The Important Molecular Markers on Chromosome 17 and Their Clinical Impact in Breast Cancer

Abstract
:
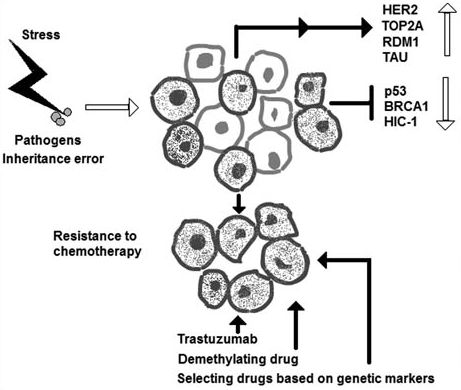
1. Introduction
2. HER2
3. TOP2A
4. TAU
5. RDM1
6. BRCA1
7. P53
8. HIC-1
9. Conclusion
Acknowledgments
- Conflict of InterestThere is no potential or actual personal, financial or political interest related to this article.
References
- Reinholz, MM; Bruzek, AK; Visscher, DW; Lingle, WL; Schroeder, MJ; Perez, EA; Jenkins, RB. Breast cancer and aneusomy 17: Implications for carcinogenesis and therapeutic response. Lancet Oncol 2009, 10, 267–277. [Google Scholar]
- Ross, JS; McKenna, BJ. The HER-2/neu oncogene in tumors of the gastrointestinal tract. Cancer Invest 2001, 19, 554–568. [Google Scholar]
- Brien, TP; Depowski, PL; Sheehan, CE; Ross, JS; McKenna, BJ. Prognostic factors in gastric cancer. Modern Pathol 1998, 11, 870–877. [Google Scholar]
- Sharif, S; Ramanathan, RK; Potter, D; Cieply, K; Krasinskas, AM. HER2 gene amplification and chromosome 17 copy number do not predict survival of patients with resected pancreatic adenocarcinoma. Digest Dis Sci 2008, 53, 3026–3032. [Google Scholar]
- Shafizadeh, N; Grenert, JP; Sahai, V; Kakar, S. Epidermal growth factor receptor and HER-2/neu status by immunohistochemistry and fluorescence in situ hybridization in adenocarcinomas of the biliary tree and gallbladder. Hum Pathol 2010, 41, 485–492. [Google Scholar]
- Rosenthal, SI; Depowski, PL; Sheehan, CE; Ross, JS. Comparison of HER-2/neu oncogene amplification detected by fluorescence in situ hybridization in lobular and ductal breast cancer. Appl Immunohistochem Mol Morphol 2002, 10, 40–46. [Google Scholar]
- Bedard, PL; Piccart-Gebhart, MJ. Current paradigms for the use of HER2-targeted therapy in early-stage breast cancer. Clin Breast Cancer 2008, 8, S157–S165. [Google Scholar]
- Yin, W; Jiang, Y; Shen, Z; Shao, Z; Lu, J. Trastuzumab in the adjuvant treatment of her2-positive early breast cancer patients: A meta-analysis of published randomized controlled trials. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Wolff, AC; Hammond, ME; Schwartz, JN; Hagerty, KL; Allred, DC; Cote, RJ; Dowsett, M; Fitzgibbons, PL; Hanna, WM; Langer, A; et al. American society of clinical oncology/college of american pathologists guideline recommendations for human epidermal growth factor receptor 2 testing in breast cancer. Arch Pathol Lab Med 2007, 131, 18–43. [Google Scholar]
- Chia, S; Norris, B; Speers, C; Cheang, M; Gilks, B; Gown, AM; Huntsman, D; Olivotto, IA; Nielsen, TO; Gelmon, K. Human epidermal growth factor receptor 2 overexpression as a prognostic factor in a large tissue microarray series of node-negative breast cancers. J Clin Oncol 2008, 26, 5697–5704. [Google Scholar]
- Guiu, S; Liegard, M; Favier, L; van Praagh, I; Largillier, R; Weber, B; Coeffic, D; Moreau, L; Priou, F; Campone, M; et al. Long-term follow-up of her2-overexpressing stage ii or iii breast cancer treated by anthracycline-free neoadjuvant chemotherapy. Ann Oncol 2011, 22, 321–328. [Google Scholar]
