A Simple Bioconjugate Attachment Protocol for Use in Single Molecule Force Spectroscopy Experiments Based on Mixed Self-Assembled Monolayers

Abstract
:1. Introduction
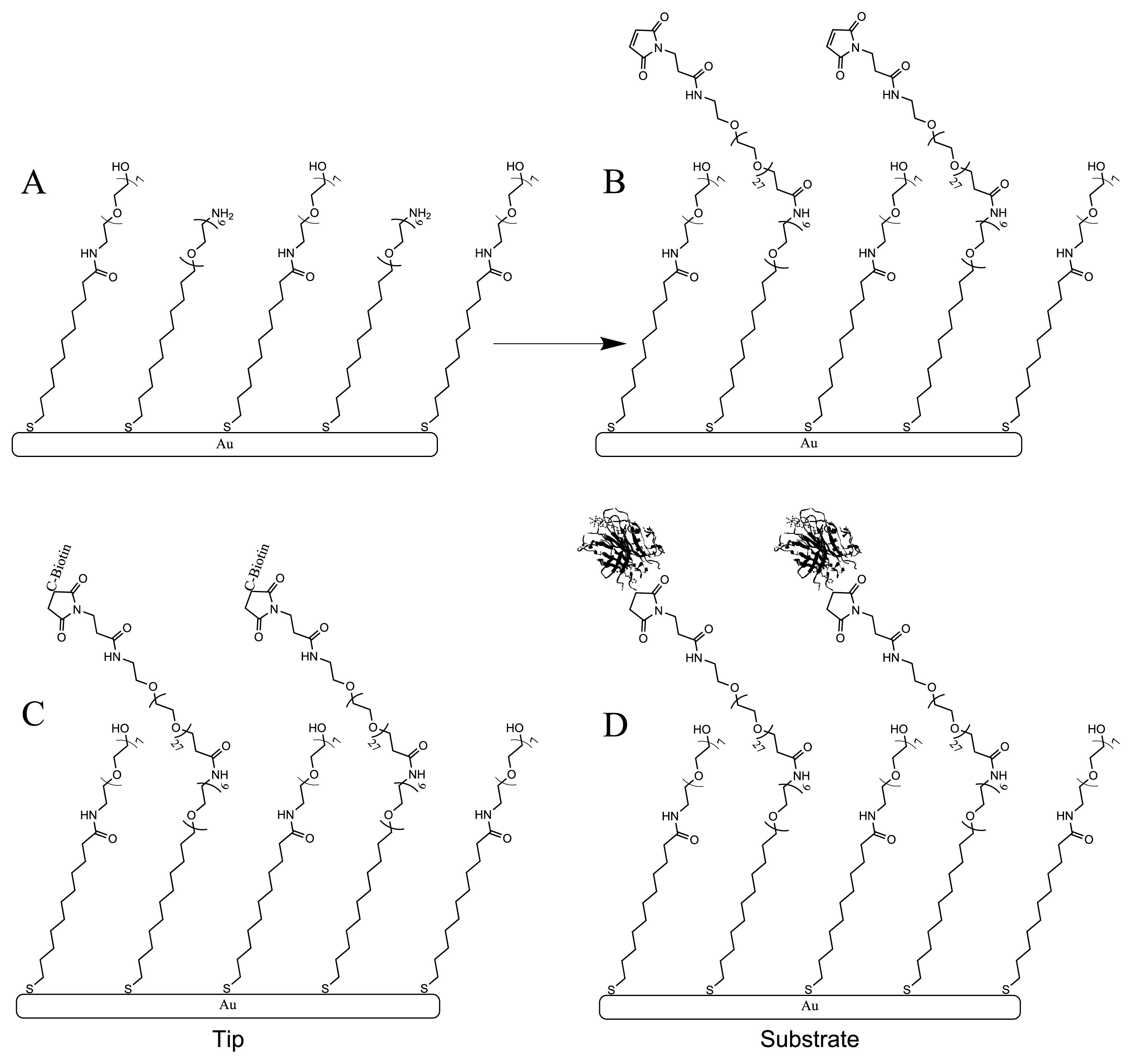
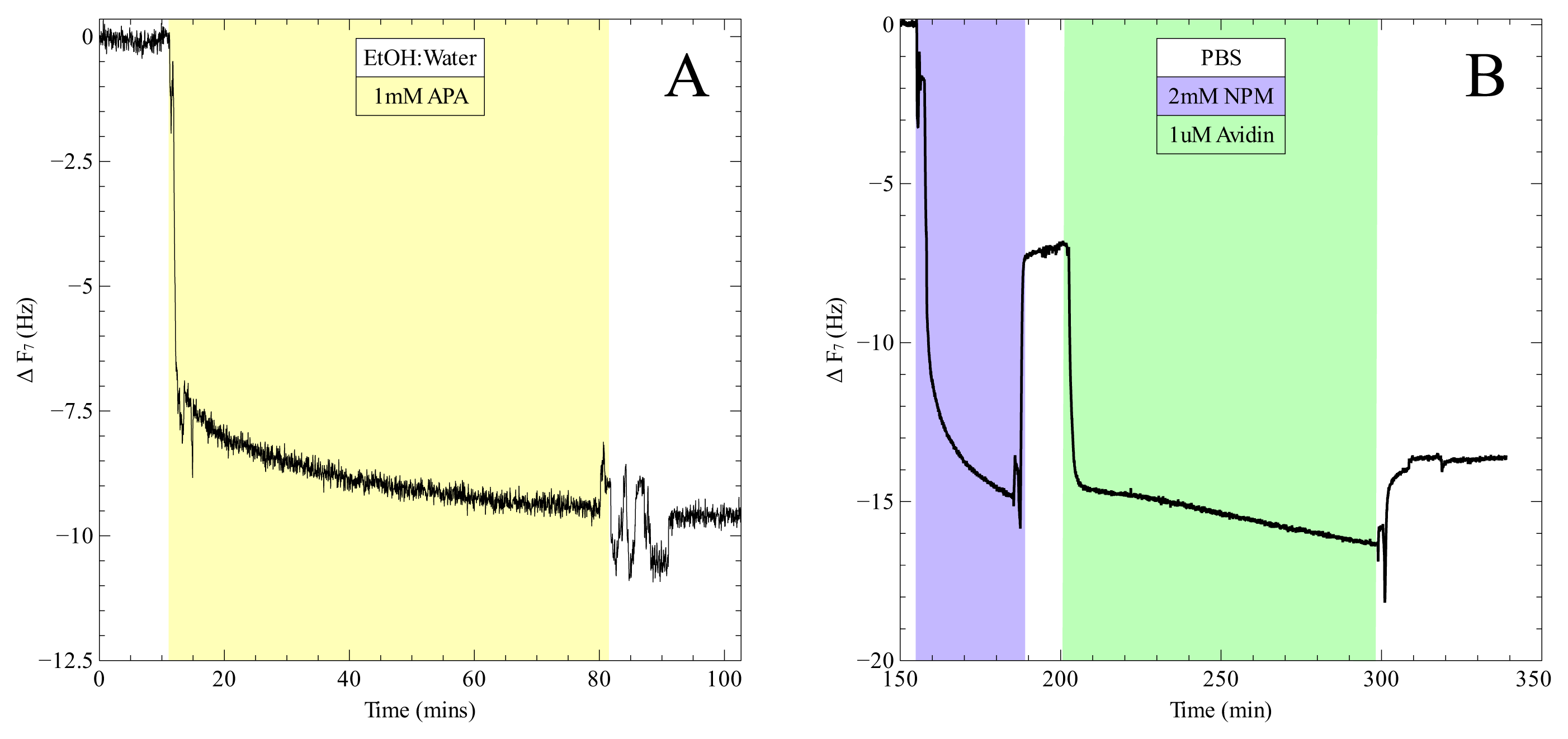
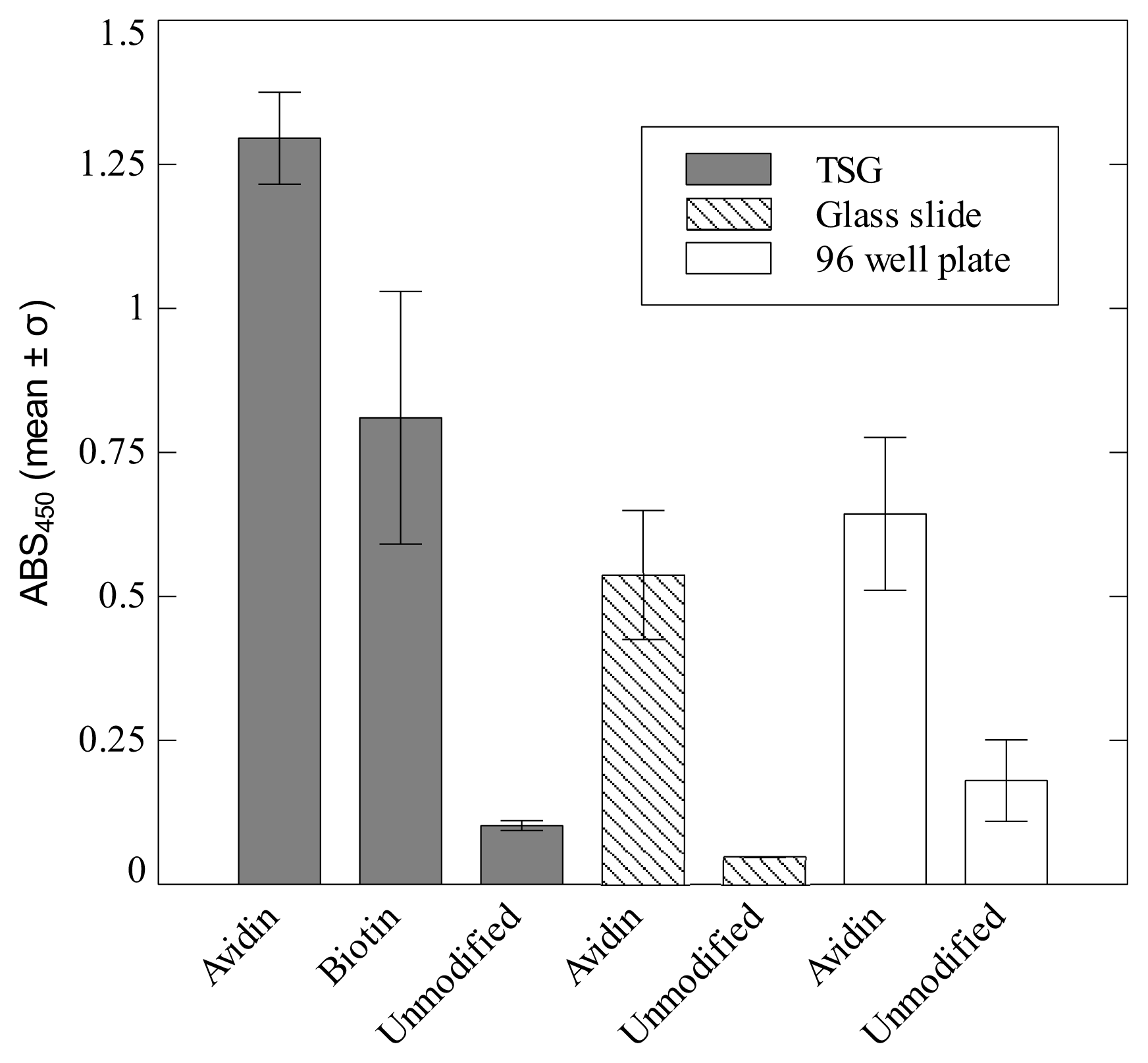
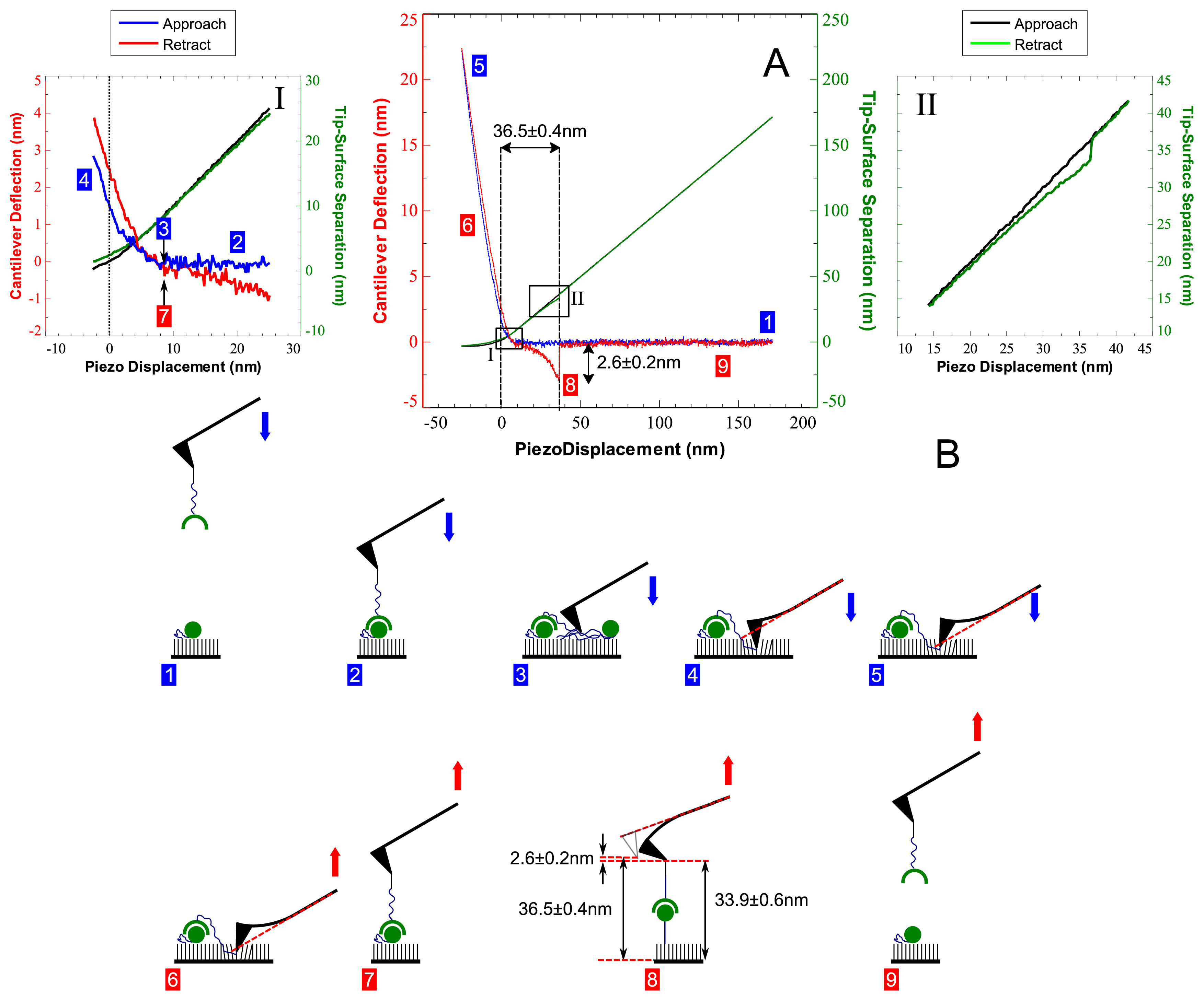
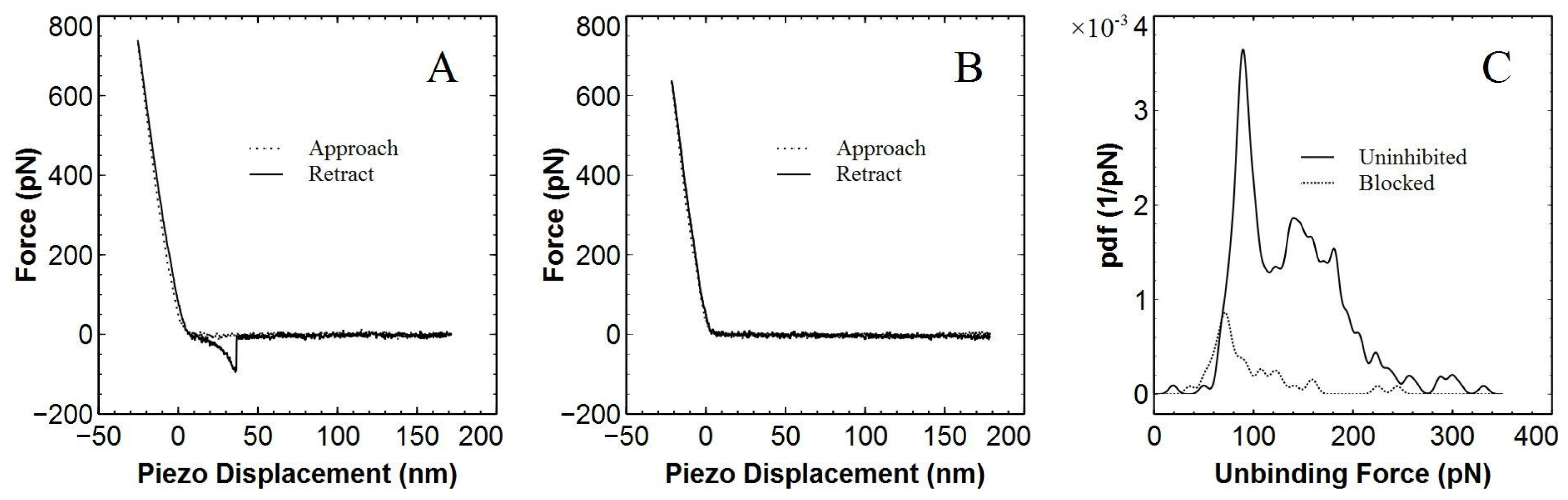
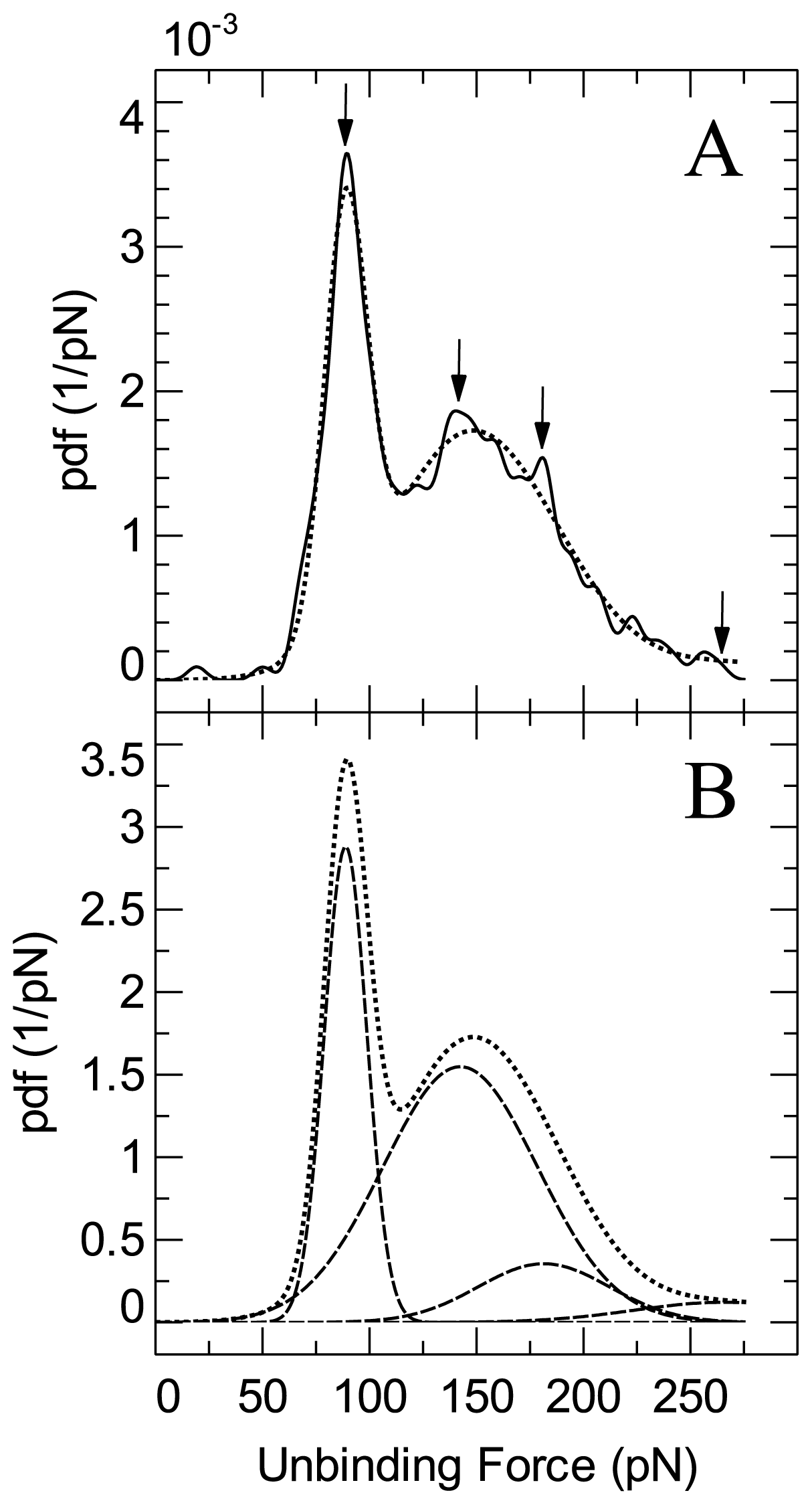
2. Results and Discussion
3. Experimental Section
3.1. Materials and Instrumentation
3.2. Preparation of Template-Stripped Gold (TSG) Surfaces
3.3. Monolayer Preparation
Substrate
Cantilevers
3.4. Enzyme-linked Immunosorbent Assay (ELISA)
Standard adhesion assay
Modified adhesion assay
3.5. Atomic Force Microscopy
3.6. Quartz Crystal Microbalance Assay
4. Conclusions
Acknowledgments
References
- Binnig, G.; Quate, C.; Gerber, C. Atomic Force Microscope. Phys. Rev. Lett 1986, 56, 930–933. [Google Scholar]
- Florin, E.; Moy, V.; Gaub, H. Adhesion forces between individual ligand-receptor pairs. Science 1994, 264, 415–417. [Google Scholar]
- Evans, E.; Ritchie, K. Dynamic strength of molecular adhesion bonds. Biophys. J 1997, 72, 1541–1555. [Google Scholar]
- Merkel, R.; Nassoy, P.; Leung, A.; Ritchie, K.; Evans, E. Energy landscapes of receptor-ligand bonds explored with dynamic force spectroscopy. Nature 1999, 397, 50–53. [Google Scholar]
- De Paris, R.; Strunz, T.; Oroszlan, K.; Guntherodt, H.J. Force spectroscopy and dynamics of the biotin-avidin bond studied by scanning force microscopy. Single Mol 2000, 1, 285–290. [Google Scholar]
- Yuan, C.; Chen, A.; Kolb, P.; Moy, V. Energy landscape of streptavidin-biotin complexes measured by atomic force microscopy. Biochemistry 2000, 39, 10219–10223. [Google Scholar]
