2.2. Effect of VHb Expression on Biomass at Lower Temperatures
Cell growth of VHb
+ and VHb
− strains incubated at 250 rpm in shake-flasks under two temperatures of 30 °C and 23 °C were compared to study the effects of VHb expression and lower temperature cultures on
P. pastoris biomass. As shown in
Figure 1A, the two strains growth during the first 48 h of 30 °C cultures were almost identical. After glucose as well as ethanol completely depleted (
Figure 1B) and then methanol added at 48 h, the differences in cell growth occurred immediately and increased with culture time caused by induced VHb expression. The maximal biomass of VHb
+ and VHb
− strains reached 13.7 and 11.4 mg-DCW/mL respectively, which was enhanced 20% by the effect of VHb expression. Both strains also displayed identical cell growth during the first 72 h of 23 °C cultures, thereafter, their growth differences significantly increased with time. VHb
+ and VHb
− strains can grow to maximal values of 16.0 and 11.2 mg-DCW/mL respectively, which was a substantial increase of 43% by VHb effect. Therefore, VHb effect on biomass was positive at both temperatures, however, the effect was more pronounced at 23 °C.
Since lower temperature cultures are not favorable for
P. pastoris growth, the biomass of two strains during the first 72 h of the 23 °C cultures were only half of those at 30 °C. The glucose consumption rates of two strains at 23 °C were also slower than that at 30 °C (
Figure 1B,C). However, VHb
− strain cultured at two temperatures can reach the same final cell density with different kinetics. At 30 °C most of biomass was produced before methanol induction and growth ceased after induction, while at 23 °C growth continued for a prolonged time. For VHb
+ strain, growth was faster after induction than for the VHb
− strain at 30 °C as well as at 23 °C, leading to more biomass produced. After 144 h of 23 °C cultures cell density of VHb
+ strain surpassed itself at 30 °C, and at the end of cultures biomass was increased by 17%. Thus, VHb expression in
