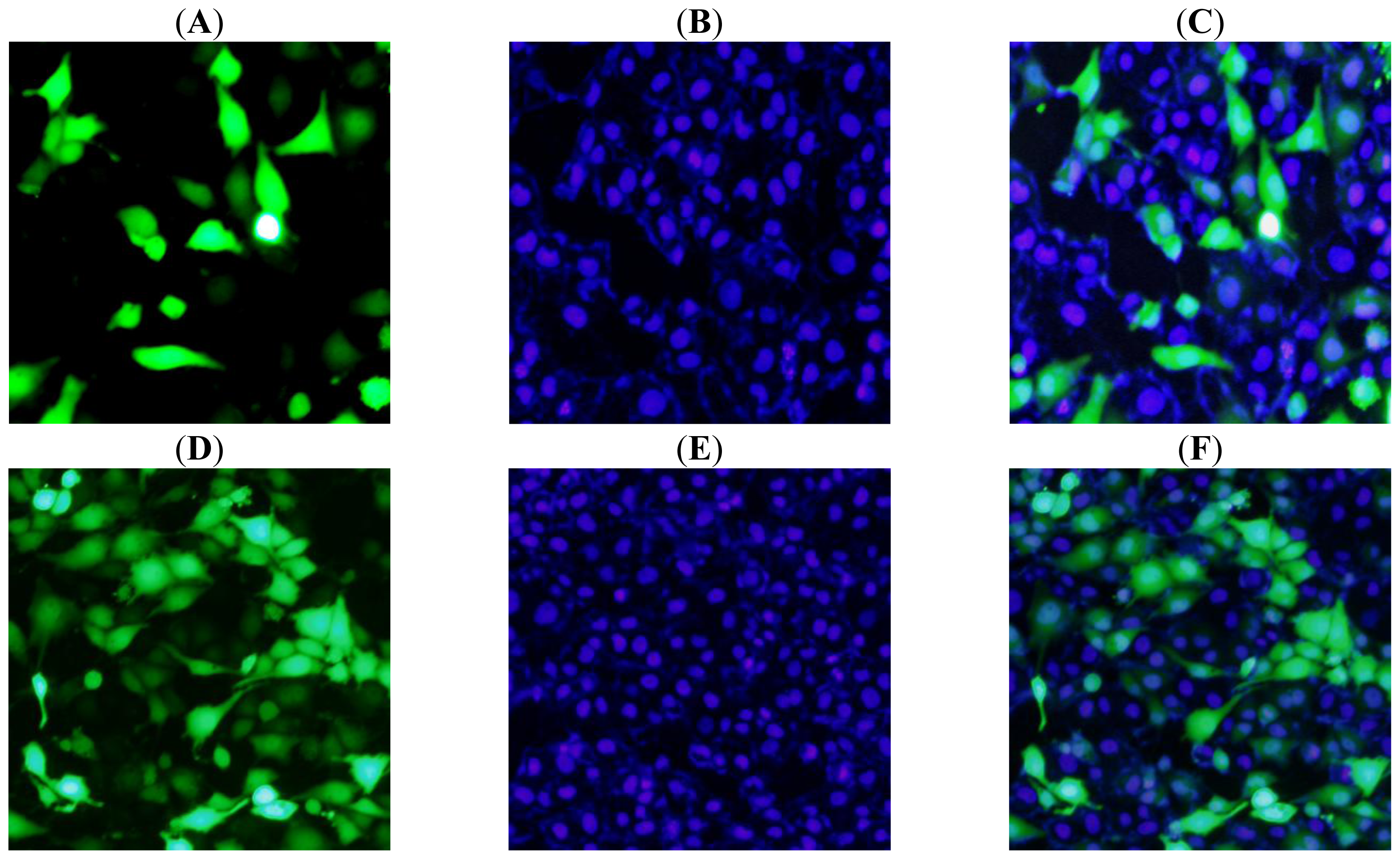
2.5. Subcellular Localization of Tob1 in PK15 Cells
3. Experimental Section
3.1. Source of Animals and Tissues
3.2. RNA Extraction and cDNA Preparation
3.3. Cloning, Sequencing and Analysis of Tob1 mRNA Sequences
3.4. Isolation of 3′ cDNA by GLGI
3.5. TaqMan Real-Time PCR Analysis
3.6. Real-Time PCR Analysis During C2C12 Differentiation Time
3.7. Chromosome Mapping Using IMpRH
3.8. Construction of the Subcellular Localization Vector
3.9. Cell Culture and Transient Transfection
4. Conclusions
Acknowledgements
References
- Matsuda, S; Rouaul, J; Magaud, J; Berthet, C. In search of a function for the TIS21/PC3/BTG1/TOB family. FEBS Lett 2001, 49, 67–72. [Google Scholar]
- Miller, CW; Koeffle, HP. Cyclin-dependent kinase inhibitors in human neoplasms. Leukemia 1997, 11, 370–371. [Google Scholar]
- Sasajim, H; Nakagawa, K; Yokosawa, H. Anti-proliferative proteins of the BTG/Tob family are degraded by the ubiquitin-proteasome system. Eur. J. Biochem 2002, 269, 3596–3604. [Google Scholar]
- Feng, Z; Tang, ZL; Li, K; Liu, B; Yu, M; Zhao, SH. Molecular characterization of the BTG2 and BTG3 genes in fetal muscle development of pigs. Gene 2007, 403, 170–177. [Google Scholar]
- Matsuda, S; Kawamura, TJ; Ohsugi, M; Yoshida, M; Emi, M; Nakamura, Y; Onda, M; Yoshida, Y; Nishiya, A; Yamamoto, T. Tob, a novel protein that interacts with p185erbB2, is associated with anti-proliferative activity. Oncogene 1996, 12, 705–713. [Google Scholar]
- Suzuk, T; Tsuzuku, JK; Ajim, R; Nakamura, T; Yoshida, Y; Yamamoto, T. Phosphorylation of three regulatory serines of Tob by Erk1 and Erk2 is required for Ras-mediated cell proliferation and transformation. Gene. Develop 2002, 16, 1356–1370. [Google Scholar]
- Tzachanis, D; Freema, GJ; Hirano, N; Van Puijenbroek, AA; Delfs, MW; Berezovskaya, A; Nadler, LM; Boussiotis, VA. Tob is a negative regulator of activation that is expressed in anergic and quiescent T cells. Nat. Immunol 2001, 2, 1174–1182. [Google Scholar]
- Maekawa, M; Nishida, E; Tanoue, T. Identification of the antiproliferative protein Tob as a MAPK substrate. J. Biol. Chem 2002, 277, 37783–37788. [Google Scholar]
- Jin, M; Wang, XM; Tu, Y; Zhang, XH; Gao, X; Guo, N; Xie, Z; Zhao, G; Jing, N; Li, BM; Yu, L. The negative cell cycle regulator, Tob (transducer of ErbB2), is a multifunctional protein involved in hippocampus-dependent learning and memory. Neuroscience 2005, 131, 647–659. [Google Scholar]
- Yoshida, Y; Tanaka, S; Umemori, H; Minowa, O; Usui, M; Ikematsu, N; Hosoda, E; Imamura, T; Kuno, J; Yamashita, T; Miyazono, K; Noda, M; Noda, T; Yamamoto, T. Negative regulation of BMP/Smad signaling by Tob in osteoblasts. Cell 2000, 103, 1085–1097. [Google Scholar]