- Piramowicz, M.; Czuba, P.; Targosz, M.; Burda, K.; Szymonski, M. Dynamic force measurements of avidin-biotin and streptavdin-biotin interactions using AFM. Acta Biochim. Pol 2006, 53, 93–100. [Google Scholar]
- Lo, Y.; Huefner, N.; Chan, W.; Stevens, F.; Harris, J.; Beebe, T. Specific interactions between biotin and avidin studied by atomic force microscopy using the Poisson statistical analysis method. Langmuir 1999, 15, 1373–1382. [Google Scholar]
- Lo, Y.; Simons, J.; Beebe, T. Temperature dependence of the biotin-avidin bond-rupture force studied by atomic force microscopy. J. Phys. Chem. B 2002, 106, 9847–9852. [Google Scholar]
- Ebner, A.; Wildling, L.; Kamruzzahan, A.S.M.; Rankl, C.; Wruss, J.; Hahn, C.D.; Hoelzl, M.; Zhu, R.; Kienberger, F.; Blaas, D.; et al. A new, simple method for linking of antibodies to atomic force microscopy tips. Bioconjug. Chem 2007, 18, 1176–1184. [Google Scholar]
- Kamruzzahan, A.S.M.; Ebner, A.; Wildling, L.; Kienberger, F.; Riener, C.K.; Hahn, C.D.; Pollheimer, P.D.; Winklehner, P.; Hoelzl, M.; Lackner, B.; et al. Antibody linking to atomic force microscope tips via disulfide bond formation. Bioconjug. Chem 2006, 17, 1473–1481. [Google Scholar]
- Riener, C.; Stroh, C.; Ebner, A.; Klampfl, C.; Gall, A.; Romanin, C.; Lyubchenko, Y.; Hinterdorfer, P.; Gruber, H. Simple test system for single molecule recognition force microscopy. Anal. Chim. Acta 2003, 479, 59–75. [Google Scholar]
- Chilkoti, A.; Boland, T.; Ratner, B.; Stayton, P. The relationship between ligand-binding thermodynamics and protein-ligand interaction forces measured by atomic force microscopy. Biophys. J 1995, 69, 2125–2130. [Google Scholar]
- Lo, Y.; Zhu, Y.; Beebe, T. Loading-rate dependence of individual ligand-receptor bond-rupture forces studied by atomic force microscopy. Langmuir 2001, 17, 3741–3748. [Google Scholar]