P. pastoris at lower temperatures can further enhance final cell density. Additionally, growth phase was prolonged significantly by VHb expression, resulting in enhanced biomass accumulation. This implies that substrates of glucose and methanol were used with better efficiency as to increasing the yield coefficient
Yx/
s, since no remaining substrates concentrations were detected at the end of cultures.
The viabilities of VHb
+ and VHb
− strains at both temperatures were also compared. As shown in
Figure 1D, the differences in the viabilities of two strains increased with culture time at both temperatures. The decreasing rates of both strains were slower at 23 °C and eventually obtained higher viabilities than that at 30 °C. At 23 °C the viabilities of VHb
+ and VHb
− strains at the end of 216 h of cultures were 96.0% and 93.1% respectively, whereas at 30 °C they were 95.2% and 88.4% respectively. Evidently, VHb expression can benefit the viability at both temperatures. Additionally, the effect of lower temperature cultures on cell viability was higher in VHb
− strain than in VHb
+ strain. This is due to the viability of VHb
+ strain was already over 95%, thus a further increase was more difficult. Lower temperature cultures can reduce cell death as to increase cell viability in this study. The phenomenon is also observed in other
P. pastoris studies [
10,
11], probably because AOX1 activity is higher at lower temperatures so that the toxicity of methanol to cells can be reduced [
8].
2.3. Analysis of VHb Protein at Two Temperatures
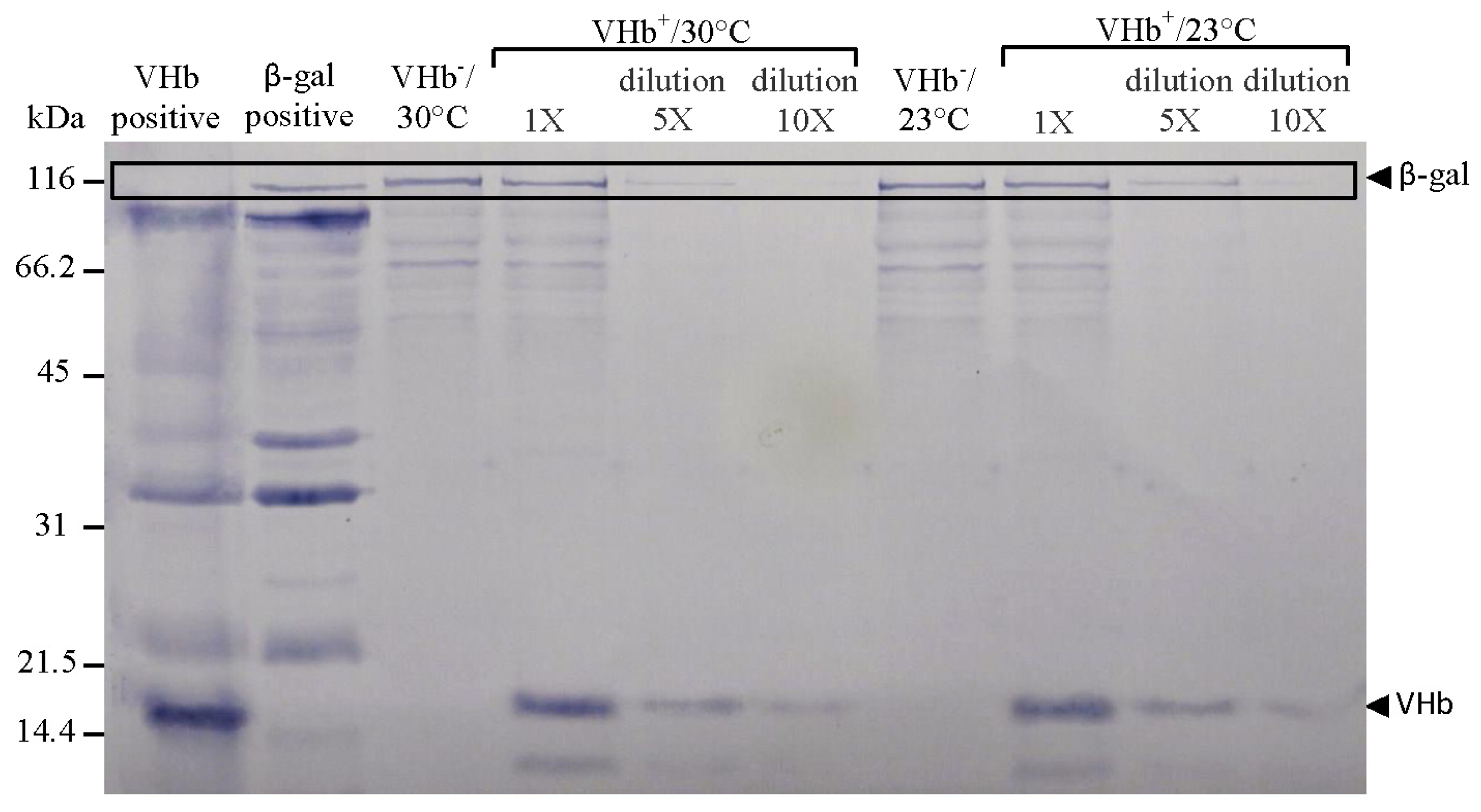
It proved worthwhile to investigate why VHb effect on biomass was higher at 23 °C than at 30 °C. First, VHb expression levels were analyzed. Cells were harvested at the end of 216 h of cultures and adjusted to the same OD
600 before being disrupted. Western-blotting technique was then used to confirm the expressed protein and evaluate protein expression levels. Unexpectedly, the levels of expressed VHb polypeptides (globin portion) at two temperatures were nearly equal (
