Design and Characterization of a Peptide Mimotope of the HIV-1 gp120 Bridging Sheet

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results and Discussion
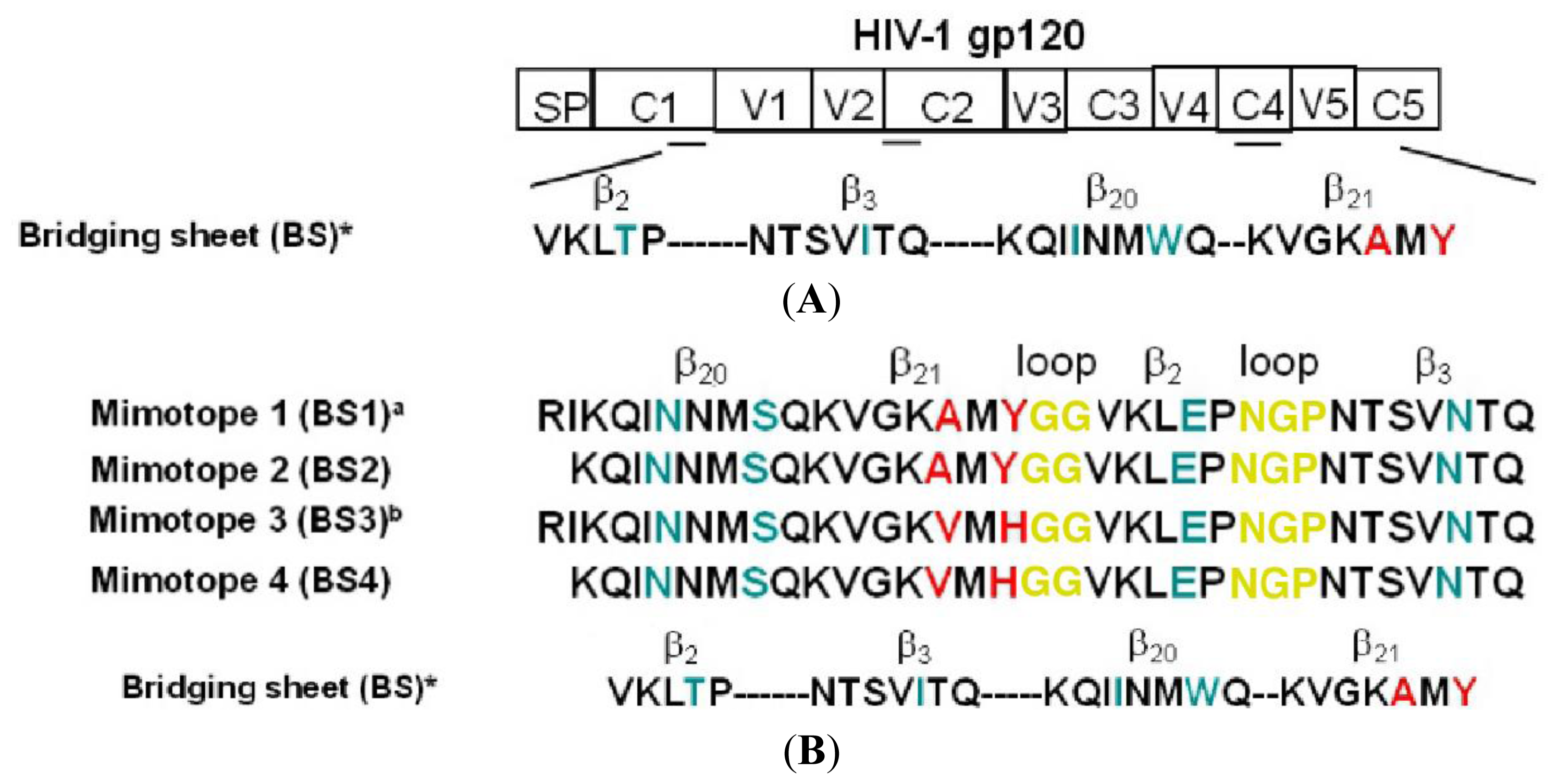
2.1. Rational Design of Bridging Sheet Peptide Mimotopes
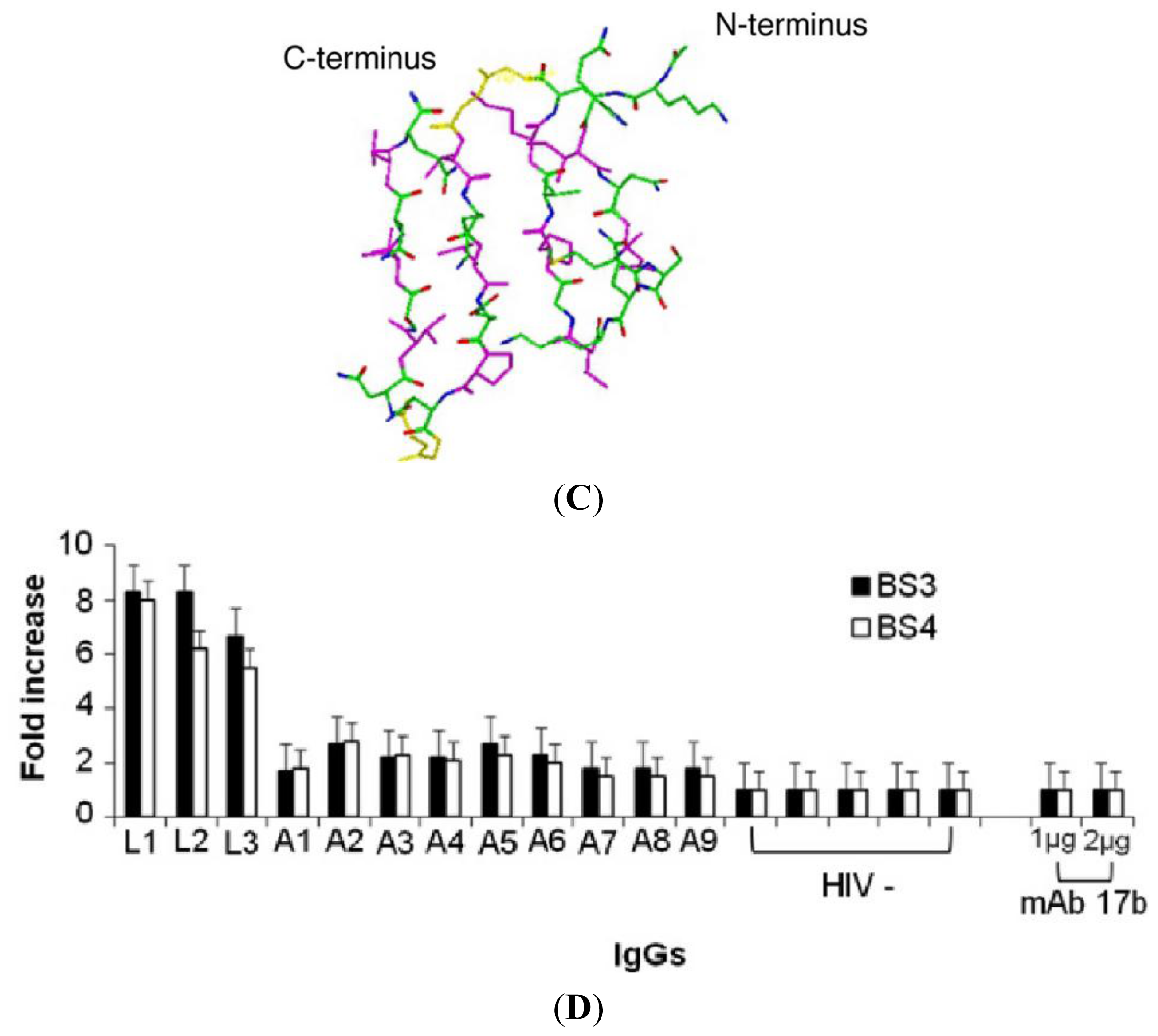
