1. Introduction
Ovarian carcinoma (OC) is the most lethal gynecologic malignancy. The majority of patients are diagnosed with advanced-stage disease and despite aggressive surgery and adjuvant platinum-based combination therapy, succumb to their disease, primarily due to chemotherapy resistance [
1,
2]. Despite the obvious role of FIGO stage, tumor grade, residual disease (RD) volume after surgery and the presence of ascites in providing prognostic information in OC, these factors fail to predict clinical outcome for individual patients owing to a significant degree of tumor heterogeneity. Widely accepted biological factors that are able to predict chemoresponse and survival in OC are unavailable to date, a fact that in part reflects our limited understanding of metastatic OC, the cause of death for most patients.
The extracellular matrix (ECM) is a critical environmental determinant of tumor cell behavior. The matrix serves as a scaffold for the tumor cells to adhere, migrate and proliferate in response to the reservoir of growth factors and cytokines stored in it. Hyaluronan (also termed hyaluronic acid or hyaluronate; HA) is a prominent component of the ECM in many tissues, particularly in rapidly remodeling ones. Hyaluronan is a very large, linear, negatively-charged polysaccharide, which is composed of repeating non-sulphated disaccharides of glucuronate and
N-acetylglucosamine and exists as a high molecular weight polymer of approximately 10
4–10
7 Da. Its strong negative charge attracts a large associated volume of water capable of expansion up to 10,000 times its actual polymer volume. HA functions in normal physiology by maintaining tissue hydration and osmotic balance in tissues, and additionally regulates cell adhesion, migration, apoptosis and proliferation via interaction with specific cell surface receptors. In addition to its role in embryogenesis, there is increasing evidence for the involvement of HA in tumor progression of multiple cancers, through its effect on proliferation, apoptosis, angiogenesis, migration and invasion, as well as promotion of epithelial-to-mesenchymal transition (EMT) [
3,
4]. Multiple studies have documented the role of HA and its receptor CD44 in the binding of OC cells to mesothelial cells, a critical event in OC dissemination in the abdomen, and its role in tumor progression and in determining prognosis in this cancer [
5].
The biosynthesis of HA is regulated by three HA synthase isozymes, HAS1-3. There is a 50%–70% amino acid sequence identity between the different genes. However, they differ in their enzymatic activities. HAS1 and HAS2 synthesize HA molecules with high molecular weight, whereas HAS3 is produces low molecular weight molecules. HAS2 is the most commonly expressed family member in normal mammalian tissues, whereas HAS3 is the predominant enzyme in pathological conditions, such as inflammation and cancer [
6].
HA degradation is induced by a family of six hyaluronidases, consisting of Hyal1, Hyal2, Hyal3, Hyal4, PH-20/PSAM1 and the pseudogene HyalP1 [
7]. Of these, Hyal1 and Hyal2 are the most widely distributed and best characterized. The major transcript of Hyal3 does not possess enzymatic activity by itself and may merely have a supportive role in Hyal1 expression [
8].
Despite the wealth of literature regarding HA and CD44 expression in OC, little is known regarding the expression and clinical role of HA-synthesizing and HA-metabolizing enzymes in OC. Hyaluronidase activity was reported to be high in malignant ovarian tumors compared to their endometrial and cervical counterparts [
9]. However, hyaluronidase activity was lower in OC compared to benign and borderline tumors of the ovary in another study [
10]. In a study of HAS1-3 expression in 33 primary OC, HAS1 expression was reported to be associated with shorter overall survival (OS) [
11]. In the only study in which both HAS and Hyal members were investigated, analysis of 39 ovary specimens, including 20 serous OC, demonstrated expression of HAS2 and HAS3 mRNA, with low or undetectable HAS1, and HAS mRNA levels were not significantly related to HAS protein or hyaluronan accumulation. HYAL1 mRNA level was lower in grade 3 OC compared to normal ovaries and correlated with the enzymatic activity of tissue hyaluronidases, and inversely with HA staining intensity, with no such association for HYAL2 [
8].
Only one of the above studies analyzed metastatic OC specimens, and none included effusion specimens. Metastatic OC, particularly effusions, have different expression of cancer-associated molecules at the mRNA and protein level [
12]. OC of different histology is additionally increasingly regarded to represent different diseases with a unique biology and expression profile [
13,
14]. In view of these observations, the present study focused on the expression and clinical role of the various HAS and Hyal enzymes and their splice variants in serous OC effusions, and compared these expression pattern with solid primary and metastatic serous OC specimens.