- Ludwig, M.; Dettmann, W.; Gaub, H. Atomic force microscope imaging contrast based on molecular recognition. Biophys. J 1997, 72, 445–448. [Google Scholar]
- Wong, S.; Joselevich, E.; Woolley, A.; Cheung, C.; Lieber, C. Covalently functionalized nanotubes as nanometre-sized probes in chemistry and biology. Nature 1998, 394, 52–55. [Google Scholar]
- Wong, J.; Chilkoti, A.; Moy, V. Direct force measurements of the streptavidin-biotin interaction. Biomol. Eng 1999, 16, 45–55. [Google Scholar]
- Lee, G.; Chrisey, L.; Colton, R. Direct measurement of the forces between complementary strands of DNA. Science 1994, 266, 771–773. [Google Scholar]
- Rico, F.; Moy, V.T. Energy landscape roughness of the streptavidin-biotin interaction. J. Mol. Recognit 2007, 20, 495–501. [Google Scholar]
- Berquand, A.; Xia, N.; Castner, D.; Clare, B.; Abbott, N.; Dupres, V.; Adriaensen, Y.; Dufrene, Y. Antigen binding forces of single antilysozyme Fv fragments explored by atomic force microscopy. Langmuir 2005, 21, 5517–5523. [Google Scholar]
- Grandbois, M.; Beyer, M.; Rief, M.; Clausen-Schaumann, H.; Gaub, H. How strong is a covalent bond? Science 1999, 283, 1727–1730. [Google Scholar]
- Zhang, X.; Wojcikiewicz, E.; Moy, V. Force spectroscopy of the leukocyte function-associated antigen-1/intercellular adhesion molecule-1 interaction. Biophys. J 2002, 83, 2270–2279. [Google Scholar]
- Nevo, R.; Stroh, C.; Kienberger, F.; Kaftan, D.; Brumfeld, V.; Elbaum, M.; Reich, Z.; Hinterdorfer, P. A molecular switch between alternative conformational states in the complex of Ran and importin beta 1. Nat. Struct. Biol 2003, 10, 553–557. [Google Scholar]
- Baumgartner, W.; Hinterdorfer, P.; Ness, W.; Raab, A.; Vestweber, D.; Schindler, H.; Drenckhahn, D. Cadherin interaction probed by atomic force microscopy. Proc. Natl. Acad. Sci. USA 2000, 97, 4005–4010. [Google Scholar]