2.2. Antigenic Properties of Synthetic BS3 and BS4 Peptides
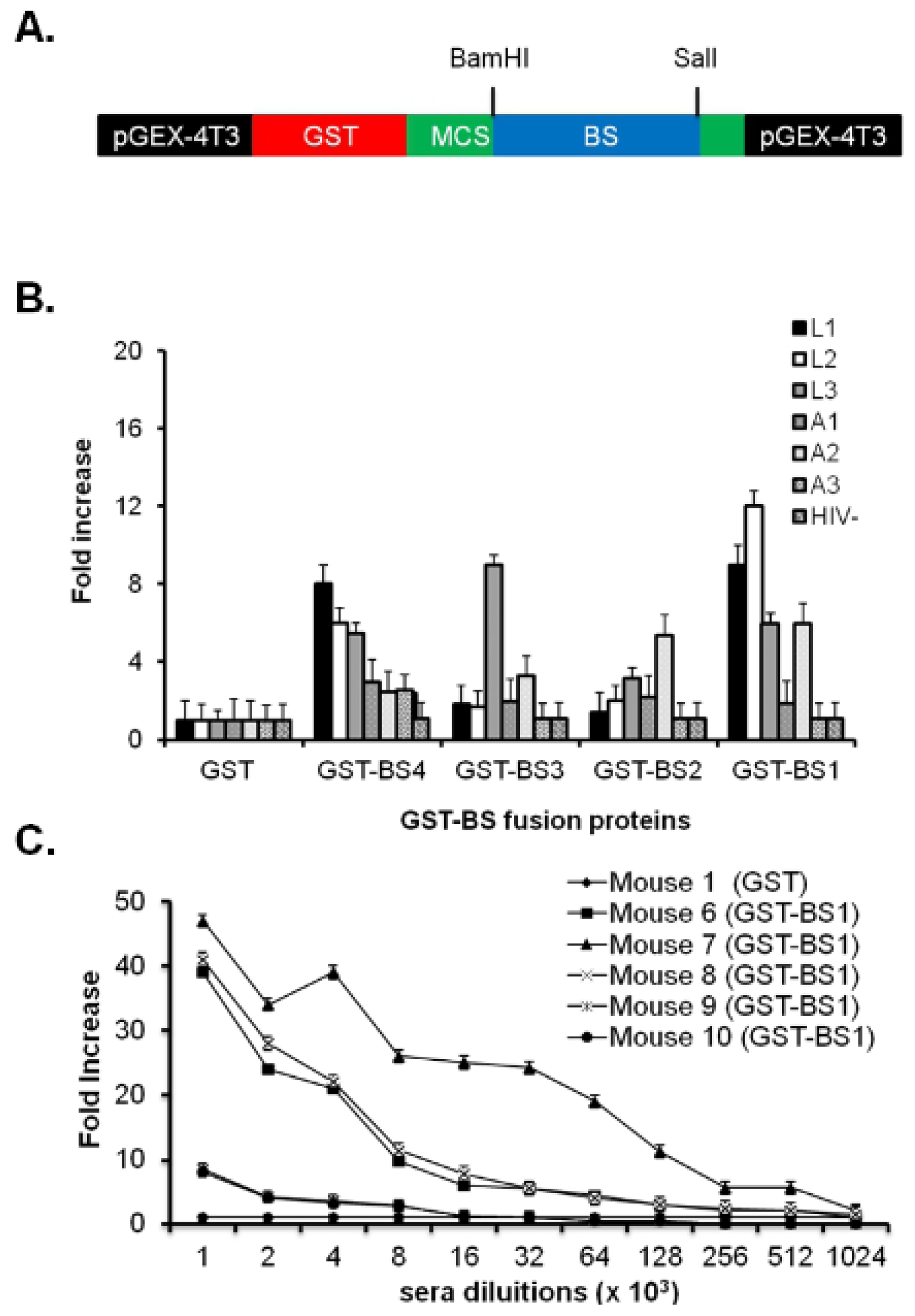
2.3. Antigenic and Immunogenic Characterization of BS Peptide Mimotopes Linked to GST Protein
2.4. E2-BS1 Sixty-Mer and Its Antigenic Characterization
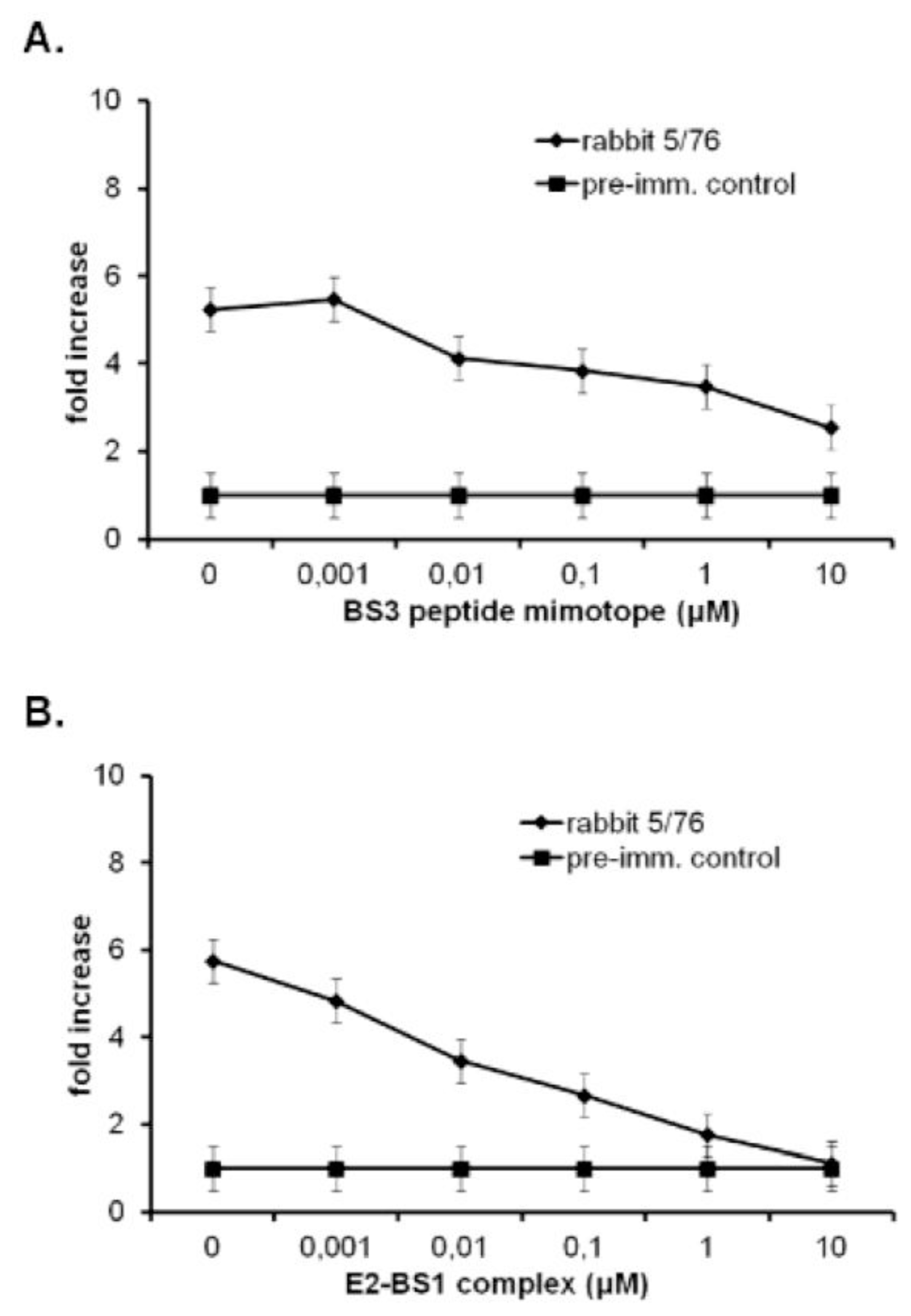
2.5. The Immune Response in Mice and Rabbits
2.6. Moderate Neutralizing Activity of Antibodies Produced in Rabbits
3. Experimental Section
3.1. Ethics Statement
3.2. Sera, Env Proteins, Mice, Rabbits
3.3. Peptide Synthesis
3.4. E2-BS1 Cloning, Expression and Purification
3.5. Electron Microscopy of E2-BS1 Complex
3.6. Immunization Schedules in Mice
3.7. Immunization Protocols in Rabbits
3.8. ELISA
3.9. Competitive ELISA Assay
3.10. Measurement of Neutralizing Response
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
- Conflict of InterestNo competing interests exist.