- Wolff, AC; Hammond, ME; Schwartz, JN; Hagerty, KL; Allred, DC; Cote, RJ; Dowsett, M; Fitzgibbons, PL; Hanna, WM; Langer, A; et al. American society of clinical oncology/college of american pathologists guideline recommendations for human epidermal growth factor receptor 2 testing in breast cancer. J Clin Oncol 2007, 25, 118–145. [Google Scholar]
- Carlson, RW; Moench, SJ; Hammond, ME; Perez, EA; Burstein, HJ; Allred, DC; Vogel, CL; Goldstein, LJ; Somlo, G; Gradishar, WJ; et al. Her2 testing in breast cancer: Nccn task force report and recommendations. J Natl Compr Cancer Netw 2006, 4, S1–S22, quiz S23–S24. [Google Scholar]
- Bouchalova, K; Trojanec, R; Kolar, Z; Cwiertka, K; Cernakova, I; Mihal, V; Hajduch, M. Analysis of erbb2 and top2a gene status using fluorescence in situ hybridization versus immunohistochemistry in localized breast cancer. Neoplasma 2006, 53, 393–401. [Google Scholar]
- Salido, M; Tusquets, I; Corominas, JM; Suarez, M; Espinet, B; Corzo, C; Bellet, M; Fabregat, X; Serrano, S; Sole, F. Polysomy of chromosome 17 in breast cancer tumors showing an overexpression of erbb2: A study of 175 cases using fluorescence in situ hybridization and immunohistochemistry. Breast Cancer Res 2005, 7, R267–R273. [Google Scholar]
- Zhu, X; Lu, Y; Lu, H; Yang, W; Tu, X; Cai, X; Zhou, X. Genetic alterations and protein expression of her2 and chromosome 17 polysomy in breast cancer. Hum Pathol 2011. [Google Scholar] [CrossRef]
- Moelans, CB; de Weger, RA; van Diest, PJ. Absence of chromosome 17 polysomy in breast cancer: Analysis by cep17 chromogenic in situ hybridization and multiplex ligation-dependent probe amplification. Breast Cancer Res Treat 2010, 120, 1–7. [Google Scholar]
- Shah, SS; Wang, Y; Tull, J; Zhang, S. Effect of high copy number of her2 associated with polysomy 17 on her2 protein expression in invasive breast carcinoma. Diagn Mol Pathol 2009, 18, 30–33. [Google Scholar]
- Downey, L; Livingston, RB; Koehler, M; Arbushites, M; Williams, L; Santiago, A; Guzman, R; Villalobos, I; Di Leo, A; Press, MF. Chromosome 17 polysomy without human epidermal growth factor receptor 2 amplification does not predict response to lapatinib plus paclitaxel compared with paclitaxel in metastatic breast cancer. Clin Cancer Res 2010, 16, 1281–1288. [Google Scholar]
- Slamon, DJ; Press, MF. Alterations in the top2a and her2 genes: Association with adjuvant anthracycline sensitivity in human breast cancers. J Natl Cancer Inst 2009, 101, 615–618. [Google Scholar]
- O’Malley, FP; Chia, S; Tu, D; Shepherd, LE; Levine, MN; Bramwell, VH; Andrulis, IL; Pritchard, KI. Topoisomerase ii alpha and responsiveness of breast cancer to adjuvant chemotherapy. J Natl Cancer Inst 2009, 101, 644–650. [Google Scholar]
- Orlando, L; del Curto, B; Gandini, S; Ghisini, R; Pietri, E; Torrisi, R; Balduzzi, A; Cardillo, A; Dellapasqua, S; Veronesi, P; et al. Topoisomerase IIalpha gene status and prediction of pathological complete remission after anthracycline-based neoadjuvant chemotherapy in endocrine non-responsive Her2/neu-positive breast cancer. Breast 2008, 17, 506–511. [Google Scholar]