Figure 2). This finding differed from what we first thought, namely, 23 °C cultures could enhance VHb expression (primarily based on the studies reporting that lower temperature cultures can improve recombinant protein expression in
P. pastoris [
10–
12]) as to increase VHb effect on biomass. Evidently, VHb expression cannot be increased by lower temperature cultures.
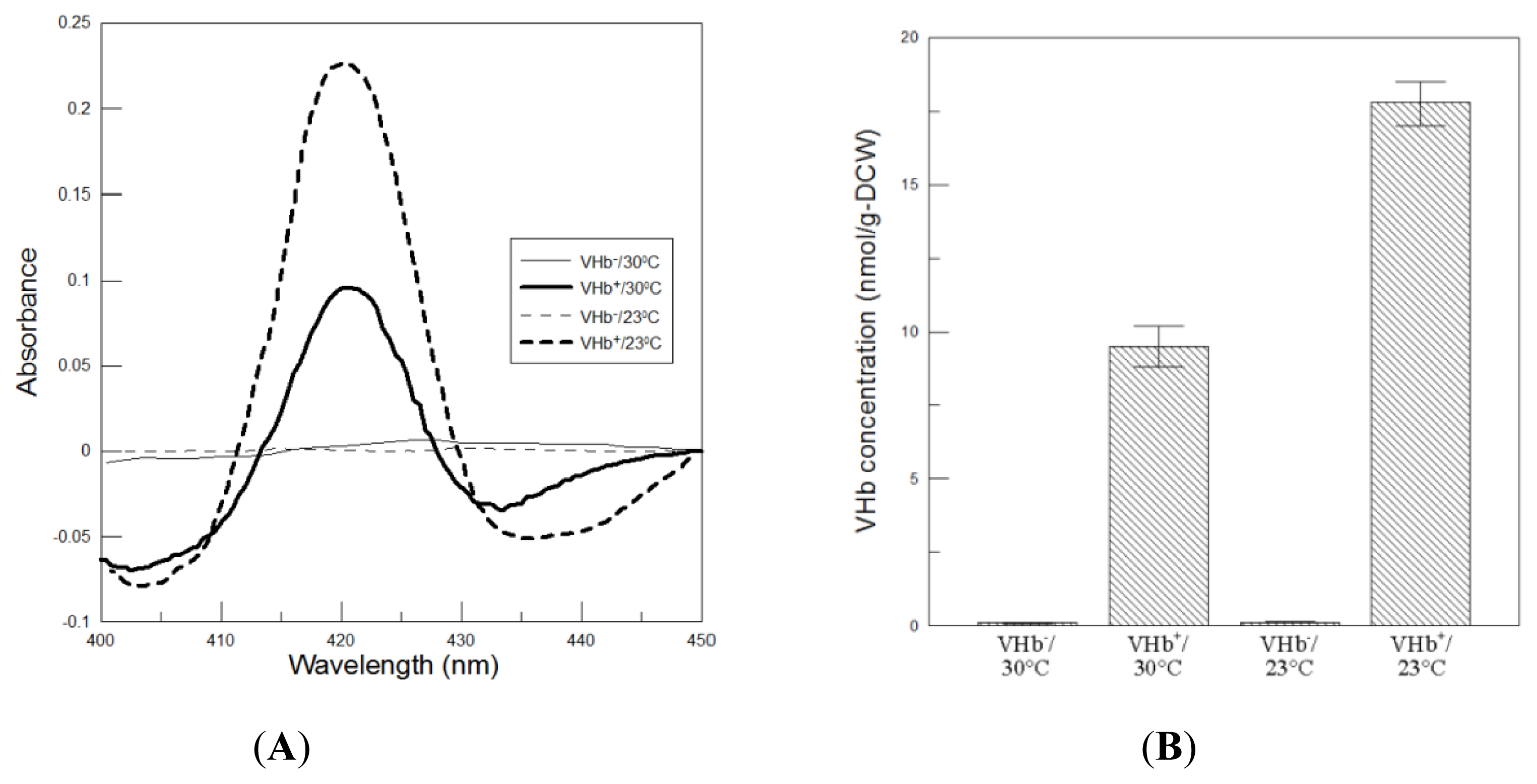
Subsequently, VHb activity was determined using the CO-difference spectra assay under 25 °C. Samples were reduced with dithionite in the presence and absence of CO. Then the treated samples were scanned on a spectrophotometer. As seen in
Figure 3A, the difference spectra of VHb
+ strain showed a peak in the Soret region at 419 nm, which is characteristic of active VHb. Suddenly, VHb activity obtained at the end of 216 h of 23 °C cultures was 17.8 nmol/g-DCW, which was approximately twofold higher than that obtained at 30 °C (
Figure 3B). The finding was in favorable agreement with the fact that the color of total cell lysates of VHb
+ strain cultured at 23 °C was redder than itself at 30 °C (
Figure 3C). This implies that at 30 °C probably there was a significant fraction of VHb that lacked the essential cofactor of heme group. As a result, lower temperature cultures cannot improve VHb expression but can significantly increase VHb activity.
In addition, the VHb expressed at both temperatures exhibited a spectrum characteristic of the oxygenated form. The phenomenon was also observed previously in
Vitreoscilla [
13] and
E. coli [
14] that the expressed VHb existed in the physiologically active ferrous oxygenated form. In our study, the spectrum of the oxygenated VHb obtained from two temperature cultivations had a Soret peak at 415 nm and α and β bands at 580 and 550 nm, respectively (
Figure 3D), which represented the hexacoordinated ferrous heme state. After reduced with dithionite (the deoxygenated form) the Soret peak shifted to 425 nm and one broad peak at 562 nm replaced the α and β bands, indicating the pentacoordinated ferrous heme state. The heights of α and β peaks obtained from 23 °C cultivation were more intense than that at 30 °C.