References
- Overbaugh, J.; Morris, L. The Antibody Response against HIV-1. Cold Spring Harb. Perspect. Med 2012, 2. [Google Scholar] [CrossRef]
- Forthal, D.N.; Gilbert, P.B.; Landucci, G.; Phan, T. Recombinant gp120 vaccine-induced antibodies inhibit clinical strains of HIV-1 in the presence of Fc receptor-bearing effector cells and correlate inversely with HIV infection rate. J. Immunol 2007, 178, 6596–6603. [Google Scholar]
- Fauci, A.S.; Johnston, M.I.; Dieffenbach, C.W.; Burton, D.R.; Hammer, S.M.; Hoxie, J.A.; Martin, M.; Overbaugh, J.; Watkins, D.I.; Mahmoud, A.; Greene, W.C. HIV vaccine research: The way forward. Science 2008, 321, 530–532. [Google Scholar]
- Mascola, J.R.; Montefiori, D.C. The role of antibodies in HIV vaccines. Ann. Rev. Immunol 2010, 28, 413–444. [Google Scholar]
- Tomaras, G.D.; Yates, N.L.; Liu, P.; Qin, L.; Fouda, G.G.; Chavez, L.L.; Decamp, A.C.; Parks, R.J.; Ashley, V.C.; Lucas, J.T.; et al. Initial B-cell responses to transmitted human immunodeficiency virus type 1: Virion-binding immunoglobulin M (IgM) and IgG antibodies followed by plasma anti-gp41 antibodies with ineffective control of initial viremia. J. Virol 2008, 82, 12449–12463. [Google Scholar]
- Stamatatos, L.; Morris, L.; Burton, D.R.; Mascola, J.R. Neutralizing antibodies generated during natural HIV-1 infection: Good news for an HIV-1 vaccine? Nat. Med 2009, 15, 866–870. [Google Scholar]
- Keele, B.F.; Giorgi, E.E.; Salazar-Gonzalez, J.F.; Decker, J.M.; Pham, K.T.; Salazar, M.G.; Sun, C.; Grayson, T.; Wang, S.; Li, H.; et al. Identification and characterization of transmitted and early founder virus envelopes in primary HIV-1 infection. Proc. Natl. Acad. Sci. USA 2008, 105, 7552–7557. [Google Scholar]
- Frost, S.D.; Liu, Y.; Pond, S.L.; Chappey, C.; Wrin, T.; Petropoulos, C.J.; Little, S.J.; Richman, D.D. Characterization of human immunodeficiency virus type 1 (HIV-1) envelope variation and neutralizing antibody responses during transmission of HIV-1 subtype B. J. Virol 2005, 79, 6523–6527. [Google Scholar]
- Deeks, S.G.; Schweighardt, B.; Wrin, T.; Galovich, J.; Hoh, R.; Sinclair, E.; Hunt, P.; McCune, J.M.; Martin, J.N.; Petropoulos, C.J.; Hecht, F.M. Neutralizing antibody responses against autologous and heterologous viruses in acute versus chronic human immunodeficiency virus (HIV) infection: Evidence for a constraint on the ability of HIV to completely evade neutralizing antibody responses. J. Virol 2006, 80, 6155–6164. [Google Scholar]
- Gray, E.S.; Moore, P.L.; Choge, I.A.; Decker, J.M.; Bibollet-Ruche, F.; Li, H.; Leseka, N.; Treurnicht, F.; Mlisana, K.; Shaw, G.M.; Karim, S.S.; Williamson, C.; Morris, L. Neutralizing antibody responses in acute human immunodeficiency virus type 1 subtype C infection. J. Virol 2007, 81, 6187–6196. [Google Scholar]
- Bunnik, E.M.; Pisas, L.; van Nuenen, A.C.; Schuitemaker, H. Autologous neutralizing humoral immunity and evolution of the viral envelope in the course of subtype B human immunodeficiency virus type 1 infection. J. Virol 2008, 82, 7932–7941. [Google Scholar]