- Miyoshi, Y; Kurosumi, M; Kurebayashi, J; Matsuura, N; Takahashi, M; Tokunaga, E; Egawa, C; Masuda, N; Kono, S; Morimoto, K; et al. Predictive factors for anthracycline-based chemotherapy for human breast cancer. Breast Cancer 2010, 17, 103–109. [Google Scholar]
- McGrogan, BT; Gilmartin, B; Carney, DN; McCann, A. Taxanes, microtubules and chemoresistant breast cancer. Biochim Biophys Acta 2008, 1785, 96–132. [Google Scholar]
- Pusztai, L. Markers predicting clinical benefit in breast cancer from microtubule-targeting agents. Ann Oncol 2007, 18, xii15–xii20. [Google Scholar]
- Andre, F; Hatzis, C; Anderson, K; Sotiriou, C; Mazouni, C; Mejia, J; Wang, B; Hortobagyi, GN; Symmans, WF; Pusztai, L. Microtubule-associated protein-tau is a bifunctional predictor of endocrine sensitivity and chemotherapy resistance in estrogen receptor-positive breast cancer. Clin Cancer Res 2007, 13, 2061–2067. [Google Scholar]
- Tanaka, S; Nohara, T; Iwamoto, M; Sumiyoshi, K; Kimura, K; Takahashi, Y; Tanigawa, N. Tau expression and efficacy of paclitaxel treatment in metastatic breast cancer. Cancer Chemother Pharmacol 2009, 64, 341–346. [Google Scholar]
- Milne, GT; Ho, T; Weaver, DT. Modulation of saccharomyces cerevisiae DNA double-strand break repair by srs2 and rad51. Genetics 1995, 139, 1189–1199. [Google Scholar]
- Hamimes, S; Arakawa, H; Stasiak, AZ; Kierzek, AM; Hirano, S; Yang, YG; Takata, M; Stasiak, A; Buerstedde, JM; van Dyck, E. RDM1, a novel RNA recognition motif (RRM)-containing protein involved in the cell response to cisplatin in vertebrates. J Biol Chem 2005, 280, 9225–9235. [Google Scholar]
- Hamimes, S; Bourgeon, D; Stasiak, AZ; Stasiak, A; Van Dyck, E. Nucleic acid-binding properties of the RRM-containing protein RDM1. Biochem Biophys Res Commun 2006, 344, 87–94. [Google Scholar]
- Messaoudi, L; Yang, YG; Kinomura, A; Stavreva, DA; Yan, G; Bortolin-Cavaille, ML; Arakawa, H; Buerstedde, JM; Hainaut, P; Cavaille, J; et al. Subcellular distribution of human rdm1 protein isoforms and their nucleolar accumulation in response to heat shock and proteotoxic stress. Nucleic Acids Res 2007, 35, 6571–6587. [Google Scholar]
- Abe, H; Wada, M; Kohno, K; Kuwano, M. Altered drug sensitivities to anticancer agents in radiation-sensitive DNA repair deficient yeast mutants. Anticancer Res 1994, 14, 1807–1810. [Google Scholar]
- Yamaguchi-Iwai, Y; Sonoda, E; Buerstedde, JM; Bezzubova, O; Morrison, C; Takata, M; Shinohara, A; Takeda, S. Homologous recombination, but not DNA repair, is reduced in vertebrate cells deficient in rad52. Mol Cell Biol 1998, 18, 6430–6435. [Google Scholar]
- van Hille, B; Clerc, X; Creighton, AM; Hill, BT. Differential expression of topoisomerase i and rad52 protein in yeast reveals new facets of the mechanism of action of bisdioxopiperazine compounds. Br J Cancer 1999, 81, 800–807. [Google Scholar]
