Subminimal Inhibitory Concentrations of the Disinfectant Benzalkonium Chloride Select for a Tolerant Subpopulation of Escherichia coli with Inheritable Characteristics

Abstract
:1. Introduction
2. Results and Discussion
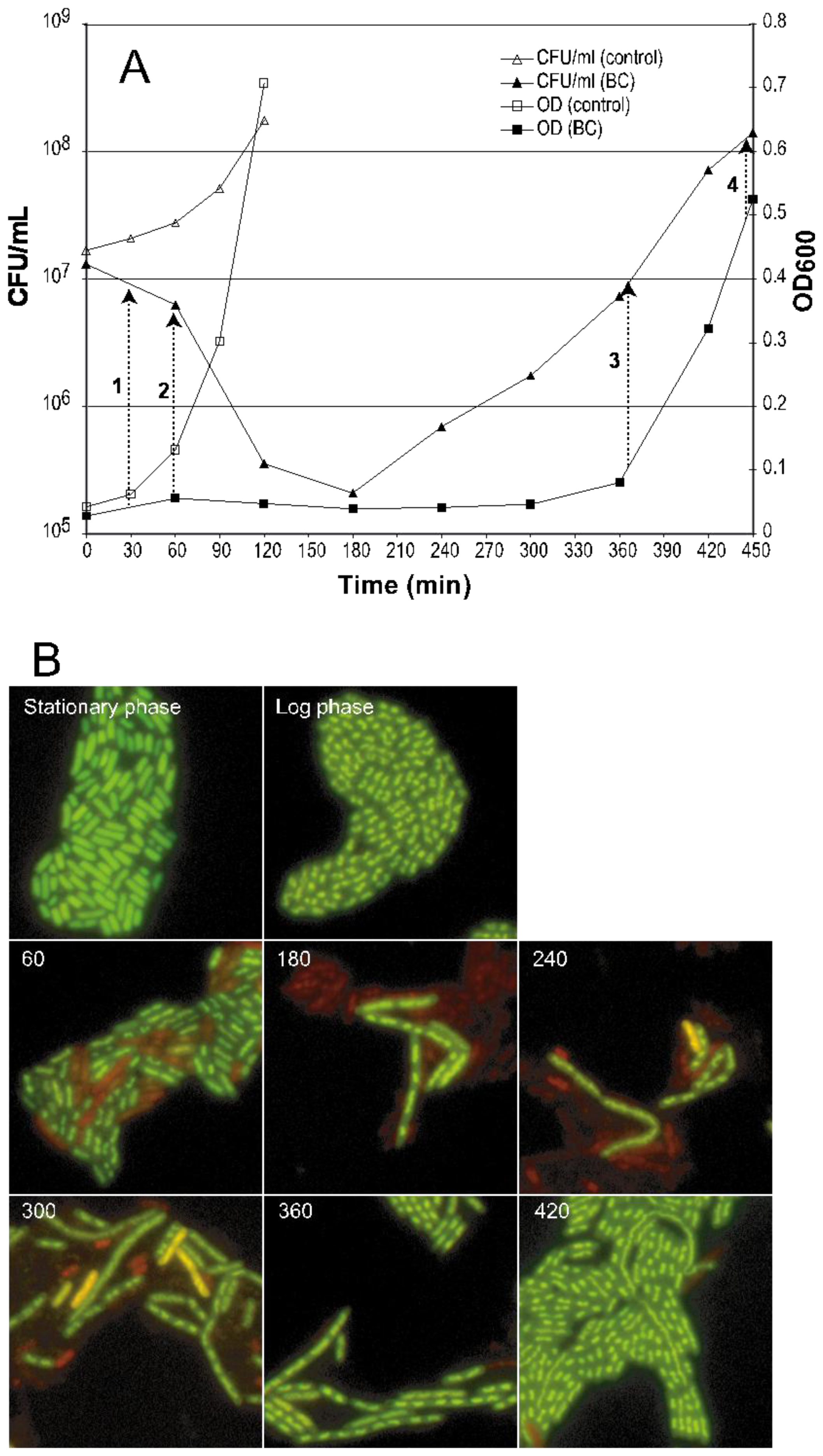
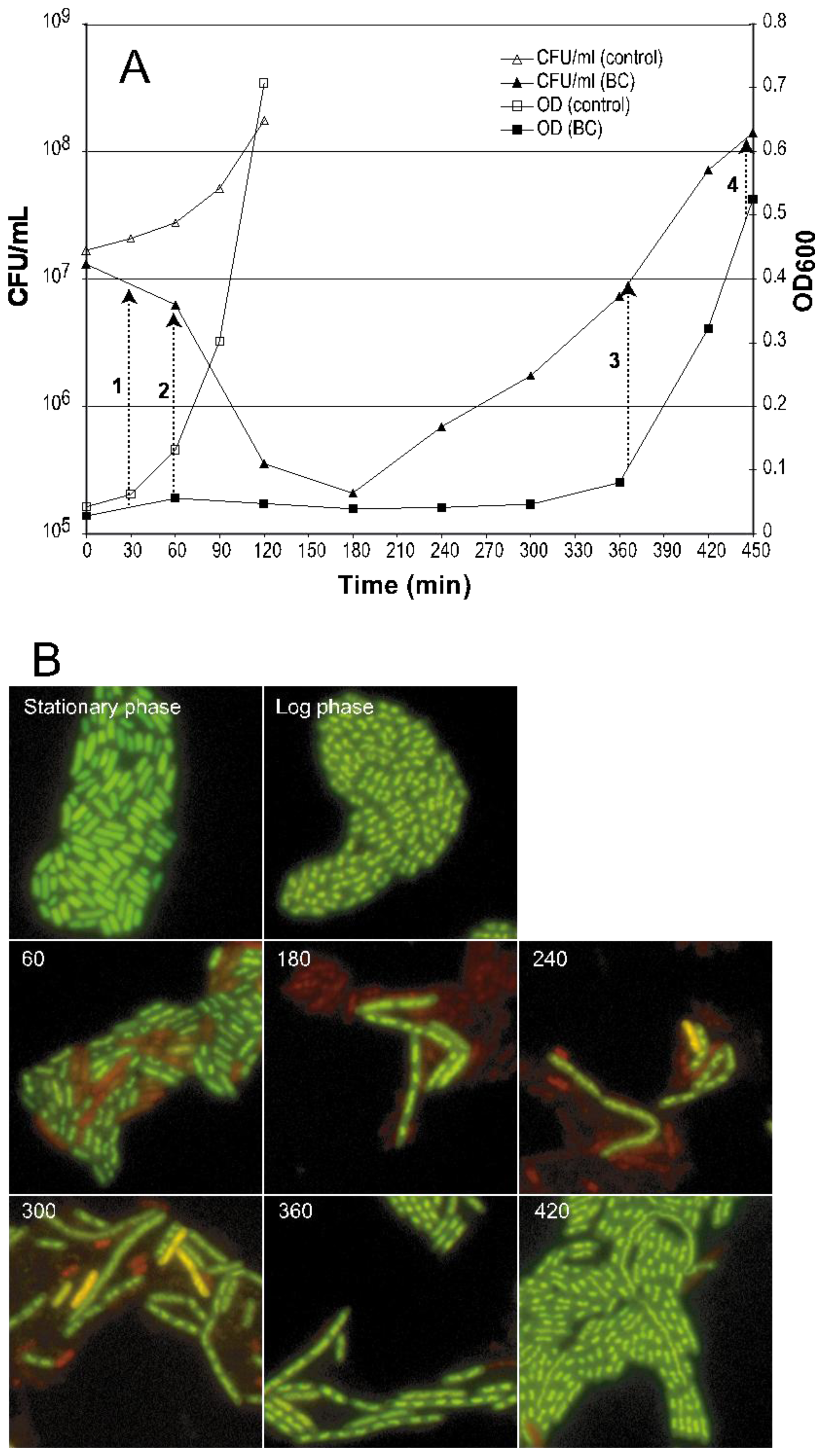
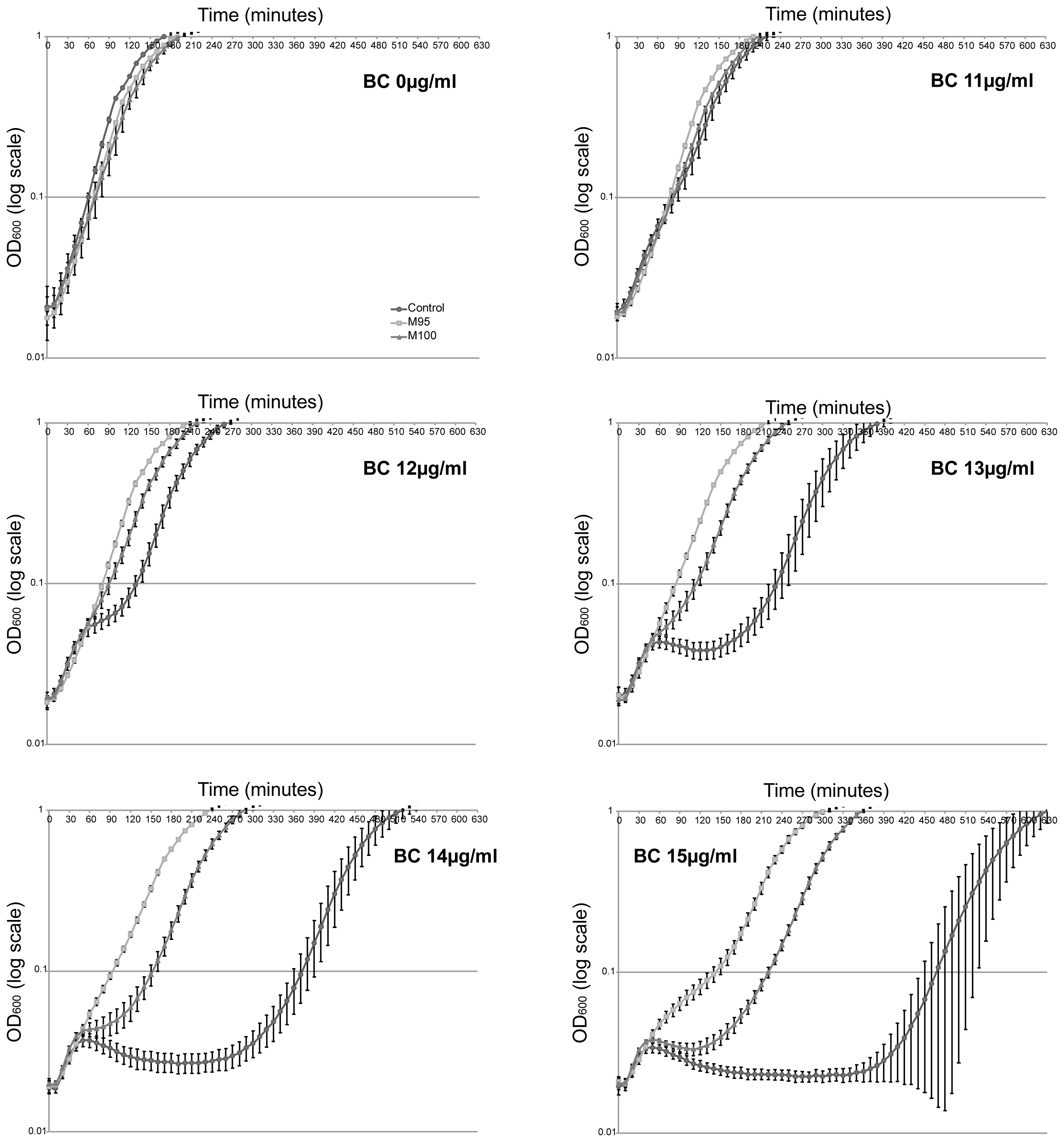
2.1. Morphological Changes and Cultivability as Analyzed by CFU after BC Exposure
2.2. Microarray Analyses
2.3. Susceptibility of Knock-out Mutants
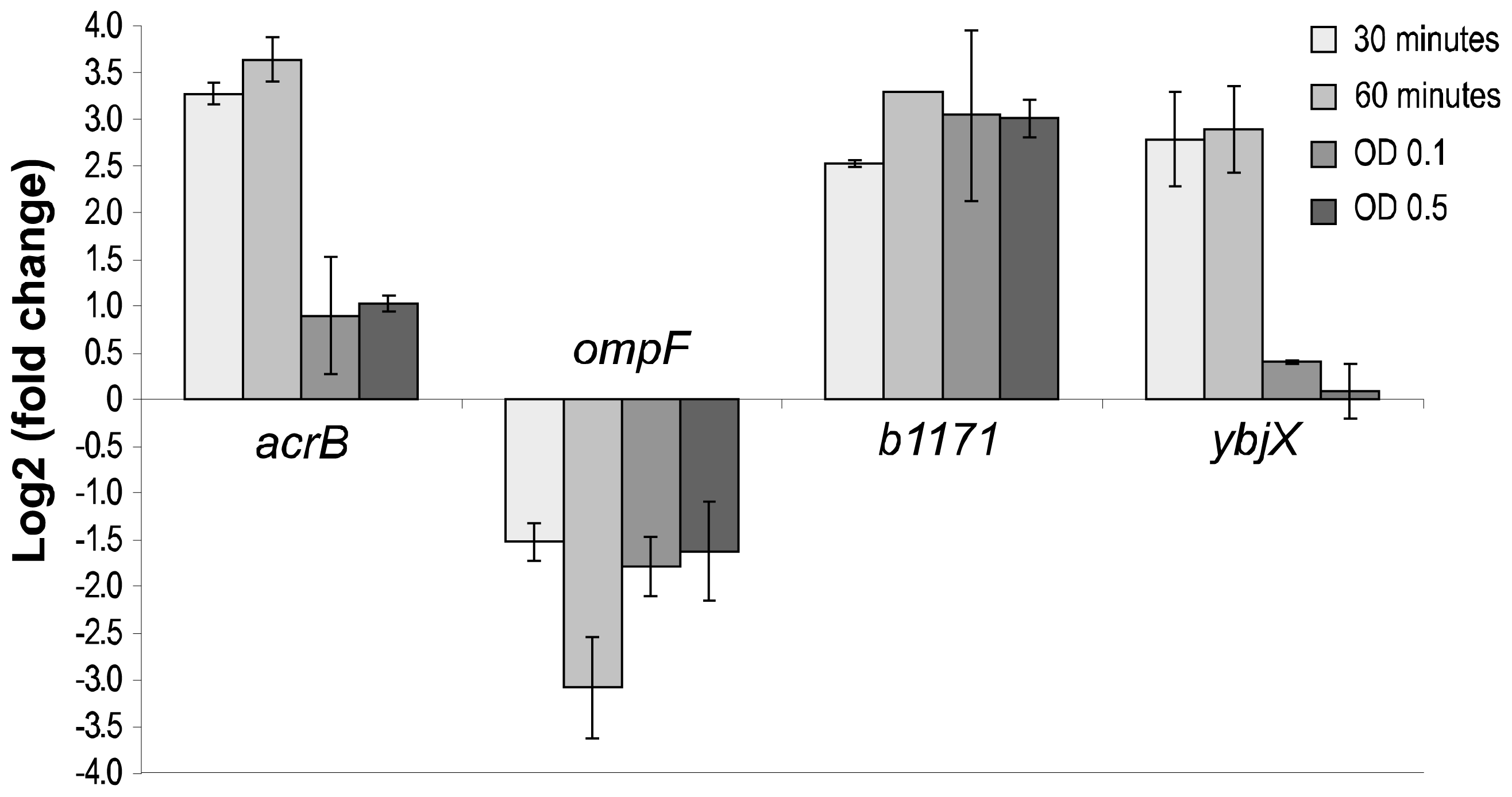
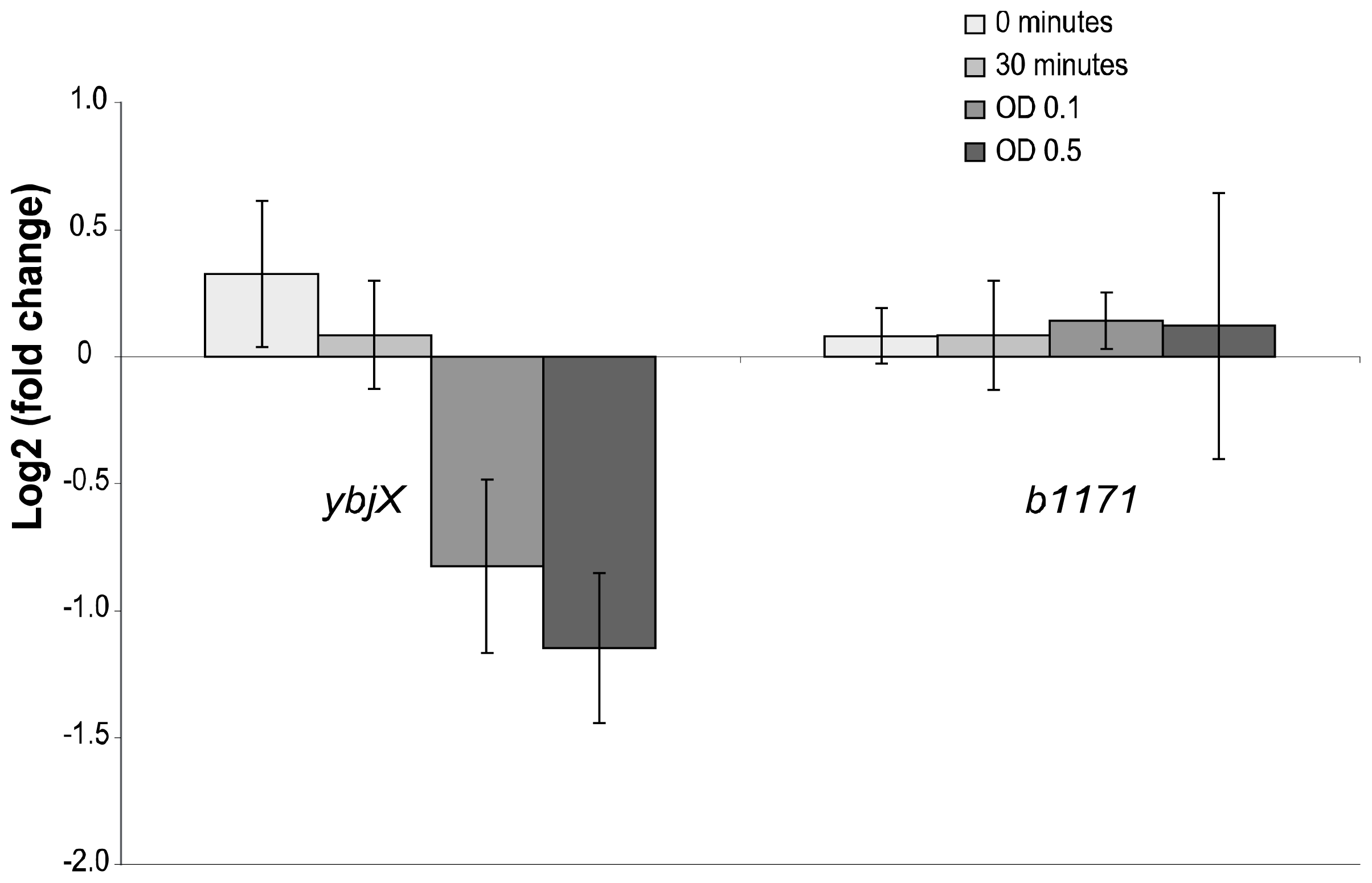
2.4. Real-Time PCR Analyses
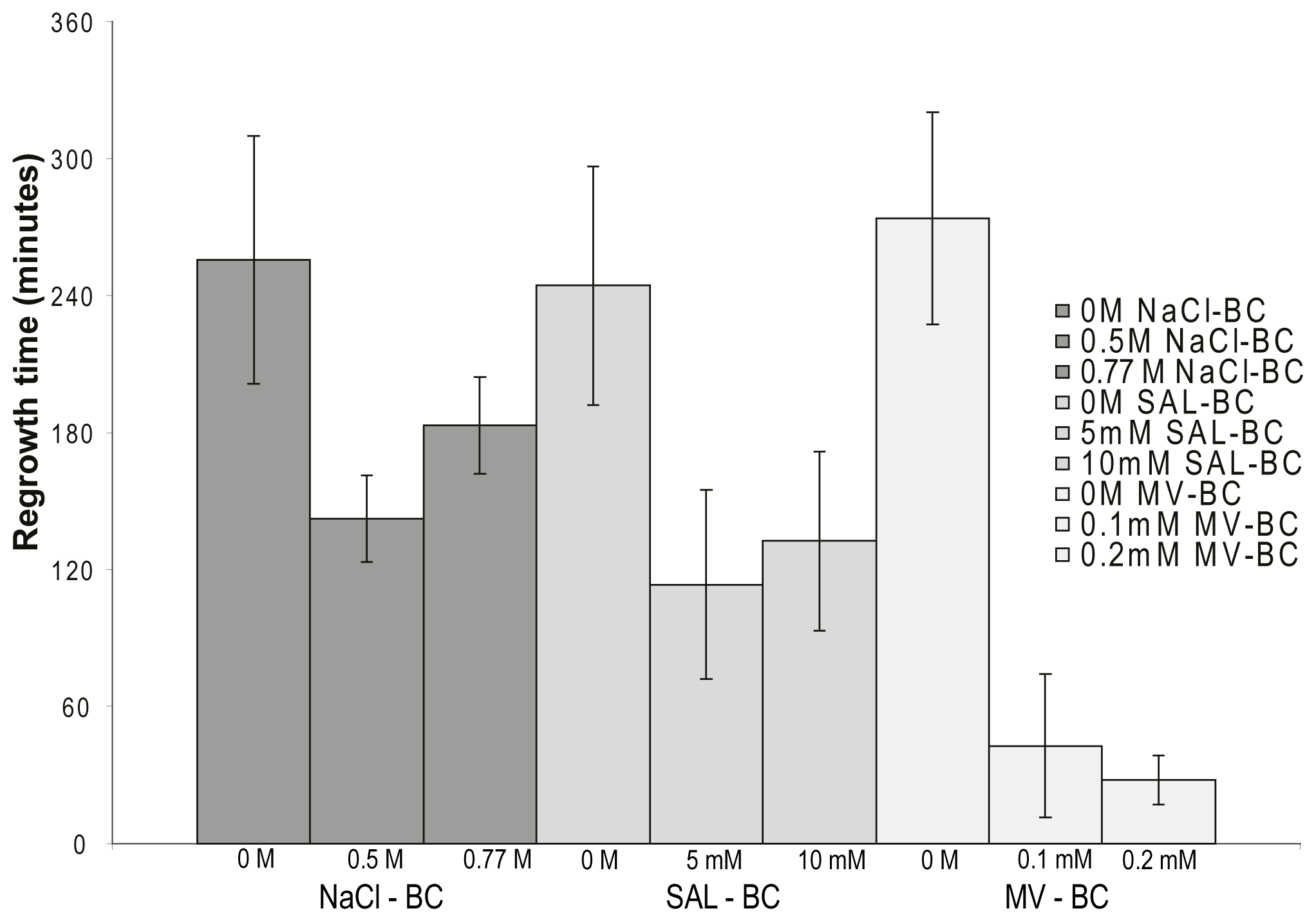
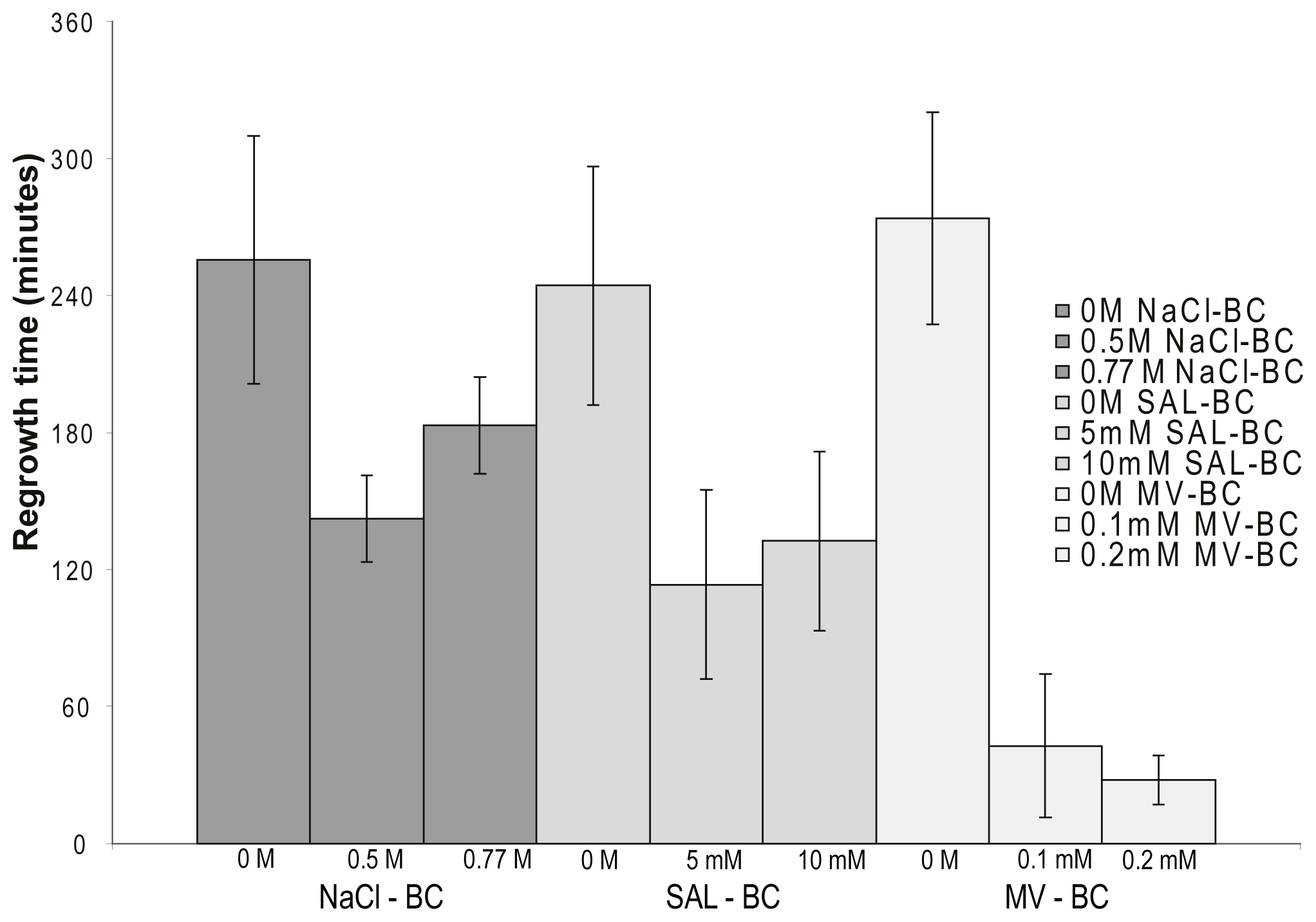
2.5. Osmotic- and Oxidative Stress
2.6. Characteristics of BC Isolates
3. Experimental Section
3.1. Strains and Growth Conditions
3.2. BacLight Live-Dead Fluorescence Microscopy
3.3. RNA Extraction
3.4. Reverse Transcription
3.5. Quantitative Real-time PCR
3.6. DNA Extraction
3.7. Knock-out Strain Analyses
3.8. Data Analysis
3.9. Pre-Exposure to Osmotic- and Oxidative Stress Experiments
3.10. Genome Sequencing (454 (Roche) Sequencing Platform)
4. Conclusions
Acknowledgments
References
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: Activity, action, and resistance. Clin. Microbiol. Rev 1999, 12, 147–179. [Google Scholar]
- Gilbert, P.; Moore, L.E. Cationic antiseptics: Diversity of action under a common epithet. J. Appl. Microbiol 2005, 99, 703–715. [Google Scholar]
- Hegstad, K.; Langsrud, S.; Lunestad, B.T.; Scheie, A.A.; Sunde, M.; Yazdankhah, S.P. Does the wide use of quaternary ammonium compounds enhance the selection and spread of antimicrobial resistance and thus threaten our health? Microb. Drug Resist 2010, 16, 91–104. [Google Scholar]
- Langsrud, S.; Sundheim, G.; Borgmann-Strahsen, R. Intrinsic and acquired resistance to quaternary ammonium compounds in food-related Pseudomonas spp. J. Appl. Microbiol 2003, 95, 874–882. [Google Scholar]