- Jia, SJ; Meng, AM. Tob genes in development and homeostasis. Develop. Dynam 2007, 236, 913–921. [Google Scholar]
- Yoshida, Y; Nakamura, T; Komoda, M; Satoh, H; Suzuki, T; Tsuzuku, JK; Miyasaka, T; Yoshida, EH; Umemori, H; Kunisaki, RK; Tani, K; Ishii, S; Mori, S; Suganuma, M; Noda, T; Yamamoto, T. Mice lacking a transcriptional corepressor Tob are predisposed to cancer. Gene. Dev 2003, 17, 1201–1206. [Google Scholar]
- Iwanaga, K; Sueoka, N; Sato, A; Sakuragi, T; Sakao, Y; Tominaga, M; Suzuki, T; Yoshida, Y; K-Tsuzuku, J; Yamamoto, T; Hayashi, S; Nagasawa, K; Sueoka, E. Alteration of expression or phosphorylation status of Tob, a novel tumor suppressor gene product, is an early event in lung cancer. Cancer Lett 2003, 202, 71–79. [Google Scholar]
- Shi, KH; Zhang, LX; Meng, AM. Cloning and expression analysis of zebrafish Tob1 gene. Dev. Genes Evol 2004, 214, 309–311. [Google Scholar]
- Xiong, B; Rui, Y; Zhang, M; Shi, K; Jia, S; Tian, T; Yin, K; Huang, H; Lin, S; Zhao, X; Chen, Y; Chen, YG; Lin, SC; Meng, AM. Tob1 controls dorsal development of zebrafish embryos by antagonizing maternal betacatenin transcriptional activity. Dev Cell 2006, 11, 225–238. [Google Scholar]
- Tang, ZL; Li, Y; Wan, P; Li, XP; Zhao, SH; Liu, B; Fan, B; Zhu, MJ; Yu, M; Li, K. LongSAGE analysis of skeletal muscle at three prenatal stages in Tongcheng and Landrace pigs. Genome Biol 2007, 8, R115. [Google Scholar]
- Yoshida, Y; Matsud, S; Yamamoto, T. Cloning and characterization of the mouse Tob gene. Gene 1997, 191, 109–113. [Google Scholar]
- Kent, WJ; Sugnet, CW; Furey, TS; Roskin, KM; Pringle, TH; Zahler, AM; Haussler, D. The human genome browser at UCSC. Genome Res 2002, 12, 996–1006. [Google Scholar]
- Kawamura-Tsuzuku, J; Suzuki, T; Yoshida, Y; Yamamoto, T. Nuclear localization of Tob is important for regulation of its antiproliferative activity. Oncogene 2004, 23, 6630–6638. [Google Scholar]
- Maekawa, M; Yamamoto, T; Nishida, E. Regulation of subcellular localization of the antiproliferative protein Tob by its nuclear export signal and bipartite nuclear localization signal sequences. Exp. Cell Res 2004, 295, 59–65. [Google Scholar]
- Boeckmann, B; Bairoch, A; Apweiler, R; Blatter, MC; Estreicher, A; Gasteiger, E; Martin, MJ; Michoud, K; O’Donovan, C; Phan, I; Pilbout, S; Schneider, M. The SWISS-PROT protein knowledgebase and its supplement TrEMBL in 2003. Nucl. Acids Res 2003, 31, 365–370. [Google Scholar]
- Gasteiger, E; Gattiker, A; Hoogland, C; Ivanyi, I; Appel, RD; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucl. Acids Res 2003, 31, 3784–3788. [Google Scholar]
- Chen, J; Sun, M; Lee, S; Zhou, G; Rowley, JD; Wang, SM. Identifying novel transcripts and novel genes in the human genome by using novel SAGE tags. Proc. Nat. Acad. Sci. USA 2002, 99, 12257–12262. [Google Scholar]