- Zhang, X.; Bogorin, D.; Moy, V. Molecular basis of the dynamic strength of the sialyl Lewis X-selectin interaction. ChemPhysChem 2004, 5, 175–182. [Google Scholar]
- Livnah, O.; Bayer, E.; Wilchek, M.; Sussman, J. 3-dimensional structures of avidin and the avidin- biotin complex. Proc. Natl. Acad. Sci. USA 1993, 90, 5076–5080. [Google Scholar]
- Pincet, F.; Husson, J. The solution to the streptavidin-biotin paradox: The influence of history on the strength of single molecular bonds. Biophys. J 2005, 89, 4374–4381. [Google Scholar]
- Strunz, T.; Oroszlan, K.; Schumakovitch, I.; Guntherodt, H.; Hegner, M. Model energy landscapes and the force-induced dissociation of ligand-receptor bonds. Biophys. J 2000, 79, 1206–1212. [Google Scholar]
- Grubmuller, H.; Heymann, B.; Tavan, P. Ligand binding: Molecular mechanics calculation of the streptavidin biotin rupture force. Science 1996, 271, 997–999. [Google Scholar]
- Bizzarri, A.R.; Cannistraro, S. The application of atomic force spectroscopy to the study of biological complexes undergoing a biorecognition process. Chem. Soc. Rev 2010, 39, 734–749. [Google Scholar]
- Guo, S.; Ray, C.; Kirkpatrick, A.; Lad, N.; Akhremitchev, B.B. Effects of multiple-bond ruptures on kinetic parameters extracted from force spectroscopy measurements: Revisiting biotin-streptavidin interactions. Biophys. J 2008, 95, 3964–3976. [Google Scholar]
- Allen, S.; Chen, X.; Davies, J.; Davies, M.; Dawkes, A.; Edwards, J.; Roberts, C.; Sefton, J.; Tendler, S.; Williams, P. Detection of antigen-antibody binding events with the atomic force microscope. Biochemistry 1997, 36, 7457–7463. [Google Scholar]
- Fritz, J.; Katopodis, A.; Kolbinger, F.; Anselmetti, D. Force-mediated kinetics of single P-selectin ligand complexes observed by atomic force microscopy. Proc. Natl. Acad. Sci. USA 1998, 95, 12283–12288. [Google Scholar]