- Russell, A.D. Do biocides select for antibiotic resistance? J. Pharm. Pharmacol 2000, 52, 227–233. [Google Scholar]
- Sidhu, M.S.; Heir, E.; Sorum, H.; Holck, A. Genetic linkage between resistance to quaternary ammonium compounds and beta-lactam antibiotics in food-related Staphylococcus spp. Microb. Drug Resist 2001, 7, 363–371. [Google Scholar]
- Braoudaki, M.; Hilton, A.C. Adaptive resistance to biocides in Salmonella enterica and Escherichia coli O157 and cross-resistance to antimicrobial agents. J. Clin. Microbiol 2004, 42, 73–78. [Google Scholar]
- Braoudaki, M.; Hilton, A.C. Mechanisms of resistance in Salmonella enterica adapted to erythromycin, benzalkonium chloride and triclosan. Int. J. Antimicrob. Agents 2005, 25, 31–37. [Google Scholar]
- Langsrud, S.; Sundheim, G.; Holck, A.L. Cross-resistance to antibiotics of Escherichia coli adapted to benzalkonium chloride or exposed to stress-inducers. J. Appl. Microbiol 2004, 96, 201–208. [Google Scholar]
- Sidhu, M.S.; Sorum, H.; Holck, A. Resistance to quaternary ammonium compounds in food-related bacteria. Microb. Drug Resist 2002, 8, 393–399. [Google Scholar]
- Bore, E.; Hebraud, M.; Chafsey, I.; Chambon, C.; Skjaeret, C.; Moen, B.; Moretro, T.; Langsrud, O.; Rudi, K.; Langsrud, S. Adapted tolerance to benzalkonium chloride in Escherichia coli K-12 studied by transcriptome and proteome analyses. Microbiology 2007, 153, 935–946. [Google Scholar]
- Nishino, K.; Yamaguchi, A. Analysis of a complete library of putative drug transporter genes in Escherichia coli. J. Bacteriol 2001, 183, 5803–5812. [Google Scholar]
- Ishikawa, S.; Matsumura, Y.; Yoshizako, F.; Tsuchido, T. Characterization of a cationic surfactant-resistant mutant isolated spontaneously from Escherichia coli. J. Appl. Microbiol 2002, 92, 261–268. [Google Scholar]