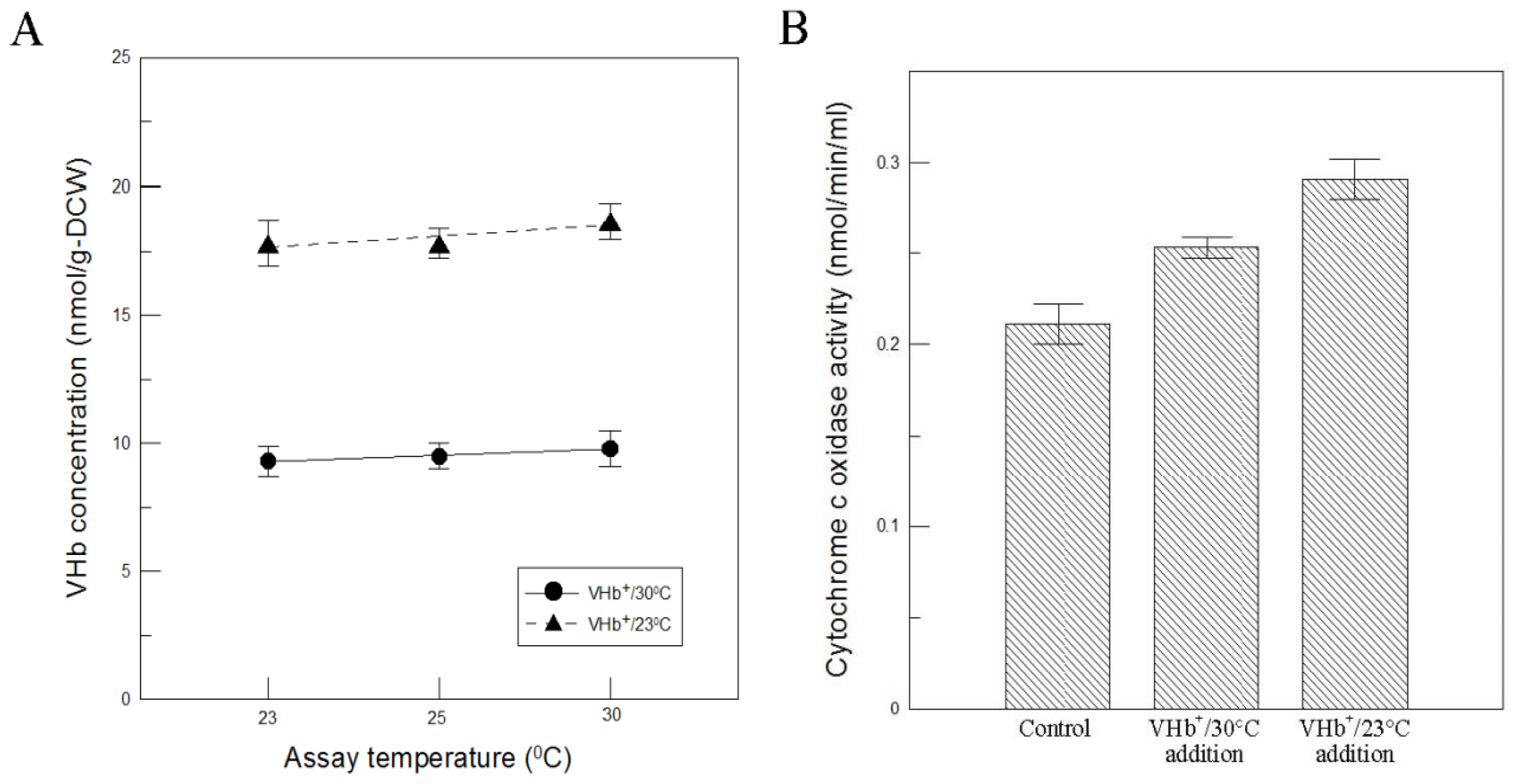
To clarify if the above results could be caused by the VHb reactivity being more favorable only at lower temperatures, VHb activity was determined at different reaction temperatures. The results shown in
Figure 4A indicated that VHb activity obtained at two temperatures cultures did not obviously change within the assay temperatures between 30 °C and 23 °C, thus this reason can be ruled out. Additionally, the exogenous provision of VHb to the assay conditions of the oxygen-consuming enzyme (cytochrome c oxidase) was also carried out to examine if VHb can increase the oxidase activity. First, the cleared lysates of VHb
+ cells were incubated at 42 °C for 6 h to inactivate cytochrome c oxidase activity. Since cytochrome c oxidase is sensitive to temperatures, whereas VHb can endure temperatures of up to 80 °C [
15]. The VHb-containing samples were then added to the cleared lysates of freshly disrupted VHb
− cells to measure cytochrome c oxidase activity. As shown in
Figure 4B, VHb-containing samples obtained at 23 °C cultures increased cytochrome c oxidase activity by 0.08 nmol min
−1 mL
−1, which was twice that obtained at 30 °C cultures. The result was consistent with that shown in
Figure 3 and confirmed again that VHb activity obtained at 23 °C cultures was twofold higher than that at 30 °C cultures. It has been proven that VHb can improve respiratory efficiency and ATP synthesis in recombinant hosts [
1]. Due to twofold VHb activity achieved, it is reasonable to obtain more biomass and higher viability by VHb
+ strain at 23 °C than at 30 °C.