- Bonanni, B.; Kamruzzahan, A.; Bizzarri, A.; Rankl, C.; Gruber, H.; Hinterdorfer, P.; Cannistraro, S. Single molecule recognition between Cytochrome C 551 and gold-immobilized Azurin by force spectroscopy. Biophys. J 2005, 89, 2783–2791. [Google Scholar]
- Clifford, C.; Seah, M. The determination of atomic force microscope cantilever spring constants via dimensional methods for nanomechanical analysis. Nanotechnology 2005, 16, 1666–1680. [Google Scholar]
- Cleveland, J.; Manne, S.; Bocek, D.; Hansma, P. A Nondestructive Method For Determining The Spring Constant Of Cantilevers For Scanning Force Microscopy. Rev. Sci. Instrum 1993, 64, 403–405. [Google Scholar]
- Hutter, J.; Bechhoefer, J. Calibration of Atomic-Force Microscope Tips. Rev. Sci. Instrum 1993, 64, 1868–1873. [Google Scholar]
- Sader, J.; Chon, J.; Mulvaney, P. Calibration of rectangular atomic force microscope cantilevers. Rev. Sci. Instrum 1999, 70, 3967–3969. [Google Scholar]
- Baumgartner, W.; Hinterdorfer, P.; Schindler, H. Data analysis of interaction forces measured with the atomic force microscope. Ultramicroscopy 2000, 82, 85–95. [Google Scholar]
- Kuhn, M.; Janovjak, H.; Hubain, M.; Muller, D. Automated alignment and pattern recognition of single-molecule force spectroscopy data. J. Microsc.-Oxford 2005, 218, 125–132. [Google Scholar]
- Kienberger, F.; Kada, G.; Gruber, H.; Pastushenko, V.; Riener, C.; Trieb, M.; Knaus, H.G.; Schindler, H.; Hinterdorfer, P. Recognition force spectroscopy studies of the NTA-His6 bond. Single Mol 2000, 1, 59–65. [Google Scholar]
- Ebner, A.; Wildling, L.; Zhu, R.; Rankl, C.; Haselgruebler, T.; Hinterdorfer, P.; Gruber, H.J. Functionalization of Probe Tips and Supports for Single-Molecule Recognition Force Microscopy. In STM and AFM Studies on (Bio)Molecular Systems: Unravelling the Nanoworld; Springer-Verlag: Berlin, Germany, 2008; Volume 285, pp. 29–76. [Google Scholar]