- Tattawasart, U.; Maillard, J.Y.; Furr, J.R.; Russell, A.D. Outer membrane changes in Pseudomonas stutzeri resistant to chlorhexidine diacetate and cetylpyridinium chloride. Int. J. Antimicrob. Agents 2000, 16, 233–238. [Google Scholar]
- Jones, M.V.; Herd, T.M.; Christie, H.J. Resistance of Pseudomonas aeruginosa to amphoteric and quaternary ammonium biocides. Microbios 1989, 58, 49–61. [Google Scholar]
- Sakagami, Y.; Yokoyama, H.; Nishimura, H.; Ose, Y.; Tashima, T. Mechanism of resistance to benzalkonium chloride by Pseudomonas aeruginosa. Appl. Environ. Microbiol 1989, 55, 2036–2040. [Google Scholar]
- Elfalaha, B.M.A.; Russell, A.D.; Furr, J.R. Effect of chlorhexidine diacetate and benzalkonium chloride on the viability of wild-type and envelope mutants of Escherichia coli and Pseudomonas aeruginosa. Lett. Appl. Microbiol 1985, 1, 21–24. [Google Scholar]
- Moen, B.; Oust, A.; Langsrud, S.; Langsrud, O.; Hobman, J.L.; Constantinidou, C.; Kohler, A.; Rudi, K. Global responses of Escherichia coli to adverse conditions determined by microarrays and FT-IR specrtroscopy. Can. J. Microbiol 2009, 55, 714–728. [Google Scholar]
- Deckers, D.; Masschalck, B.; Aertsen, A.; Callewaert, L.; van Tiggelen, C.G.; Atanassova, M.; Michiels, C.W. Periplasmic lysozyme inhibitor contributes to lysozyme resistance in Escherichia coli. Cell Mol. Life Sci 2004, 61, 1229–1237. [Google Scholar]
- Monchois, V.; Abergel, C.; Sturgis, J.; Jeudy, S.; Claverie, J.M. Escherichia coli ykfE ORFan gene encodes a potent inhibitor of C-type lysozyme. J. Biol. Chem 2001, 276, 18437–18441. [Google Scholar]
- Dynes, J.J.; Lawrence, J.R.; Korber, D.R.; Swerhone, G.D.W.; Leppard, G.G.; Hitchcoock, A.P. Morphological and biochemical changes in Pseudomonas fluorescens biofilms induced by sub-lethal exposure to antimicrobial agents. Can. J. Microbiol 2009, 55, 163–178. [Google Scholar]