- Tang, ZL; Li, Y; Zhao, SH; Liu, B; Fan, B; Li, K. A modified GLGI method for identification of novel porcine genes from long serial analysis of gene expression tags. Chin. J. Agr. Biotechnol 2007, 4, 111–115. [Google Scholar]
- Livak, KJ; Schmittgen, TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar]
- Yerle, M; Pinton, P; Robic, A; Alfonso, A; Palvadeau, Y; Delcros, C; Hawken, R; Alexander, L; Beattie, C; Schook, L; Milan, D; Gellin, J. Construction of a whole-genome radiation hybrid panel for high-resolution gene mapping in pigs. Cytogenet. Cell Genet 1998, 82, 182–188. [Google Scholar]
- Milan, D; Hawken, R; Cabau, C; Leroux, S; Genet, C; Lahbib, Y; Tosser, G; Robic, A; Hatey, F; Alexander, L; Beattie, C; Schook, L; Yerle, M; Gellin, J. IMpRH server: An RH mapping server available on the web. Bioinformatics 2000, 16, 558–559. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T33 | T65 | T90 | L33 | L65 | L90 | |
|---|---|---|---|---|---|---|
| T33 | 1 (1) | 0.5058 (3) | 0.0045** (11) | 1 (1) | 0.3232 (3) | 0.4929 (2) |
| T65 | 1 | 0.0287* | 0.3162 | 0.6567 | 0.5321 | |
| T90 | 1 | 0.0034** | 0.0273* | 0.0129* | ||
| L33 | 1 | 0.3384 | 0.4616 | |||
| L65 | 1 | 0.5546 | ||||
| L90 | 1 |
| Gene | Retention fraction% | Chromosome | Linked marker | Breakage frequency | RH distance(Ray) | LOD score |
|---|---|---|---|---|---|---|
| Tob1 | 27 | 12 | SS04H11 | 0.5 | 0.69 | 5.52 |
| Primer | Sequences (5′–3′) | Size (bp) | Tm (°C) |
|---|---|---|---|
| Tob1F | AAGCAGCCCGAACAAGAC | 1462 | 55.6 |
| Tob1R | AATCAGCCATGTCCTTGC | ||
| Tob1-MF | GACCCCGTCCTCGCCAAC | 170 | 64 |
| Tob1-MR | TGTTCGGGCTGCTTCCACC | ||
| GLGIF | CATGCAGTATTCTAACCAGCA | 60 | 54.4 |
| GLGIR | ACTATCTAGAGCGGCCGCTT | ||
| Exp-F | TGATCGAGCAGGCATCCAA | 116 | 58 |
| Exp-R | TTCGCCGATCTGGTAGGAAAC | ||
| GAPDH-F | GGTGAAGGTCGGAGTGAACG | 233 | 60 |
| GAPDH-R | CTCGCTCCTGGAAGATGGTG | ||
| Loc-F | AGCGCTATGCAGCTTGAAATCCAAGTAG | 1038 | 61 |
| Loc-R | AGATCTGTTAGCCATAACAGGCTGGAAT |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yuan, J.; Cao, J.-Y.; Tang, Z.-L.; Wang, N.; Li, K. Molecular Characterization of Tob1 in Muscle Development in Pigs. Int. J. Mol. Sci. 2011, 12, 4315-4326. https://doi.org/10.3390/ijms12074315
Yuan J, Cao J-Y, Tang Z-L, Wang N, Li K. Molecular Characterization of Tob1 in Muscle Development in Pigs. International Journal of Molecular Sciences. 2011; 12(7):4315-4326. https://doi.org/10.3390/ijms12074315
Chicago/Turabian StyleYuan, Jing, Ji-Yue Cao, Zhong-Lin Tang, Ning Wang, and Kui Li. 2011. "Molecular Characterization of Tob1 in Muscle Development in Pigs" International Journal of Molecular Sciences 12, no. 7: 4315-4326. https://doi.org/10.3390/ijms12074315