Our results showed that VHb expression level at 23 °C was equal to that at 30 °C, but VHb activity obtained at 23 °C was twofold higher than that at 30 °C. A similar phenomenon has also been reported in
E. coli where total globin protein synthesis was barely affected by reducing the temperature from 30 °C to 26 °C, whereas lower temperature cultures produced a twofold higher soluble rHb1.1 than higher temperature cultures did [
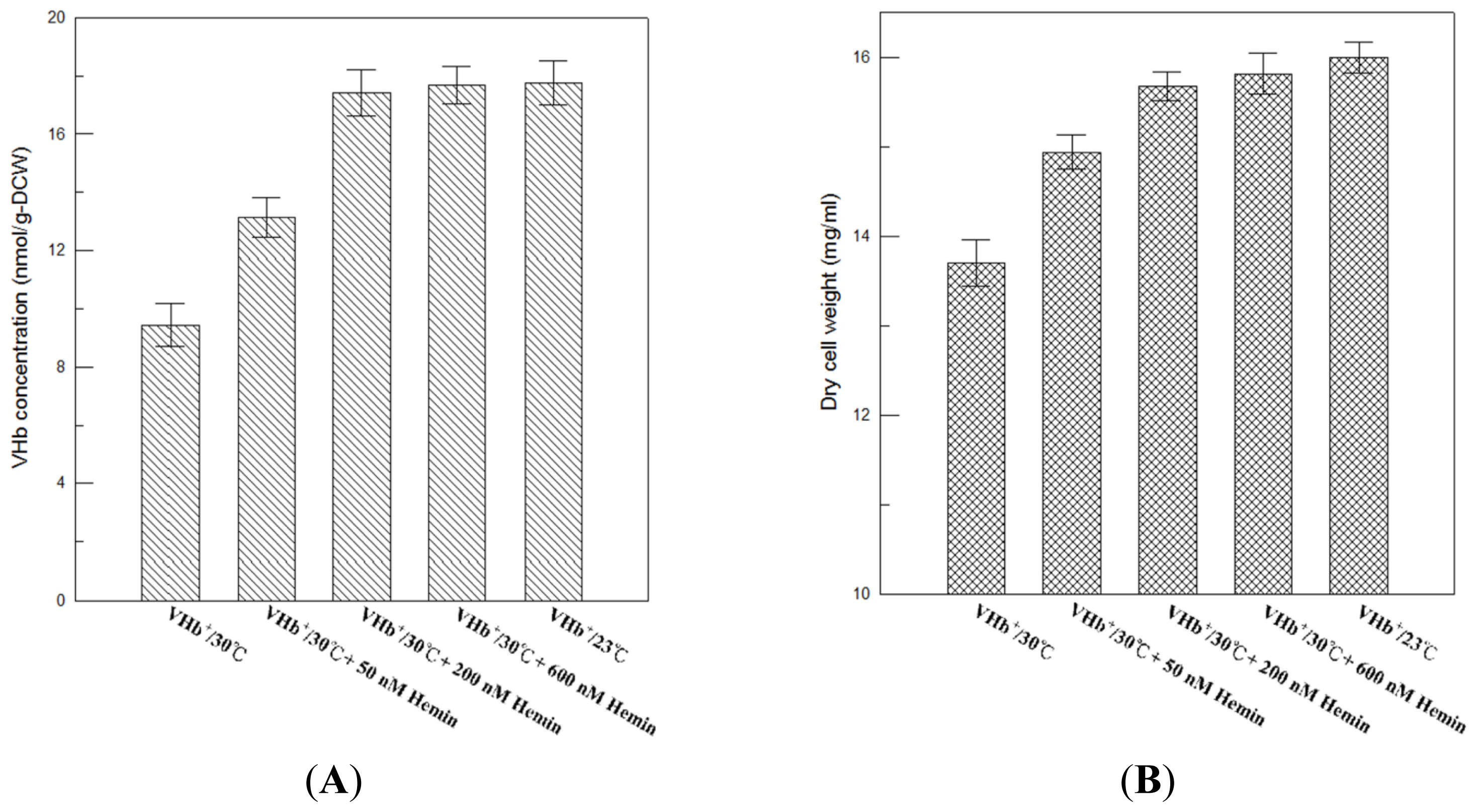
16]. A significant difference between active VHb at 23 °C and 30 °C could be caused by improper VHb folding that lower temperature cultures can reduce protein synthesis rate, improve protein folding, and increase protein solubility, or due to inadequate heme incorporation that lower temperature cultures could favor heme biosynthesis so that improve heme incorporation, folding, and subunit assembly. In order to determine exactly why in the difference, the exogenous provision of hemin to 30 °C culture broth of VHb
+ strain was performed to examine if the availability of heme is lower at 30 °C in
P. pastoris. As shown in
Figure 5A, VHb activity of VHb
+ strain at the end of 216 h of 30 °C cultures increased with hemin concentration and saturated at 200 nM hemin supplement, which was the same level as that of 23 °C cultures (17.8 nmol/g-DCW). This reveals that the amount of heme in
P. pastoris at 30 °C was lower than that at 23 °C, resulting in lower VHb activity at 30 °C than 23 °C. Therefore, a different heme production in
P. pastoris caused this twofold difference between active VHb at 23 °C and 30 °C. In addition, a similar effect of hemin on final cell concentration was observed (
Figure 5B). Increase of VHb activity caused by the exogenous provision of hemin also improved final cell concentration, and the final biomass of 30 °C culture supplemented with 200 nM hemin was close to that of 23 °C culture.