- Zhu, C.; Matthews, T.J.; Chen, C.H. Neutralization epitopes of the HIV-1 primary isolate DH012. Vaccine 2003, 21, 3301–3306. [Google Scholar]
- Kwong, P.D.; Wyatt, R.; Robinson, J.; Sweet, R.W.; Sodroski, J.; Hendrickson, W.A. Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody. Nature 1998, 393, 648–659. [Google Scholar]
- Chan, D.C.; Fass, D.; Berger, J.M.; Kim, P.S. Core structure of gp41 from the HIV envelope glycoprotein. Cell 1997, 89, 263–273. [Google Scholar]
- Weissenhorn, W.; Dessen, A.; Harrison, S.C.; Skehel, J.J.; Wiley, D.C. Atomic structure of the ectodomain from HIV-1 gp41. Nature 1997, 387, 426–230. [Google Scholar]
- Kwong, P.D.; Wyatt, R.; Majeed, S.; Robinson, J.; Sweet, R.W.; Sodroski, J.; Hendrickson, W.A. Structures of HIV-1 gp120 envelope glycoproteins from laboratory-adapted and primary isolates. Structure 2000, 8, 1329–1339. [Google Scholar]
- Huang, C.C.; Stricher, F.; Martin, L.; Decker, J.M.; Majeed, S.; Barthe, P.; Hendrickson, W.A.; Robinson, J.; Roumestand, C.; Sodroski, J.; et al. Scorpion-toxin mimics of CD4 in complex with human immunodeficiency virus gp120 crystal structures, molecular mimicry, and neutralization breadth. Structure 2005, 13, 755–768. [Google Scholar]
- Rizzuto, C.D.; Wyatt, R.; Hernandez-Ramos, N.; Sun, Y.; Kwong, P.D.; Hendrickson, W.A.; Sodroski, J. A conserved HIV gp120 glycoprotein structure involved in chemokine receptor binding. Science 1998, 280, 1949–1953. [Google Scholar]
- Chen, B.; Vogan, E.M.; Gong, H.; Skehel, J.J.; Wiley, D.C.; Harrison, S.C. Structure of an unliganded simian immunodeficiency virus gp120 core. Nature 2005, 433, 834–841. [Google Scholar]
- Caffrey, M. Model for the structure of the HIV gp41 ectodomain: Insight into the intermolecular interactions of the gp41 loop. Biochim. Biophys. Acta 2001, 1536, 116–122. [Google Scholar]
- Wyatt, R.; Sodroski, J. The HIV-1 envelope glycoproteins: Fusogens, antigens, and immunogens. Science 1998, 280, 1884–1888. [Google Scholar]
- Eckert, D.M.; Kim, P.S. Design of potent inhibitors of HIV-1 entry from the gp41 N-peptide region. Proc. Natl. Acad. Sci. USA 2001, 98, 11187–11192. [Google Scholar]
- Wu, S.R.; Loving, R.; Lindqvist, B.; Hebert, H.; Koeck, P.J.; Sjoberg, M.; Garoff, H. Single-particle cryoelectron microscopy analysis reveals the HIV-1 spike as a tripod structure. Proc. Natl. Acad. Sci. USA 2010, 107, 18844–18849. [Google Scholar]
- Berkower, I.; Patel, C.; Ni, Y.; Virnik, K.; Xiang, Z.; Spadaccini, A. Targeted deletion in the beta20-beta21 loop of HIV envelope glycoprotein gp120 exposes the CD4 binding site for antibody binding. Virology 2008, 377, 330–338. [Google Scholar]
- Pan, Y.; Ma, B.; Keskin, O.; Nussinov, R. Characterization of the conformational state and flexibility of HIV-1 glycoprotein gp120 core domain. J. Biol. Chem 2004, 279, 30523–30530. [Google Scholar]