- Mc Cay, P.H.; Ocampo-Sosa, A.A.; Fleming, G.T. Effect of subinhibitory concentrations of benzalkonium chloride on the competitiveness of Pseudomonas aeruginosa grown in continuous culture. Microbiology 2010, 156, 30–38. [Google Scholar]
- During, K.; Porsch, P.; Mahn, A.; Brinkmann, O.; Gieffers, W. The non-enzymatic microbicidal activity of lysozymes. FEBS Lett 1999, 449, 93–100. [Google Scholar]
- Gerken, H.; Misra, R. Genetic evidence for functional interactions between TolC and AcrA proteins of a major antibiotic efflux pump of Escherichia coli. Mol. Microbiol 2004, 54, 620–631. [Google Scholar]
- Ma, D.; Cook, D.N.; Alberti, M.; Pon, N.G.; Nikaido, H.; Hearst, J.E. Genes acrA and acrB encode a stress-induced efflux system of Escherichia coli. Mol. Microbiol 1995, 16, 45–55. [Google Scholar]
- Gajiwala, K.S.; Burley, S.K. HDEA, a periplasmic protein that supports acid resistance in pathogenic enteric bacteria. J. Mol. Biol 2000, 295, 605–612. [Google Scholar]
- Pomposiello, P.J.; Bennik, M.H.; Demple, B. Genome-wide transcriptional profiling of the Escherichia coli responses to superoxide stress and sodium salicylate. J. Bacteriol 2001, 183, 3890–3902. [Google Scholar]
- Shaw, K.; Miller, N.; Liu, X.; Lerner, D.; Wan, J.; Bittner, A.; Morrow, B. Comparison of the changes in global gene expression of Escherichia coli induced by four bactericidal agents. J. Mol. Microbiol. Biotechnol 2003, 5, 105–122. [Google Scholar]
- Raivio, T.L. Envelope stress responses and Gram-negative bacterial pathogenesis. Mol. Microbiol 2005, 56, 1119–1128. [Google Scholar]
- Skorko-Glonek, J.; Zurawa, D.; Kuczwara, E.; Wozniak, M.; Wypych, Z.; Lipinska, B. The Escherichia coli heat shock protease HtrA participates in defense against oxidative stress. Mol. Gen. Genet 1999, 262, 342–350. [Google Scholar]