- Miki, Y; Swensen, J; Shattuck-Eidens, D; Futreal, PA; Harshman, K; Tavtigian, S; Liu, Q; Cochran, C; Bennett, LM; Ding, W; et al. A strong candidate for the breast and ovarian cancer susceptibility gene brca1. Science 1994, 266, 66–71. [Google Scholar]
- Rhiem, K; Todt, U; Wappenschmidt, B; Klein, A; Wardelmann, E; Schmutzler, RK. Sporadic breast carcinomas with somatic brca1 gene deletions share genotype/phenotype features with familial breast carcinomas. Anticancer Res 2010, 30, 3445–3449. [Google Scholar]
- Tassone, P; di Martino, MT; Ventura, M; Pietragalla, A; Cucinotto, I; Calimeri, T; Bulotta, A; Neri, P; Caraglia, M; Tagliaferri, P. Loss of brca1 function increases the antitumor activity of cisplatin against human breast cancer xenografts in vivo. Cancer Biol Ther 2009, 8, 648–653. [Google Scholar]
- Liu, X; Holstege, H; van der Gulden, H; Treur-Mulder, M; Zevenhoven, J; Velds, A; Kerkhoven, RM; van Vliet, MH; Wessels, LF; Peterse, JL; et al. Somatic loss of brca1 and p53 in mice induces mammary tumors with features of human brca1-mutated basal-like breast cancer. Proc Natl Acad Sci USA 2007, 104, 12111–12116. [Google Scholar]
- Birgisdottir, V; Stefansson, OA; Bodvarsdottir, SK; Hilmarsdottir, H; Jonasson, JG; Eyfjord, JE. Epigenetic silencing and deletion of the brca1 gene in sporadic breast cancer. Breast Cancer Res 2006, 8, R38. [Google Scholar]
- Wei, M; Grushko, TA; Dignam, J; Hagos, F; Nanda, R; Sveen, L; Xu, J; Fackenthal, J; Tretiakova, M; Das, S; Olopade, OI. BRCA1 promoter methylation in sporadic breast cancer is associated with reduced BRCA1 copy number and chromosome 17 aneusomy. Cancer Res 2005, 65, 10692–10699. [Google Scholar]
- Lo, YL; Yu, JC; Huang, CS; Tseng, SL; Chang, TM; Chang, KJ; Wu, CW; Shen, CY. Allelic loss of the BRCA1 and BRCA2 genes and other regions on 17q and 13q in breast cancer among women from Taiwan (area of low incidence but early onset). Int J Cancer 1998, 79, 580–587. [Google Scholar]
- Soussi, T. The p53 tumor suppressor gene: From molecular biology to clinical investigation. Ann NY Acad Sci 2000, 910, 121–139. [Google Scholar]
- Rigatti, MJ; Verma, R; Belinsky, GS; Rosenberg, DW; Giardina, C. Pharmacological inhibition of mdm2 triggers growth arrest and promotes DNA breakage in mouse colon tumors and human colon cancer cells. Mol Carcinog 2011. [Google Scholar] [CrossRef]
- Tovar, C; Rosinski, J; Filipovic, Z; Higgins, B; Kolinsky, K; Hilton, H; Zhao, X; Vu, BT; Qing, W; Packman, K; et al. Small-molecule mdm2 antagonists reveal aberrant p53 signaling in cancer: Implications for therapy. Proc Natl Acad Sci USA 2006, 103, 1888–1893. [Google Scholar]
- Guerardel, C; Deltour, S; Pinte, S; Monte, D; Begue, A; Godwin, AK; Leprince, D. Identification in the human candidate tumor suppressor gene HIC-1 of a new major alternative TATA-less promoter positively regulated by p53. J Biol Chem 2001, 276, 3078–3089. [Google Scholar]
- Nishida, N; Nagasaka, T; Nishimura, T; Ikai, I; Boland, CR; Goel, A. Aberrant methylation of multiple tumor suppressor genes in aging liver, chronic hepatitis, and hepatocellular carcinoma. Hepatology 2008, 47, 908–918. [Google Scholar]