3. Discussion
HA is widely expressed in human cancer and has been shown to have a central role in tumor biology [
3]. Recently, blocking of HA by small oligosaccharides [
16] and binding of paclitaxel to HA in order to improve tumor-specific delivery [
17–
19] have been investigated as therapeutic approaches in OC models
in vivo. This emphasizes the need to study HA and its regulators in primary and metastatic disease. HA levels in tissues are affected by the balance between the activities of HAS and Hyal enzymes. In the present study, we analyzed the dynamics in the expression of these enzymes at the various anatomic sites affected by OC. We limited our cohort to patients with serous OC, the majority of which, including all patients with effusions, were diagnosed with advanced-stage disease, in order to avoid confounding factors, and primarily the different expression patterns in OC of different histological type.
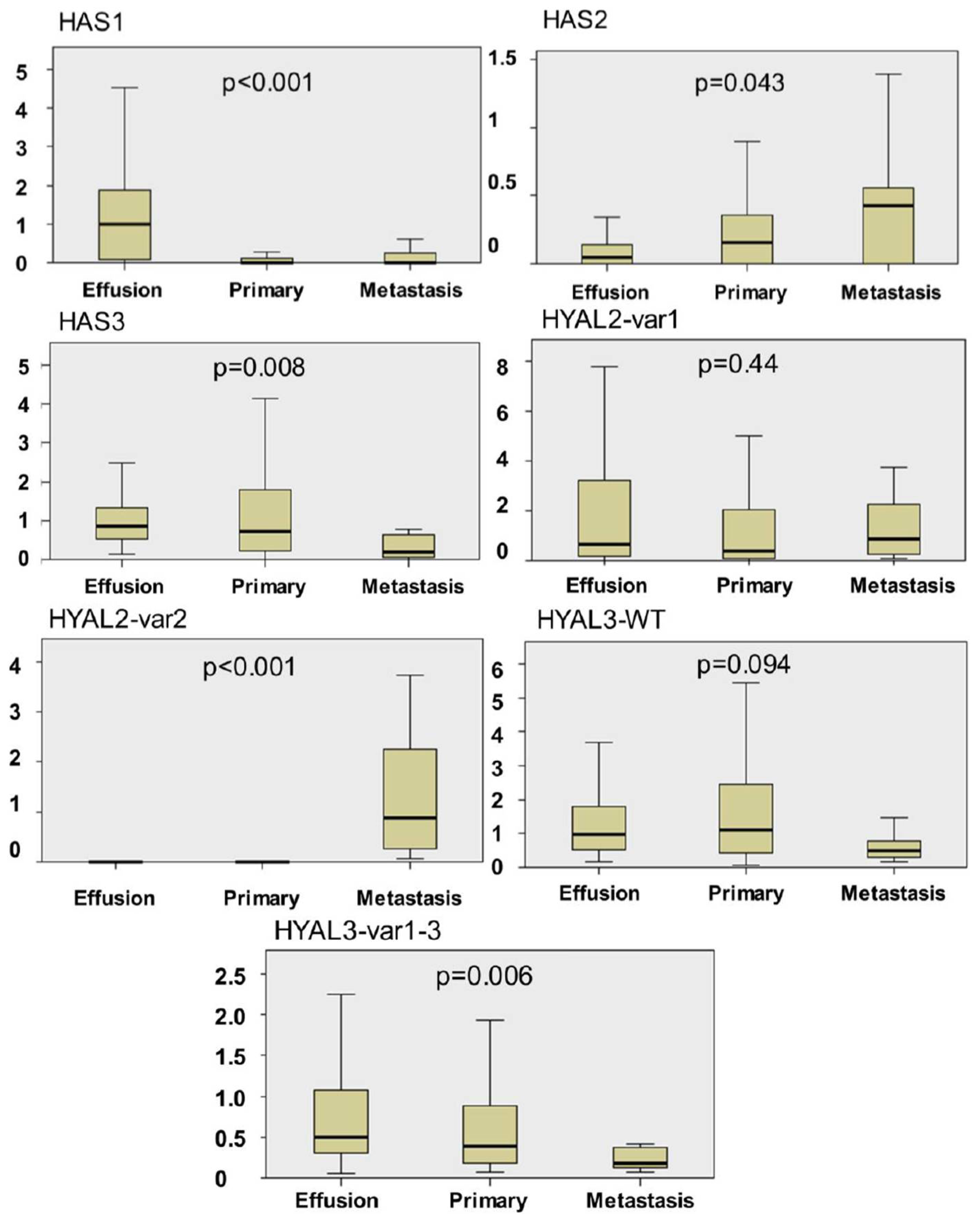
Hiltunen and co-workers previously showed higher HA levels in OC metastases compared to primary tumors, as well as benign and borderline tumors of the ovary, whereas hyaluronidase activity was lower in malignant compared to benign and borderline tumors [
10]. Higher levels of stromal HA were reported in another study by the same group [
20]. In our series, effusions had significantly higher HAS1 mRNA expression compared to primary OC and solid metastases, and were the only site were the alternative HAS1 variant was expressed, while the opposite was true for HAS2. HAS3 was more highly expressed in primary OC and effusions compared to solid metastases. The higher expression of HAS1 in effusions concurs with some extent with the observation of Nykopp
et al. [
8], who found very low levels of this enzyme in primary serous OC, although our series had more easily detectable expression of this enzyme also in primary OC.