- Neidhardt, F.C. Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd ed; ASM Press: Washington, DC, USA, 1996. [Google Scholar]
- Pratt, L.A.; Hsing, W.; Gibson, K.E.; Silhavy, T.J. From acids to osmZ: Multiple factors influence synthesis of the OmpF and OmpC porins in Escherichia coli. Mol. Microbiol 1996, 20, 911–917. [Google Scholar]
- Jung, J.U.; Gutierrez, C.; Martin, F.; Ardourel, M.; Villarejo, M. Transcription of osmB, a gene encoding an Escherichia coli lipoprotein, is regulated by dual signals. Osmotic stress and stationary phase. J. Biol. Chem 1990, 265, 10574–10581. [Google Scholar]
- Boulanger, A.; Francez-Charlot, A.; Conter, A.; Castanie-Cornet, M.P.; Cam, K.; Gutierrez, C. Multistress regulation in Escherichia coli: Expression of osmB involves two independent promoters responding either to sigmaS or to the RcsCDB His-Asp phosphorelay. J. Bacteriol 2005, 187, 3282–3286. [Google Scholar]
- Storz, G.; Hengge-Aronis, R. Bacterial Stress Responses; ASM Press: Washington, DC, USA, 2000. [Google Scholar]
- Loewen, P.C.; Hu, B.; Strutinsky, J.; Sparling, R. Regulation in the rpoS regulon of Escherichia coli. Can. J. Microbiol 1998, 44, 707–717. [Google Scholar]
- Pilsl, H.; Smajs, D.; Braun, V. Characterization of colicin S4 and its receptor, OmpW, a minor protein of the Escherichia coli outer membrane. J. Bacteriol 1999, 181, 3578–3581. [Google Scholar]
- Xu, C.; Wang, S.; Ren, H.; Lin, X.; Wu, L.; Peng, X. Proteomic analysis on the expression of outer membrane proteins of Vibrio alginolyticus at different sodium concentrations. Proteomics 2005, 5, 3142–3152. [Google Scholar]
- Bochkareva, E.S.; Girshovich, A.S.; Bibi, E. Identification and characterization of the Escherichia coli stress protein UP12, a putative in vivo substrate of GroEL. Eur. J. Biochem 2002, 269, 3032–3040. [Google Scholar]