- Hinterdorfer, P.; Kienberger, F.; Raab, A.; Gruber, H.; Baumgartner, W.; Kada, G.; Riener, C.; Wielert-Badt, S.; Borken, C.; Schindler, H. Poly(ethylene glycol): An ideal spacer for molecular recognition force microscopy/spectroscopy. Single Mol 2000, 1, 99–103. [Google Scholar]
- Yadavalli, V.K.; Forbes, J.G.; Wang, K. Functionalized self-assembled monolayers on ultraflat gold as platforms for single molecule force spectroscopy and imaging. Langmuir 2006, 22, 6969–6976. [Google Scholar]
- Zhang, X.; Halvorsen, K.; Zhang, C.Z.; Wong, W.P.; Springer, T.A. Mechanoenzymatic Cleavage of the Ultralarge Vascular Protein von Willebrand Factor. Science 2009, 324, 1330–1334. [Google Scholar]
- Lambert, J.; Jue, R.; Traut, R. Disulfide cross-linking of escherichia-coli ribosomal-proteins with 2-iminothiolane (methyl 4-mercaptobutyrimidate)–evidence that cross-linked protein pairs are formed in intact ribosomal-subunit. Biochemistry 1978, 17, 5406–5416. [Google Scholar]
- Hermanson, G. Bioconjugate Techniques; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Taninaka, A.; Takeuchi, O.; Shigekawa, H. Reconsideration of dynamic force spectroscopy analysis of streptavidin-biotin interactions. Int. J. Mol. Sci 2010, 11, 2134–2151. [Google Scholar]
- Wieland, J.; Gewirth, A.; Leckband, D. Single molecule adhesion measurements reveal two homophilic neural cell adhesion molecule bonds with mechanically distinct properties. J. Biol. Chem 2005, 280, 41037–41046. [Google Scholar]
- Wagner, P.; Hegner, M.; Guntherodt, H.; Semenza, G. Formation and In-Situ Modification of Monolayers Chemisorbed on Ultraflat Template-Stripped Gold Surfaces. Langmuir 1995, 11, 3867–3875. [Google Scholar]