Similarly to HAS members, Hyal family members were differently expressed when the 3 anatomic sites studied were compared. HYAL2-var2 mRNA was more highly expressed in solid metastases compared to effusions and primary carcinomas, whereas the opposite was true for HYAL3-var1-3. HYAL1 was uniformly absent. The latter finding is in agreement with the data Nykopp
et al., who observed reduced HYAL1 mRNA expression in OC compared to normal ovaries, benign tumors and borderline tumors using qRT-PCR [
8].
Our data suggest that HAS and Hyal family members, with the exception of Hyal1, are expressed at all anatomic sites in serous OC, but with changing expression levels. Although the majority of specimens were not patient-matched, this finding suggests a dynamic HAS synthesis along tumor progression in OC. The biological role of the splice variants we detected for HAS1, HYAL2 and HYAL3 in OC is yet to be studied.
Based on the differences in HAS and Hyal expression at the different anatomic sites, it seems to us that the more advanced disease is associated with a switch in the size of the synthesized HA to a shorter form and accompanied with an increase the catabolic power.
To the best of our knowledge, this study is the first to compare pre- and post-chemotherapy OC. The levels of HAS1 mRNA and its splice variant were significantly higher in pre-chemotherapy compared to post-chemotherapy effusions, whereas HYAL2-var1 and HYAL3-WT levels were significantly higher in post-chemotherapy effusions. The differences in HAS1 expression was significantly related to previous exposure to platinum, whereas both HAS1 and HYAL3-WT levels were related to exposure to paclitaxel. This suggests a change in the equilibrium between HA synthesis and catabolism following chemotherapy, which, in the absence of stromal cells in effusions, may reflect altered tumor phenotype through selection of chemoresistant tumor clones or direct chemotherapy effect. This finding should ideally be confirmed in analysis of patient-matched pre- and post-chemotherapy specimens, although the latter are not ubiquitous in most centers.
Stromal HA expression in primary OC was reported to be associated with clinicopathologic parameters of aggressive disease, as well as poor PFS and OS, the latter finding retaining an independent prognostic role in multivariate analysis [
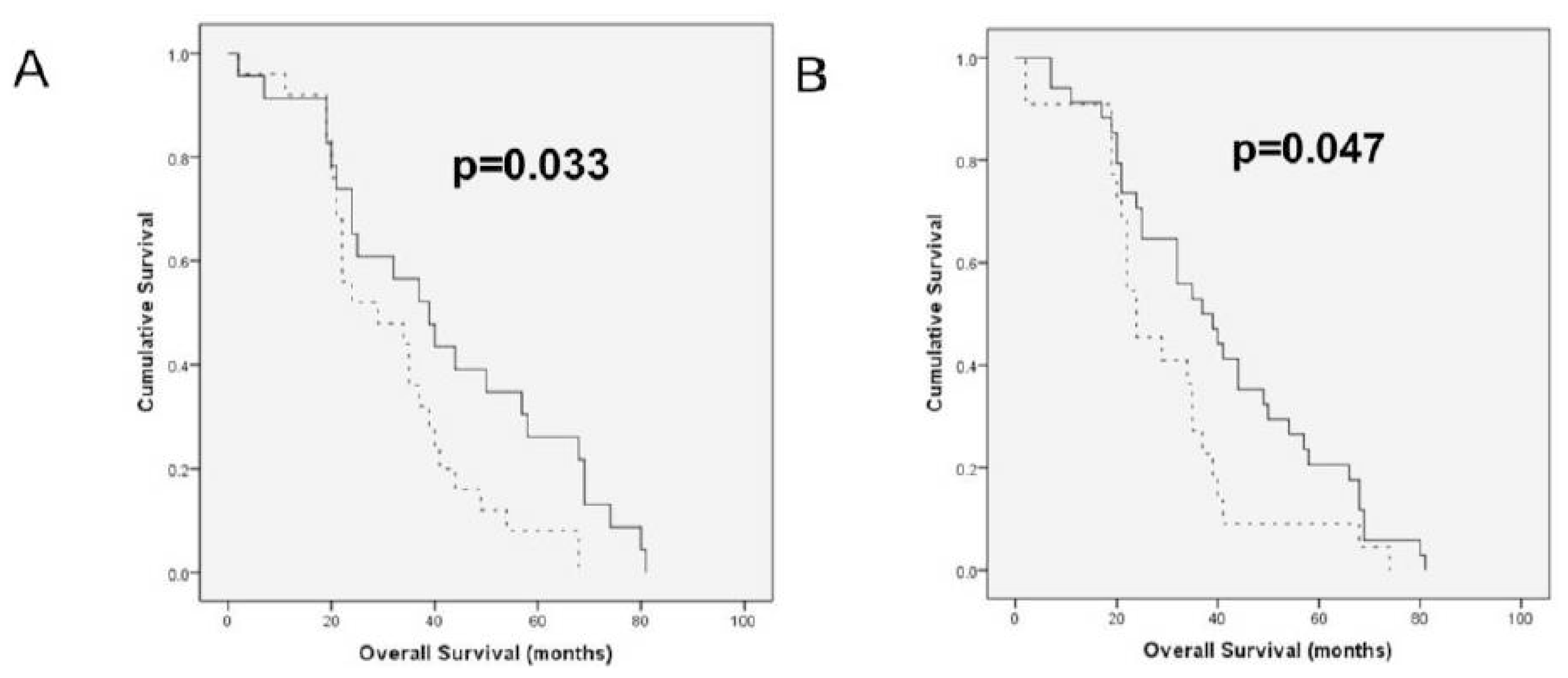