- Weber, A.; Jung, K. Profiling early osmostress-dependent gene expression in Escherichia coli using DNA macroarrays. J. Bacteriol 2002, 184, 5502–5507. [Google Scholar]
- Kim, S.Y.; Nishioka, M.; Hayashi, S.; Honda, H.; Kobayashi, T.; Taya, M. The gene yggE functions in restoring physiological defects of Escherichia coli cultivated under oxidative stress conditions. Appl. Environ. Microbiol 2005, 71, 2762–2765. [Google Scholar]
- Wagner, A.F.; Schultz, S.; Bomke, J.; Pils, T.; Lehmann, W.D.; Knappe, J. YfiD of Escherichia coli and Y06I of bacteriophage T4 as autonomous glycyl radical cofactors reconstituting the catalytic center of oxygen-fragmented pyruvate formate-lyase. Biochem. Biophys. Res. Commun 2001, 285, 456–462. [Google Scholar]
- Tong, S.; Porco, A.; Isturiz, T.; Conway, T. Cloning and molecular genetic characterization of the Escherichia coli gntR, gntK, and gntU genes of GntI, the main system for gluconate metabolism. J. Bacteriol 1996, 178, 3260–3269. [Google Scholar]
- Lehnen, D.; Blumer, C.; Polen, T.; Wackwitz, B.; Wendisch, V.F.; Unden, G. LrhA as a new transcriptional key regulator of flagella, motility and chemotaxis genes in Escherichia coli. Mol. Microbiol 2002, 45, 521–532. [Google Scholar]
- Gibson, K.E.; Silhavy, T.J. The LysR homolog LrhA promotes RpoS degradation by modulating activity of the response regulator sprE. J. Bacteriol 1999, 181, 563–571. [Google Scholar]
- EcoGene. Available online: http://bmb.med.miami.edu/EcoGene/EcoWeb/ accessed on 12 Octorber 2005.
- De Spiegeleer, P.; Sermon, J.; Vanoirbeek, K.; Aertsen, A.; Michiels, C.W. Role of porins in sensitivity of Escherichia coli to antibacterial activity of the lactoperoxidase enzyme system. Appl. Environ. Microbiol 2005, 71, 3512–3518. [Google Scholar]
- E. coli Genome Project; University of Wisconsin: Madison. Available online: http://www.genome.wisc.edu/functional/tnmutagenesis.htm accessed on 14 January 2005.