- Oh, BK; Kim, H; Park, HJ; Shim, YH; Choi, J; Park, C; Park, YN. DNA methyltransferase expression and DNA methylation in human hepatocellular carcinoma and their clinicopathological correlation. Int J Mol Med 2007, 20, 65–73. [Google Scholar]
- Park, HJ; Yu, E; Shim, YH. DNA methyltransferase expression and DNA hypermethylation in human hepatocellular carcinoma. Cancer Lett 2006, 233, 271–278. [Google Scholar]
- Skvortsova, TE; Rykova, EY; Tamkovich, SN; Bryzgunova, OE; Starikov, AV; Kuznetsova, NP; Vlassov, VV; Laktionov, PP. Cell-free and cell-bound circulating DNA in breast tumours: DNA quantification and analysis of tumour-related gene methylation. Br J Cancer 2006, 94, 1492–1495. [Google Scholar]
- Dong, SM; Lee, EJ; Jeon, ES; Park, CK; Kim, KM. Progressive methylation during the serrated neoplasia pathway of the colorectum. Mod Pathol 2005, 18, 170–178. [Google Scholar]
- Ahuja, N; Li, Q; Mohan, AL; Baylin, SB; Issa, JP. Aging and DNA methylation in colorectal mucosa and cancer. Cancer Res 1998, 58, 5489–5494. [Google Scholar]
- Kanai, Y; Hui, AM; Sun, L; Ushijima, S; Sakamoto, M; Tsuda, H; Hirohashi, S. DNA hypermethylation at the D17S5 locus and reduced HIC-1 mRNA expression are associated with hepatocarcinogenesis. Hepatology 1999, 29, 703–709. [Google Scholar]
- Kanai, Y; Ushijima, S; Ochiai, A; Eguchi, K; Hui, A; Hirohashi, S. DNA hypermethylation at the D17S5 locus is associated with gastric carcinogenesis. Cancer Lett 1998, 122, 135–141. [Google Scholar]
- Nosho, K; Irahara, N; Shima, K; Kure, S; Kirkner, GJ; Schernhammer, ES; Hazra, A; Hunter, DJ; Quackenbush, J; Spiegelman, D; et al. Comprehensive biostatistical analysis of CpG island methylator phenotype in colorectal cancer using a large population-based sample. PLoS One 2008, 3. [Google Scholar] [CrossRef]
- Tsuji, N; Furuse, K; Asanuma, K; Furuya, M; Kondoh, K; Kamagata, C; Sasaki, M; Kobayashi, D; Yagihashi, A; Takahashi, H; Watanabe, N. Mutations of the p53 gene and loss of heterozygosity at chromosome 17p13.1 are associated with increased survivin expression in breast cancer. Breast Cancer Res Treat 2004, 87, 23–31. [Google Scholar]
- Wales, MM; Biel, MA; el Deiry, W; Nelkin, BD; Issa, JP; Cavenee, WK; Kuerbitz, SJ; Baylin, SB. p53 activates expression of HIC-1, a new candidate tumour suppressor gene on 17p13.3. Nat Med 1995, 1, 570–577. [Google Scholar]
- Zhang, B; Chambers, KJ; Leprince, D; Faller, DV; Wang, S. Requirement for chromatin-remodeling complex in novel tumor suppressor HIC1-mediated transcriptional repression and growth control. Oncogene 2009, 28, 651–661. [Google Scholar]
- Zhang, W; Zeng, X; Briggs, KJ; Beaty, R; Simons, B; Chiu Yen, RW; Tyler, MA; Tsai, HC; Ye, Y; Gesell, GS; et al. A potential tumor suppressor role for Hic1 in breast cancer through transcriptional repression of ephrin-A1. Oncogene 2010, 29, 2467–2476. [Google Scholar]
- Zhang, B; Faller, DV; Wang, S. Hic1 regulates tumor cell responses to endocrine therapies. Mol Endocrinol 2009, 23, 2075–2085. [Google Scholar]