20]. In the only analysis of the prognostic role of HAS members to date, HAS1 protein expression by immunohistochemistry in a series of 33 primary OC was related to significantly shorter survival, whereas HAS2 and HAS3 had no prognostic role. In agreement with this report, higher HAS1 splice variant expression in OC effusions was significantly related to shorter OS. This suggests that the clinical relevance of HAS1 is retained along disease progression. In another view of the inverse relationship between HAS and Hyal members, higher HYAL2-var1 mRNA expression in effusions was associated with longer OS. Although these findings did not retain its significance in Cox analysis, a larger study may yet identify these molecules as independent prognostic markers. The association of HAS and Hyal members with shorter and longer OS, respectively, suggests that HA synthesis is associated with more aggressive disease in metastatic OC, with opposite results for HA breakdown, well in agreement with the role of HA in metastasis and tumor progression in OC and in other cancers.
4. Materials and Methods
4.1. Patients and Material
A total of 97 specimens from patients diagnosed with ovarian, peritoneal or tubal serous carcinoma were obtained from the Department of Gynecological Oncology at the Norwegian Radium Hospital from 1998–2003. Due to their closely linked histogenesis and phenotype, all are referred to as OC henceforth. Informed consent was obtained according to national guidelines. The study was approved by the Regional Committee for Medical Research Ethics in Norway.
Effusions: Sixty-one fresh non-fixed malignant peritoneal (n = 43) and pleural (n = 18) effusions were obtained from 56 patients (3 patients with 2 effusions, 1 with 3 effusions). Forty-seven patients had OC, 7 had primary peritoneal carcinoma, and 2 had tubal carcinoma. The majority of patients (48/56; 86%) received platinum-based therapy, most frequently (42 patients) in combination with paclitaxel.
Effusions were submitted for routine diagnostic purposes and were processed immediately after tapping. Cell blocks were prepared using the Thrombin clot method. Diagnoses were established using morphology and immunohistochemistry. Effusion specimens were centrifuged immediately after tapping, and cell pellets were frozen at −70 °C in equal amounts of RPMI 1640 medium (GIBCO-Invitrogen, Carlsbad CA) containing 50% fetal calf serum (PAA Laboratories GmbH, Pasching, Austria) and 20% dimethylsulfoxide (Merck KGaA, Darmstadt, Germany). Smears and H&E-stained cell block sections were reviewed by a surgical pathologist experienced in cytopathology (BD). The tumor cell population was >50% in all specimens.