- Nikaido, H.; Zgurskaya, H.I. AcrAB and related multidrug efflux pumps of Escherichia coli. J. Mol. Microbiol. Biotechnol 2001, 3, 215–218. [Google Scholar]
- Detweiler, C.S.; Monack, D.M.; Brodsky, I.E.; Mathew, H.; Falkow, S. virK, somA and rcsC are important for systemic Salmonella enterica serovar Typhimurium infection and cationic peptide resistance. Mol. Microbiol 2003, 48, 385–400. [Google Scholar]
- Leenanon, B.; Drake, M.A. Acid stress, starvation, and cold stress affect poststress behavior of Escherichia coli O157:H7 and nonpathogenic Escherichia coli. J. Food Prot 2001, 64, 970–974. [Google Scholar]
- Mattison, K.; Kenney, L.J. Phosphorylation alters the interaction of the response regulator OmpR with its sensor kinase EnvZ. J. Biol. Chem 2002, 277, 11143–11148. [Google Scholar]
- Cohen, S.P.; McMurry, L.M.; Hooper, D.C.; Wolfson, J.S.; Levy, S.B. Cross-resistance to fluoroquinolones in multiple-antibiotic-resistant (Mar) Escherichia coli selected by tetracycline or chloramphenicol: Decreased drug accumulation associated with membrane changes in addition to OmpF reduction. Antimicrob. Agents Chemother 1989, 33, 1318–1325. [Google Scholar]
- Harder, K.J.; Nikaido, H.; Matsuhashi, M. Mutants of Escherichia coli that are resistant to certain beta-lactam compounds lack the ompF porin. Antimicrob. Agents Chemother 1981, 20, 549–552. [Google Scholar]
- Thanassi, D.G.; Suh, G.S.; Nikaido, H. Role of outer membrane barrier in efflux-mediated tetracycline resistance of Escherichia coli. J. Bacteriol 1995, 177, 998–1007. [Google Scholar]
- Lambert, P.A.; Hammond, S.M. Potassium fluxes, first indications of membrane damage in micro-organisms. Biochem. Biophys. Res. Commun 1973, 54, 796–799. [Google Scholar]
- Ceccarelli, M.; Danelon, C.; Laio, A.; Parrinello, M. Microscopic mechanism of antibiotics translocation through a porin. Biophys. J 2004, 87, 58–64. [Google Scholar]
- Goldman, R.A.; Hasan, T.; Hall, C.C.; Strycharz, W.A.; Cooperman, B.S. Photoincorporation of tetracycline into Escherichia coli Ribosomes—Identification of the major proteins photolabeled by native tetracycline and tetracycline photoproducts and implications for the inhibitory-action of tetracycline on protein-synthesis. Biochemistry 1983, 22, 359–368. [Google Scholar]
- Agrawal, R.K.; Spahn, C.M.T.; Penczek, P.; Grassucci, R.A.; Nierhaus, K.H.; Frank, J. Visualization of tRNA movements on the Escherichia coli 70S ribosome during the elongation cycle. J. Cell Biol 2000, 150, 447–459. [Google Scholar]
- Spahn, C.M.T.; Blaha, G.; Agrawal, R.K.; Penczek, P.; Grassucci, R.A.; Trieber, C.A.; Connell, S.R.; Taylor, D.E.; Nierhaus, K.H.; Frank, J. Localization of the ribosomal protection protein Tet(O) on the ribosome and the mechanism of tetracycline resistance. Mol. Cell 2001, 7, 1037–1045. [Google Scholar]
- Blattner, F.R.; Plunkett, G., III; Bloch, C.A.; Perna, N.T.; Burland, V.; Riley, M.; Collado-Vides, J.; Glasner, J.D.; Rode, C.K.; Mayhew, G.F.; et al. The complete genome sequence of Escherichia coli K-12. Science 1997, 277, 1453–1474. [Google Scholar]
- RT Color Spot Camera Software. SPOT Imaging Solutions. Available online: http://www.spotimaging.com/software/spot-camera-software-compatibility.html accessed on 29 September 2004.
- Spot Advanced Software. SPOT Imaging Solutions. Available online: http://www.spotimaging.com/software/spotadvanced.html accessed on 29 September 2004.
- Primer Express Software ABI PRISM® Software. Life Technologies. Available online: https://products.appliedbiosystems.com/ab/en/US/adirect/ab?cmd=catNavigate2&catID=605537 accessed on 3 March 2005.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function and Gene Name | Log 2 (BC/Control) | Gene Description and References |
|---|---|---|
| Efflux System | ||
| acrB | 0.9 | AcrAB-TolC efflux pump [24,25]. The acrB gene has previously been linked to BC resistance [8,12]. |
| Osmotic and/or Oxidative Stress | ||
| hdeA | 1.6 | The hdeA gene encodes the acid stress chaperone HdeA that enhances survival in extreme acid conditions [26]. The hdeA gene has also been shown to be induced by sodium salicylate [27]. |
| htrA (degP) | 0.8 | Membrane-associated serine endoprotease (known heat shock protein that has also been shown to be induced by ampicillin and kanamycin in E. coli [28] and are hypothesized to participate in the degradation of oxidatively-damaged proteins localized in the cell envelope [29,30]. |
| ompF | −1.7 | The OmpC and OmpF porins account for approx. 2% of the total protein content of the cell [31] and allow for the passive diffusion of solutes across the outer membrane. ompF has previously been shown to be regulated by changes in osmolarity [32] and in response to oxidative stress [27] and has been linked to BC resistance [13]. |
| osmB | 1.6 | osmB is a multistress-responsive gene and encodes an outer membrane lipoprotein of unknown function [33]. osmB is transcribed under the control of two independent promoters, one is responsible for the response to the growth phase and to osmotic shock, whereas the other is the target of response regulator RcsB [34] which is a positive regulatory gene for capsule (colanic acid) synthesis. |
| pflB | 1.1 | Pyruvate formate lyase I. Gene pflB has previously been shown to be induced by sodium salicylate [27]. |
| rpoS | 0.8 | The rpoS gene (σS) acts as the master regulator of stationary phase response and general stress response [35] and controls the expression of genes involved in (among others) osmotic- and oxidative stress [36]. |
| yciD (ompW) | 2.2 | Gene yciD (or ompW) encodes an outer membrane protein (OMP) and mutants that lacked the OmpW protein have been shown to be resistant to colicin S4 [37]. Studies on Vibrio alginolyticus [38] have shown the up regulation of an OmpW homolog in response to high osmolarity (NaCl) and it was hypothesized that OmpW might act as OmpC of E. coli in response to salinity stress. |
| ybdQ (uspG) | 1.1 | The ybdQ (or uspG) gene share homologies with UspA and have been shown to accumulate under various growth inhibitory conditions and induced by heat shock and may function as a universal stress protein [39]. Gene ybdQ has previously been shown to be up regulated in response to osmotic upshift in E. coli [40]. |