- Rodahl, M.; Hook, F.; Krozer, A.; Brzezinski, P.; Kasemo, B. Quartz-Crystal Microbalance Setup for Frequency and Q-Factor Measurements in Gaseous and Liquid Environments. Rev. Sci. Instrum 1995, 66, 3924–3930. [Google Scholar]
- Rodahl, M.; Hook, F.; Kasemo, B. QCM operation in liquids: An explanation of measured variations in frequency and Q factor with liquid conductivity. Anal. Chem 1996, 68, 2219–2227. [Google Scholar]
- Ebner, A.; Hinterdorfer, P.; Gruber, H.J. Comparison of different aminofunctionalization strategies for attachment of single antibodies to AFM cantilevers. Ultramicroscopy 2007, 107, 922–927. [Google Scholar]
- Rankl, C.; Kienberger, F.; Gruber, H.; Blaas, D.; Hinterdorfer, P. Accuracy estimation in force spectroscopy experiments. Jpn. J. Appl. Phys 2007, 46, 5536–5539. [Google Scholar]
- Kienberger, F.; Pastushenko, V.; Kada, G.; Gruber, H.; Riener, C.; Schindler, H.; Hinterdorfer, P. Static and dynamical properties of single poly(ethylene glycol) molecules investigated by force spectroscopy. Single Mol 2000, 1, 123–128. [Google Scholar]
- Ratto, T.; Rudd, R.; Langry, K.; Balhorn, R.; McElfresh, M. Nonlinearly additive forces in multivalent ligand binding to a single protein revealed with force spectroscopy. Langmuir 2006, 22, 1749–1757. [Google Scholar]
- Hegner, M.; Wagner, P.; Semenza, G. Ultralarge Atomically Flat Template-Stripped AU Surfaces For Scanning Probe Microscopy. Surf. Sci 1993, 291, 39–46. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tip | Experiment | Recognition,% |
|---|---|---|
| A | uninhibited | 27 |
| B | uninhibited | 25 |
| C | uninhibited | 6 |
| D | uninhibited | 15 |
| A | blocked | 4 |
| B | blocked | 8 |
| C | blocked | 10 |
| D | blocked | 6 |
| Reference | Rate regime ( pN s−1) | F*(r) ( pN) | F*(r = 4091) ( pN) |
|---|---|---|---|