- Pan, Y.; Ma, B.; Nussinov, R. CD4 binding partially locks the bridging sheet in gp120 but leaves the beta2/3 strands flexible. J. Mol. Biol 2005, 350, 514–527. [Google Scholar]
- Crublet, E.; Andrieu, J.P.; Vives, R.R.; Lortat-Jacob, H. The HIV-1 envelope glycoprotein gp120 features four heparan sulfate binding domains, including the co-receptor binding site. J. Biol. Chem 2008, 283, 15193–15200. [Google Scholar]
- Rizzuto, C.; Sodroski, J. Fine definition of a conserved CCR5-binding region on the human immunodeficiency virus type 1 glycoprotein 120. AIDS Res. Hum. Retroviruses 2000, 16, 741–749. [Google Scholar]
- Zhu, C.B.; Zhu, L.; Holz-Smith, S.; Matthews, T.J.; Chen, C.H. The role of the third beta strand in gp120 conformation and neutralization sensitivity of the HIV-1 primary isolate DH012. Proc. Natl. Acad. Sci. USA 2001, 98, 15227–15232. [Google Scholar]
- Xiang, S.H.; Kwong, P.D.; Gupta, R.; Rizzuto, C.D.; Casper, D.J.; Wyatt, R.; Wang, L.; Hendrickson, W.A.; Doyle, M.L.; Sodroski, J. Mutagenic stabilization and/or disruption of a CD4-bound state reveals distinct conformations of the human immunodeficiency virus type 1 gp120 envelope glycoprotein. J. Virol 2002, 76, 9888–9899. [Google Scholar]
- Mechulam, A.; Cerutti, M.; Pugniere, M.; Misse, D.; Gajardo, J.; Roquet, F.; Robinson, J.; Veas, F. Highly conserved beta16/beta17 beta-hairpin structure in human immunodeficiency virus type 1 YU2 gp120 is critical for CCR5 binding. J. Mol. Med. (Berl. ) 2005, 83, 542–552. [Google Scholar]
- Laakso, M.M.; Lee, F.H.; Haggarty, B.; Agrawal, C.; Nolan, K.M.; Biscone, M.; Romano, J.; Jordan, A.P.; Leslie, G.J.; Meissner, E.G.; et al. V3 loop truncations in HIV-1 envelope impart resistance to coreceptor inhibitors and enhanced sensitivity to neutralizing antibodies. PLoS Pathog 2007, 3. [Google Scholar] [CrossRef]
- Liu, S.Q.; Liu, C.Q.; Fu, Y.X. Molecular motions in HIV-1 gp120 mutants reveal their preferences for different conformations. J. Mol. Graph. Model 2007, 26, 306–318. [Google Scholar]
- Nolan, K.M.; Del Prete, G.Q.; Jordan, A.P.; Haggarty, B.; Romano, J.; Leslie, G.J.; Hoxie, J.A. Characterization of a human immunodeficiency virus type 1 V3 deletion mutation that confers resistance to CCR5 inhibitors and the ability to use aplaviroc-bound receptor. J. Virol 2009, 83, 3798–3809. [Google Scholar]
- Moore, P.L.; Gray, E.S.; Morris, L. Specificity of the autologous neutralizing antibody response. Curr. Opin. HIV AIDS 2009, 4, 358–363. [Google Scholar]
- Davis, K.L.; Gray, E.S.; Moore, P.L.; Decker, J.M.; Salomon, A.; Montefiori, D.C.; Graham, B.S.; Keefer, M.C.; Pinter, A.; Morris, L.; et al. High titer HIV-1 V3-specific antibodies with broad reactivity but low neutralizing potency in acute infection and following vaccination. Virology 2009, 387, 414–426. [Google Scholar]
- Burton, D.R.; Pyati, J.; Koduri, R.; Sharp, S.J.; Thornton, G.B.; Parren, P.W.; Sawyer, L.S.; Hendry, R.M.; Dunlop, N.; Nara, P.L.; et al. Efficient neutralization of primary isolates of HIV-1 by a recombinant human monoclonal antibody. Science 1994, 266, 1024–1027. [Google Scholar]