Lower temperature cultures can increase the availability of oxygen and ATP in
P. pastoris. Evidences of higher intracellular ATP levels and increased the ATP regeneration rates [
17] were obtained at lower temperature cultures. The saturated dissolved oxygen concentration of YPD medium at 23 °C is 7.12 ppm, which is 17% higher than that at 30 °C (6.08 ppm). Increased amounts of oxygen and ATP could affect the flux of several important biosynthetic routes in
P. pastoris. Increased oxygen concentration can improve the respiration rate and thereby enhanced the oxygen uptake rates (
Figure 7). Oxygen tension also influences the folding and assembly of hemeproteins by affecting the availability or redox state of heme. In addition, oxygen is required for two steps in heme biosynthesis, the formation of protoporphyrinogen IX by coproporphyrinogen III oxidase, and the formation of protoporphyrin IX by protoporphyrinogen IX oxidase. The step catalyzed by coproporphyrinogen III oxidase is rate limiting [
18]. VHb is a homodimeric protein, and each protomer incorporates a heme prosthetic group in its native state. VHb requires the presence of adequate heme production during VHb protein biosynthesis and folding. Higher amounts of heme could be produced at 23 °C than at 30 °C because lower temperature cultures can increase the availability of dissolved oxygen and ATP, which are used in several steps of heme biosynthesis [
18]. Higher availability of heme could favor heme incorporation, folding, and subunit assembly of VHb protein. As a result, more holoVHb (VHb protein carrying heme) could be obtained at 23 °C and resulted in twofold higher VHb activity than that at 30 °C. However, more apoVHb (VHb protein lacking heme) could be produced at 30 °C due to lower availability of heme. The statement also can be confirmed by the result of
Figure 3C that more VHb missing heme at 30 °C resulted in a great difference in red color compared to that of 23 °C.
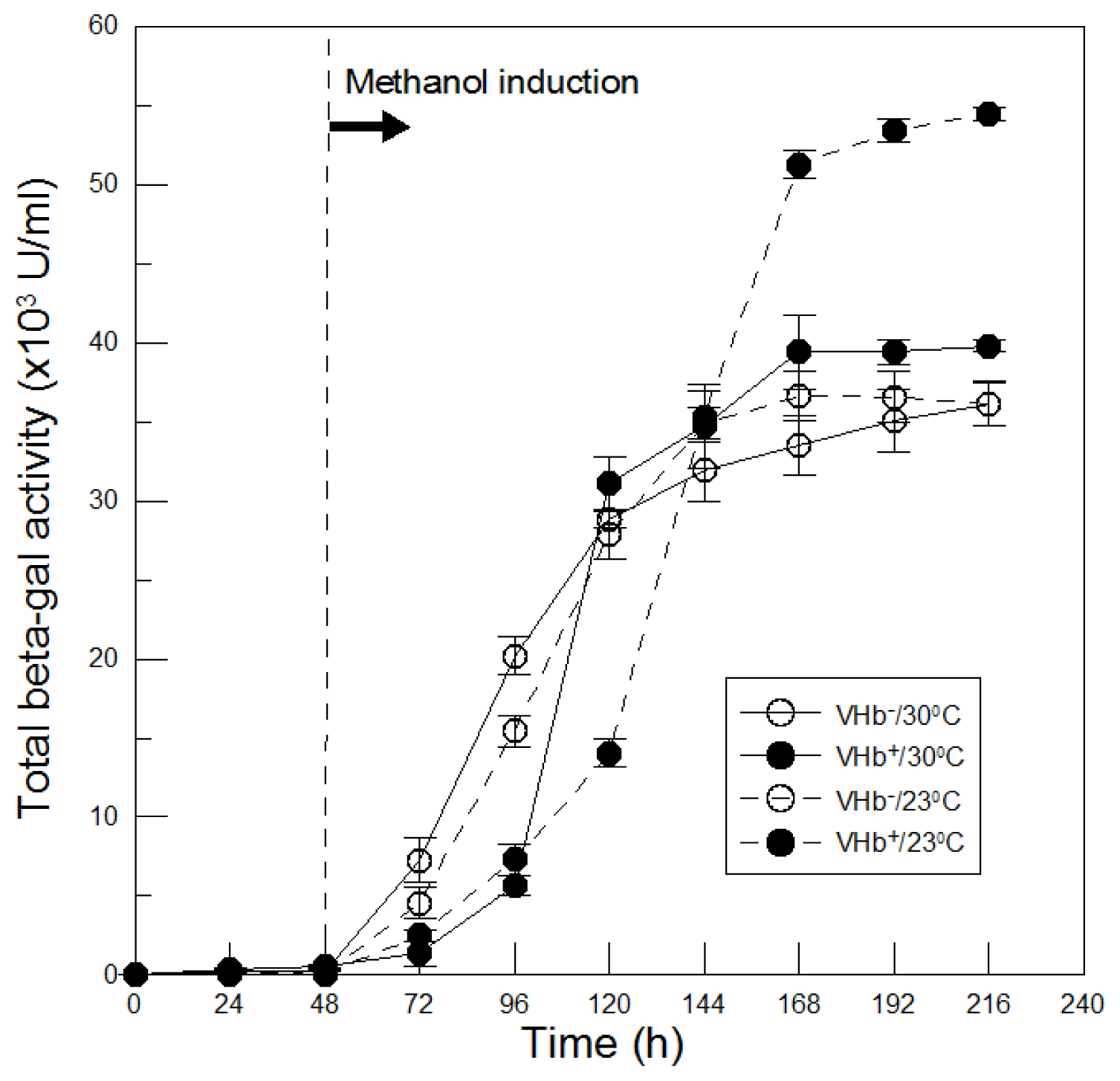
2.4. Effect of VHb Expression on Recombinant β-Galactosidase Production at Lower Temperatures
The differences in recombinant β-galactosidase production between VHb
+ and VHb
− strains at two temperatures were compared. As shown in
Figure 6, before induction no β-galactosidase activity was detected in both strains at two temperatures. However, after the absence of glucose (
Figure 1B,C) and thereby methanol induction at 48 h of cultures, β-galactosidase began to express. At 30 °C β-galactosidase of VHb