| Merkel et al. [4] | 102 to 104 | 14 ln(r) – 38 | ≈ 78 |
| De Paris et al. [5] | 103 to 104 | 15.5 ln(r) – 71.4 | ≈ 56 |
| Yuan et al. [6] | 1000 to 5000 | 20.6 ln(r) – 12.4 | ≈ 157 |
| Piramowicz et al. [7] | 1700 to 9600 | 168.6 ln(r) – 1047.6 | ≈ 355 |
| Current work | 4091 ± 69 | - | 89 ± 13 pN |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Attwood, S.J.; Simpson, A.M.C.; Stone, R.; Hamaia, S.W.; Roy, D.; Farndale, R.W.; Ouberai, M.; E.Welland, M. A Simple Bioconjugate Attachment Protocol for Use in Single Molecule Force Spectroscopy Experiments Based on Mixed Self-Assembled Monolayers. Int. J. Mol. Sci. 2012, 13, 13521-13541. https://doi.org/10.3390/ijms131013521
Attwood SJ, Simpson AMC, Stone R, Hamaia SW, Roy D, Farndale RW, Ouberai M, E.Welland M. A Simple Bioconjugate Attachment Protocol for Use in Single Molecule Force Spectroscopy Experiments Based on Mixed Self-Assembled Monolayers. International Journal of Molecular Sciences. 2012; 13(10):13521-13541. https://doi.org/10.3390/ijms131013521
Chicago/Turabian StyleAttwood, Simon J., Anna M. C. Simpson, Rachael Stone, Samir W. Hamaia, Debdulal Roy, Richard W. Farndale, Myriam Ouberai, and Mark E.Welland. 2012. "A Simple Bioconjugate Attachment Protocol for Use in Single Molecule Force Spectroscopy Experiments Based on Mixed Self-Assembled Monolayers" International Journal of Molecular Sciences 13, no. 10: 13521-13541. https://doi.org/10.3390/ijms131013521
APA StyleAttwood, S. J., Simpson, A. M. C., Stone, R., Hamaia, S. W., Roy, D., Farndale, R. W., Ouberai, M., & E.Welland, M. (2012). A Simple Bioconjugate Attachment Protocol for Use in Single Molecule Force Spectroscopy Experiments Based on Mixed Self-Assembled Monolayers. International Journal of Molecular Sciences, 13(10), 13521-13541. https://doi.org/10.3390/ijms131013521