| yfiD | 2.0 | Glycine radical cofactor that can reactivate pyruvate formate lyase after oxidative stress. Gene yfiD has been shown to be up regulated during oxidative stress and was hypothesized to function in protecting bacterial cells from oxidative stress [41,42]. |
| Other | ||
| b2097 (fbaB) | 1.0 | Fructose 1,6-bisphosphate aldolase |
| edd | −2.1 | Phosphogluconate dehydratase |
| eutH | −0.9 | Putative integral membrane protein |
| fruB | 1.0 | Fructosephosphotransferase enzyme III |
| gntK | −3.3 | Genes gntKTU constitute the GntI system and metabolize gluconate via the Entner-Doudroff pathway [43]. gntT and gntU, encodes a high- and low- affinity gluconate transporters, respectively, and gntK, encode a thermo-resistant gluconokinase. |
| gntT | −3.1 | |
| gntU_1 | −3.1 | |
| hdhA | 0.6 | 7-alpha-hydroxysteroid dehydrogenase |
| lrhA | −1.0 | Gene lhrA encodes a LysR-type regulator LrhA [44] a new transcriptional key regulator of flagella, motility and chemotaxis genes in E. coli. LrhA is also shown to function as a regulatory component in the RpoS-dependent growth phase repression of ompF [45]. |
| malZ | 0.7 | Maltodextrin glucosidase |
| nagE | −0.6 | N-acetylglucosamine-specific enzyme II of phosphotransferase system |
| nirD | 3.5 | Nitrite reductase [NAD(P)H] subunit |
| plsB | 0.7 | Glycerolphosphate acyltransferase activity |
| rbsB | 1.5 | d-ribose binding protein, periplasmic |
| rplN | −0.4 | 50S ribosomal subunit protein L14 |
| speB | −0.6 | Agmatinase |
| tktA | 0.4 | Transketolase; binds Zn(II) |
| torS | −0.7 | Sensor kinase for torCAD operon |
| tpiA | 0.8 | Triosephosphate isomerase |
| ybbU (allR) | 0.7 | Repressor for allantoin (all) and glyoxylate (gcl) utilization operons |
| yecI (ftnB) | 1.2 | Ferritin-like protein, function unknown |
| yhiW (gadW) | 1.7 | Positive AraC-type regulator of gadA and gadBC |
| yihG | 0.6 | Characterized as poly(A) polymerase II, but this claim has been contradicted |
| ykfE (ivy) | 1.4 | The ykfE gene has previously been shown to be a strong inhibitor of C-type lysozyme and was correspondingly renamed ivy [19,20]. |
| Unknown | ||
| b1171 | 2.1 | Gene b1171 (or ymgD) has no known function and produced no hit on a Pubmed search. The protein sequence produced no close hits other than a 100% identity to a hypothetical protein in Shigella flexneri (BLAST). |
| b2107 | 1.0 | Function unknown |
| b2295 | 0.5 | Function unknown |
| yaiL | 0.8 | Function unknown |
| ybjX | 0.6 | Gene ybjX has no known function but has 99% protein homology to a putative enzyme in Shigella flexneri 2a and 55% protein homology to a putative VirK protein in Salmonella enterica (180 identical of 330 amino acids) (BLAST). |
| yieE | 1.0 | Function unknown |
| yieF | 0.8 | The yieE gene has no known function but showed 97% identity (246 identical of 253 amino acids) to a hypothetical protein from Shigella flexneri and 77% identity (194 identical of 253 amino acids) to a putative cytoplasmic protein from Salmonella typhimurium (BLAST). |
| Strain | Knock-Out Strain Details a | Source or Reference |
|---|---|---|
| MG1655 | b1171::Tn5KAN-I-SceI | University of Wisconsin, Madison b |
| MG1655 | gntK::Tn5KAN-I-SceI | University of Wisconsin, Madison b |
| MG1655 | gntT::Tn5KAN-I-SceI | University of Wisconsin, Madison b |
| MG1655 | hdeA::Tn5KAN-I-SceI | University of Wisconsin, Madison b |
| MG1655 | lrhA::Tn5KAN-2 | University of Wisconsin, Madison b |
| MG1655 | ompC::Tn5KAN | [47] |
| MG1655 | ompF::Tn5KAN | [47] |
| MG1655 | osmB::Tn5KAN-2 | University of Wisconsin, Madison b |
| MG1655 | ybjX::Tn5KAN-I-SceI | University of Wisconsin, Madison b |
| MG1655 | yciD::Tn5KAN-I-SceI | University of Wisconsin, Madison b |
| MG1655 | yieF::Tn5KAN-I-SceI | University of Wisconsin, Madison b |
| MG1655 | ivy::KAN | [19] |
| MG1655 | pAA410-arabinose-inducible Ivy overexpression strain | [19] |
| Allele Variation | Single Point Mutations (+/−) | Reference Position | Gene Annotations | Amino Acid Substitution | ||
|---|---|---|---|---|---|---|
| Control | M95 | M100 | ||||
| G→T | + | + a | + a | 720994 | Gene: kdpD | Gln→Lys |
| A→G | + a | + | + | 911614 | Gene: hcp | Leu→Pro |
| C→T | + | + a | + | 3957957 | No coding region | - |
| T→G | − b | + | − b | 962012 | Gene: rpsA | Asp→Glu |
| Gene | Probe or Primers | Sequence (5′-3′) | Denaturation Temp. (°C) |
|---|---|---|---|
| accD | probe | CGCAGTGAATTCC | 70 |
| forward primer | TGCCGCCTGGATTCCA | 59 | |
| reverse primer | GTCGATCGCGCCTTTCTC | 58 | |
| gapA | probe | CGAAACTGCTCGTAAAC | 69 |
| forward primer | AAGCAACTGGTCTGTTCCTGACT | 58 | |
| reverse primer | TTCGCACCAGCGGTGAT | 58 | |
| acrB | probe | TGCGATGGTTTTCG | 69 |
| forward primer | GCGCTTTCTCGCAAATCAA | 59 | |
| reverse primer | CGATTGCGGGCAGGTTAA | 59 | |
| ybjX | probe | CATCGCCGTAGTTTT | 68 |
| forward primer | TCGGCCGGGTAAATTCTG | 58 | |
| reverse primer | CGAGCGCAGCAAAAATTTCT | 59 | |
| b1171_ymgD | probe | CCCGCAAATGC | 68 |
| forward primer | ACTGAATCAGGTTTGCGCTAAAG | 59 | |
| reverse primer | TCATCAATTGCCGTGATCAAC | 58 | |
| ompF | probe | ACCTGGGTAAAAACGA | 68 |
| forward primer | TGGCCTGAACTTCGCTGTT | 58 | |
| reverse primer | GAACGGCGTGCAGTGTCA | 59 | |
| KAN | probe | TGCGCCGGTTGC | 69 |
| forward primer | TTGATGCGCTGGCAGTGT | 58 | |
| reverse primer | GGACAATTACAAACAGGAATCGAA | 58 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Moen, B.; Rudi, K.; Bore, E.; Langsrud, S. Subminimal Inhibitory Concentrations of the Disinfectant Benzalkonium Chloride Select for a Tolerant Subpopulation of Escherichia coli with Inheritable Characteristics. Int. J. Mol. Sci. 2012, 13, 4101-4123. https://doi.org/10.3390/ijms13044101
Moen B, Rudi K, Bore E, Langsrud S. Subminimal Inhibitory Concentrations of the Disinfectant Benzalkonium Chloride Select for a Tolerant Subpopulation of Escherichia coli with Inheritable Characteristics. International Journal of Molecular Sciences. 2012; 13(4):4101-4123. https://doi.org/10.3390/ijms13044101
Chicago/Turabian StyleMoen, Birgitte, Knut Rudi, Erlend Bore, and Solveig Langsrud. 2012. "Subminimal Inhibitory Concentrations of the Disinfectant Benzalkonium Chloride Select for a Tolerant Subpopulation of Escherichia coli with Inheritable Characteristics" International Journal of Molecular Sciences 13, no. 4: 4101-4123. https://doi.org/10.3390/ijms13044101