− strain was produced immediately and increased with time, leading to a maximal value of 36,000 U/mL achieved at the end of 216 h of cultures. While β-galactosidase of VHb
+ strain differed from that of VHb
− strain. In the early methanol induction phase, β-galactosidase production of VHb
+ strain was slower than that of VHb
− strain. This could be since recombinant VHb protein was co-induced by methanol in VHb
+ strain; VHb consumed some of cellular resources that could be used for β-galactosidase biosynthesis. However, expressed VHb protein could compensate VHb
+ strain for improving β-galactosidase production. Therefore, after 120 h of cultures β-galactosidase production of VHb
+ strain exceeded that of VHb
− strain and then reached a maximal activity of 40,000 U/mL at the end of cultures, which was enhanced 11% by VHb effect.
β-Galactosidase production of VHb
− strain at 23 °C was analogous to itself at 30 °C and their maximal activities were nearly equivalent. This indicates that β-galactosidase produced by VHb
− strain was independent of culture temperatures, which differed from some previous studies reporting that lower temperature cultures can improve target protein production in
P. pastoris [
10–
12]. Therefore, the effect of lower temperature cultures on recombinant protein production depends on different target proteins. Comparing β-galactosidase produced at 23 °C by two strains, β-galactosidase production of VHb
+ strain was also slower in the early methanol induction phase due to co-induced two proteins as mentioned above. Later, however, VHb effect significantly accelerated β-galactosidase production and eventually reached a maximal activity of 55,000 U/mL, which was 50% much higher than that of VHb
− strain. Additionally, after 144 h of cultures β-galactosidase production of VHb
+ strain at 23 °C surpassed itself at 30 °C, and finally obtained an increase of 38% at the end of cultures. This enhancement production of β-galactosidase is not due to lower temperature cultures improving β-galactosidase production but caused by twofold VHb activity obtained at 23 °C cultures.
The results demonstrated that 23 °C cultures further increased VHb activity as to enhance VHb effect on β-galactosidase production as well as on biomass. For various proteins expression in different hosts, culture temperatures cannot usually satisfy both the biomass and protein expression simultaneously. But it can be achieved in this study by culturing P. pastoris at 23 °C and obtained a significant higher biomass and β-galactosidase production than that at 30 °C, primarily due to VHb function.