Biopsies: Thirty-six solid specimens (27 primary serous carcinomas, 9 metastases) were studied for comparative purposes. Metastases were from the omentum (n = 7), uterus (n = 1) or lymph node (=1). Specimens were not patient-matched, with the exception of one paired effusion and primary carcinoma, one paired effusion and solid metastases, and 2 metastases from a third patient (omentum and lymph node).
Clinicopathologic data were available for 25 of the 27 patients with primary OC. Histological grade was as follows: low-grade: 4 tumors; high-grade: 21 tumors. One patient was diagnosed with FIGO stage II disease, 18 at stage III and 6 at stage IV. Twenty-two tumors were obtained prior to chemotherapy exposure, and 3 were obtained after neoadjuvant chemotherapy. Data regarding residual disease volume were available for 23 patients, of whom 8 were debulked to 0 cm macroscopic disease, 6 to 1 cm and 9 to ≥ 2 cm.
Surgical specimens were submitted for routine diagnostic purposes, snap-frozen and kept at −70 °C with no medium. Frozen sections from all tumors were evaluated for the presence of a >50% tumor component and absence of necrosis. H&E-stained sections were reviewed by a gynecological pathologist (BD) to establish tumor type and histological grade.
4.2. RT-PCR Analysis
Total RNA was extracted using a commercial kit (Tri Reagent; Sigma-Aldrich, St. Louis, MO, USA), and 0.5 μg total RNA were reverse-transcribed using M-MLV Reverse Transcriptase (Promega, Madison, WI, USA) with incubation for 2 h at 37 °C, followed by 5 min at 95 °C, and diluted to 1:5 with RNase-free water. The RT-PCR analysis was performed on cDNA samples on a DNA thermal cycler (Eppendorf Mastercycler gradient; Eppendorf, Hamburg, Germany) using primer sets detecting HAS and HYAL transcripts and 28S ribosomal RNA. Primer sequences and cycle parameters are detailed in
Tables 3 and
4, respectively. RNA from HT-1080 fibrosarcoma cells served as control.
Products were separated on 1.5% agarose gels, isolated using the Invisorb® Spin DNA extraction kit (Invitek GmbH, Berlin, Germany) and sequenced. Gels were photographed by the KODAK EDAS 290 system. Densitometer analysis of films was performed using a computerized image analysis (NIH IMAGE 1.63) program. mRNA levels were established by calculating the target molecule/28S ratio (all cases scored for band intensity compared to control). Expression intensity of ≤5% of control levels was interpreted as negative. Measurements were made at the linear phase of the reaction.
4.3. Quantitative PCR (qPCR)
mRNA levels of HYAL2, 28S and the reference gene RPLPO were analyzed in 84 specimens (52 effusions, 25 primary OC, 7 solid metastases) by qRT-PCR. cDNA was amplified by qPCR in 18 μL reactions using the DyNAmo™ SYBR
® Green qPCR Kit with ROX™ passive reference dye (Finnzymes Oy, Espoo, Finland), with 200 nM of sense and antisense primers. The cycling program was according to the manufacturer’s instructions. The reaction was performed on the Mx3000P
® QPCR System (Stratagene). Oligonucleotide primers were designed by the Primer Express program (Applied Biosystems, Foster City, CA, USA). Primer sequence and assay conditions are detailed in
Tables 5 and
6, respectively.
4.4. Statistical Analysis
Data were analyzed using the SPSS-PC package, version 18.0 (Chicago, IL, USA). Comparative analysis of HAS and HYAL mRNA expression level in effusions, primary carcinomas and solid metastases were performed using the Kruskal-Wallis H test. Analyses of the association between expression levels and clinicopathologic parameters for patients with effusions were performed using the Mann-Whitney U test. For this analysis, clinicopathologic parameters were grouped as follows: Age: ≤60 vs. >60 years; effusion site: peritoneal vs. pleural; FIGO stage: III vs. IV; chemotherapy status: pre- vs. post-chemotherapy specimens; RD volume (≤1 cm vs. >1 cm); response to chemotherapy for primary disease and for disease recurrence: complete vs. partial response/stable disease/progression. Histological grade was not included in these analyses as the majority of patients had high-grade tumors.
Progression-free survival (PFS) and OS were calculated from the date of the last chemotherapy treatment/diagnosis to the date of recurrence/death or last follow-up, respectively. Univariate survival analyses of PFS and OS were executed using the Kaplan-Meier method and log-rank test. Expression categories in survival analyses were clustered as higher vs. lower than median expression. Multivariate analysis of OS was performed using the Cox proportional hazard model (Method = Enter).





{kind=link}
{kind=link}
{kind=link}