- Nicoll, G; Crichton, DN; McDowell, HE; Kernohan, N; Hupp, TR; Thompson, AM. Expression of the Hypermethylated in Cancer gene (HIC-1) is associated with good outcome in human breast cancer. Br J Cancer 2001, 85, 1878–1882. [Google Scholar]


{kind=link}
{kind=link}
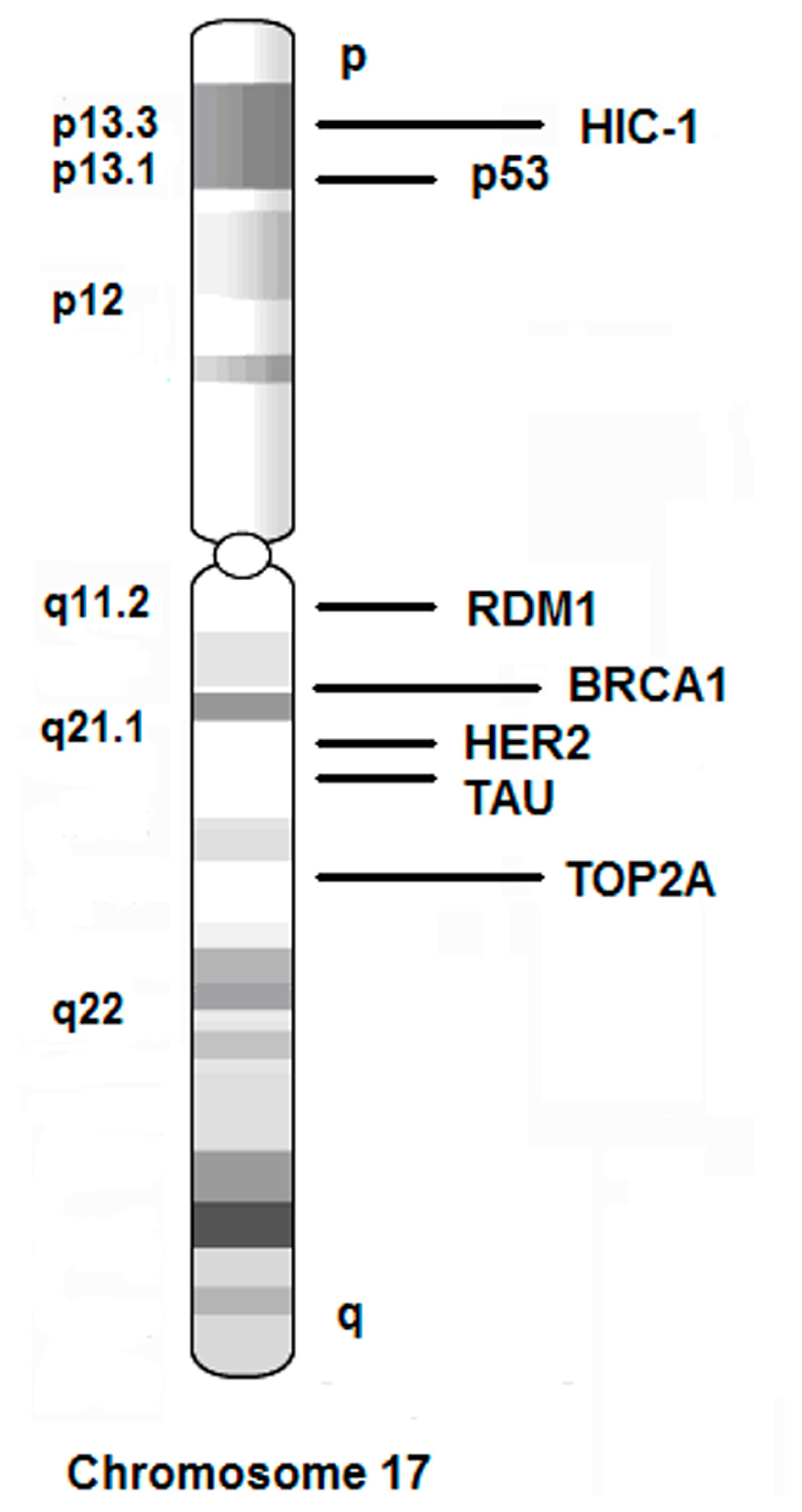
| Gene ID | Name | Location | Functions |
|---|---|---|---|
| 2064 | ERBB2/HER2 | 17q21.1 | Epidermal growth factor (EGF) receptor family of receptor tyrosine kinases. Amplification and/or overexpression have been reported in numerous cancers. |
| 7153 | TOP2A | 17q21-q22 | DNA topoisomerase, controls and alters the topologic states of DNA during transcription. It is associated with the development of drug resistance. |
| 201299 | RDM1 | 17q11.2 | RAD52 protein encoded by RDM1 is involved in DNA double-strand break repair and recombination event. Disruption of the RDM1 gene resulted in an increased sensitivity to the anti-cancer drug cisplatin. |
| 7157 | P53 | 17p13.1 | P53 responds to diverse cellular stresses to regulate target genes that induce cell cycle arrest, apoptosis, senescence, DNA repair. It is accumulated in a variety of transformed cells. |
| 672 | BRCA1 | 17q21 | BRCA1 plays a role in maintaining genomic stability. It acts as a tumor suppressor. BRCA1 combines with other tumor suppressors, to form a BRCA1-associated genome surveillance complex (BASC). Mutations in this gene are responsible for approximately 40% of inherited breast cancers and more than 80% of inherited breast and ovarian cancers. |
| 3090 | HIC-1 | 17p13.3 | Hypermethylated in cancer 1, a candidate tumor suppressor gene which undergoes allelic loss in breast and other human cancers. The human HIC-1 gene is a target gene of p53. |
| 4137 | TAU | 17q21.1 | Microtubule-associated protein TAU (MAPT), functions to keep cell shape, microvesicle transportation and spindle formation. Interfering spindle microtubule dynamics will cause cell cycle arrest and apoptosis. TAU detection helps to identify those patients who are most likely to benefit from taxane treatment and resistant to paclitaxel treatment. |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, W.; Yu, Y. The Important Molecular Markers on Chromosome 17 and Their Clinical Impact in Breast Cancer. Int. J. Mol. Sci. 2011, 12, 5672-5683. https://doi.org/10.3390/ijms12095672
Zhang W, Yu Y. The Important Molecular Markers on Chromosome 17 and Their Clinical Impact in Breast Cancer. International Journal of Molecular Sciences. 2011; 12(9):5672-5683. https://doi.org/10.3390/ijms12095672
Chicago/Turabian StyleZhang, Wei, and Yingyan Yu. 2011. "The Important Molecular Markers on Chromosome 17 and Their Clinical Impact in Breast Cancer" International Journal of Molecular Sciences 12, no. 9: 5672-5683. https://doi.org/10.3390/ijms12095672
APA StyleZhang, W., & Yu, Y. (2011). The Important Molecular Markers on Chromosome 17 and Their Clinical Impact in Breast Cancer. International Journal of Molecular Sciences, 12(9), 5672-5683. https://doi.org/10.3390/ijms12095672