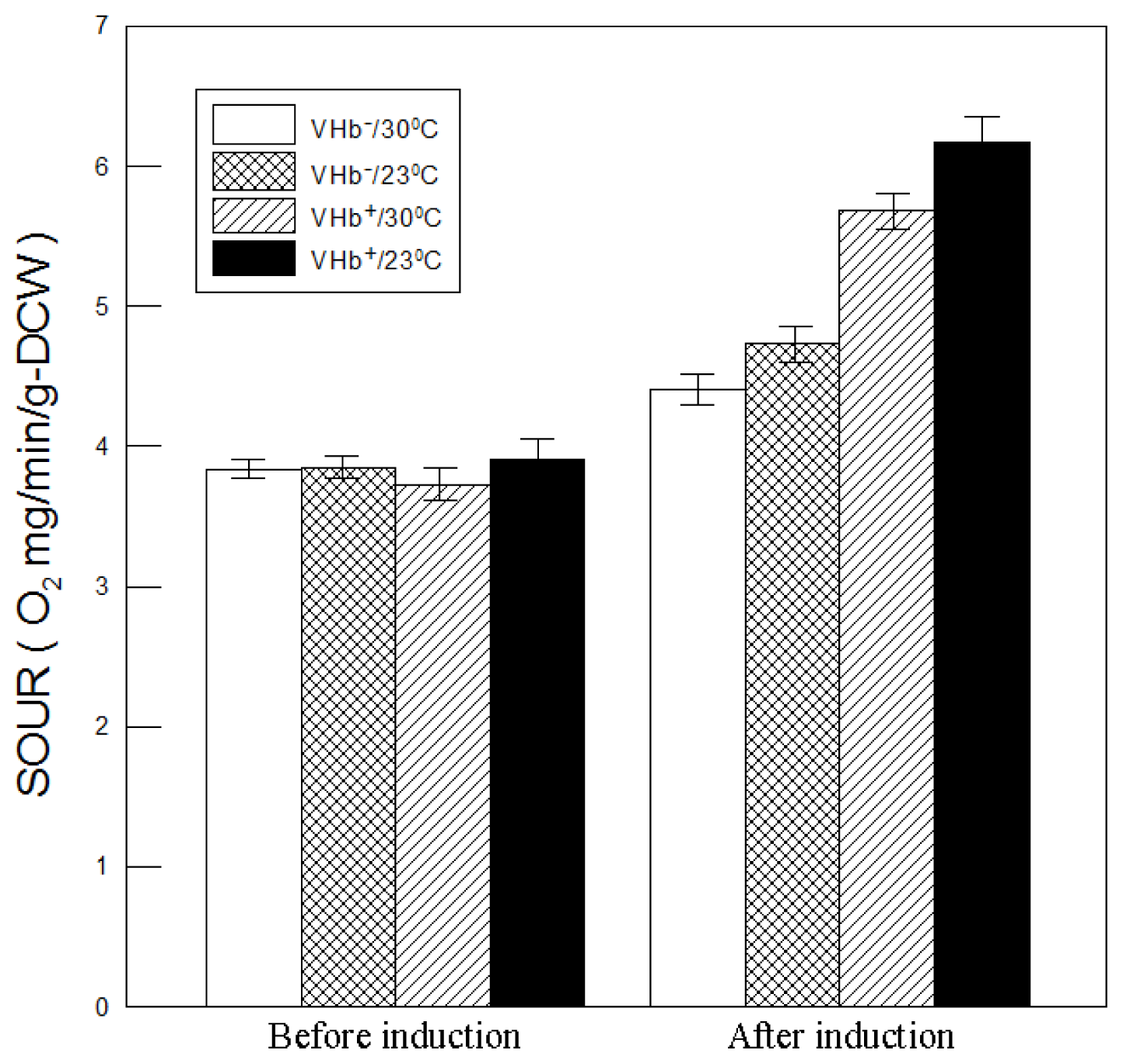
2.5. Effect of VHb Expression on Oxygen Uptake Rates at Lower Temperatures
VHb
+ strain capable of higher biomass and β-galactosidase production implies that it could have a higher oxygen uptake rate. To explore this possibility, the specific oxygen uptake rates (SOURs) were measured. As shown in
Figure 7, SOURs of both strains at two temperatures before methanol induction exhibited no obvious differences (around 3.8 O
2 mg min
−1 g-DCW
−1). After induction, however, all SOURs rose to peak values of much higher than 3.8 O
2 mg min
−1 g-DCW
−1 due to an increased oxygen demand from cell growth and methanol metabolization, which is a high oxygen-consuming process. SOURs of VHb
− strain cultured at 30 °C and 23 °C increased from 3.8 before induction to 4.4 and 4.7 O
2 mg min
−1 g-DCW
−1 after induction, respectively. While that of VHb
+ strain rose respectively to 5.6 and 6.3 O
2 mg min
−1 g-DCW
−1. The considerably higher increases in VHb
+ strain than VHb
− strain were probably caused by VHb expression improving oxygen utilization and respiratory efficiency. SOURs of VHb
+ strain at 30 °C and 23 °C were respectively 27% and 34% higher than that of VHb
− strain, indicating that VHb effect on SOURs was beneficial at both temperatures but higher at 23 °C. The VHb-expressing strain with a higher oxygen demand has also been reported previously [
19–
21].
Moreover, after induction VHb
− strain had a higher SOUR at 23 °C than at 30 °C, suggesting that cells at 23 °C demonstrated higher respiratory efficiency and metabolic activity. This is because the oxygen-consuming enzyme AOX1 activity is higher at lower temperatures [
12], which thus can reduce the toxicity of methanol to cells and thereby increase cell viability, as shown in
Figure 1D. The phenomenon was also observed in VHb
+ strain but its SOUR increase (0.56 O
2 mg min
−1 g-DCW
−1) was twice as that of VHb
− strain (0.25 O
2 mg min
−1 g-DCW
−1), which was likely caused by twofold VHb activity obtained at 23 °C cultures.
Overall, VHb+ strain had a higher SOUR at 23 °C than at 30 °C, indicating that cell performance of VHb+ strain was better at 23 °C. It can reasonably explain why VHb+ strain at 23 °C had a higher final cell density, cell viability, and β-galactosidase production. Consequently, lower temperature cultures can enlarge VHb effect on cell performance of P. pastoris.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