- Binley, J.M.; Wrin, T.; Korber, B.; Zwick, M.B.; Wang, M.; Chappey, C.; Stiegler, G.; Kunert, R.; Zolla-Pazner, S.; Katinger, H.; et al. Comprehensive cross-clade neutralization analysis of a panel of anti-human immunodeficiency virus type 1 monoclonal antibodies. J. Virol 2004, 78, 13232–13252. [Google Scholar]
- Kulkarni, S.S.; Lapedes, A.; Tang, H.; Gnanakaran, S.; Daniels, M.G.; Zhang, M.; Bhattacharya, T.; Li, M.; Polonis, V.R.; McCutchan, F.E.; et al. Highly complex neutralization determinants on a monophyletic lineage of newly transmitted subtype C HIV-1 Env clones from India. Virology 2009, 385, 505–520. [Google Scholar]
- Burton, D.R.; Weiss, R.A. AIDS/HIV. A boost for HIV vaccine design. Science 2010, 329, 770–773. [Google Scholar]
- Corti, D.; Langedijk, J.P.; Hinz, A.; Seaman, M.S.; Vanzetta, F.; Fernandez-Rodriguez, B.M.; Silacci, C.; Pinna, D.; Jarrossay, D.; Balla-Jhagjhoorsingh, S.; et al. Analysis of memory B cell responses and isolation of novel monoclonal antibodies with neutralizing breadth from HIV-1-infected individuals. PloS One 2010, 5. [Google Scholar] [CrossRef]
- Moore, P.L.; Gray, E.S.; Sheward, D.; Madiga, M.; Ranchobe, N.; Lai, Z.; Honnen, W.J.; Nonyane, M.; Tumba, N.; Hermanus, T.; et al. Potent and broad neutralization of HIV-1 subtype C by plasma antibodies targeting a quaternary epitope including residues in the V2 loop. J. Virol 2011, 85, 3128–3141. [Google Scholar]
- Scheid, J.F.; Mouquet, H.; Feldhahn, N.; Seaman, M.S.; Velinzon, K.; Pietzsch, J.; Ott, R.G.; Anthony, R.M.; Zebroski, H.; Hurley, A.; et al. Broad diversity of neutralizing antibodies isolated from memory B cells in HIV-infected individuals. Nature 2009, 458, 636–640. [Google Scholar]
- Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.P.; Wang, S.K.; Ramos, A.; Chan-Hui, P.Y.; Moyle, M.; et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466–470. [Google Scholar]
- Walker, L.M.; Phogat, S.K.; Chan-Hui, P.Y.; Wagner, D.; Phung, P.; Goss, J.L.; Wrin, T.; Simek, M.D.; Fling, S.; Mitcham, J.L.; et al. Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Science 2009, 326, 285–289. [Google Scholar]
- Wu, X.; Yang, Z.Y.; Li, Y.; Hogerkorp, C.M.; Schief, W.R.; Seaman, M.S.; Zhou, T.; Schmidt, S.D.; Wu, L.; Xu, L.; et al. Rational design of envelope identifies broadly neutralizing human monoclonal antibodies to HIV-1. Science 2010, 329, 856–861. [Google Scholar]
- Johnston, M.I.; Fauci, A.S. HIV vaccine development—improving on natural immunity. N. Engl. J. Med 2011, 365, 873–875. [Google Scholar]
- Scheid, J.F.; Mouquet, H.; Ueberheide, B.; Diskin, R.; Klein, F.; Oliveira, T.Y.; Pietzsch, J.; Fenyo, D.; Abadir, A.; Velinzon, K.; et al. Sequence and structural convergence of broad and potent HIV antibodies that mimic CD4 binding. Science 2011, 333, 1633–1637. [Google Scholar]
- Wu, X.; Zhou, T.; Zhu, J.; Zhang, B.; Georgiev, I.; Wang, C.; Chen, X.; Longo, N.S.; Louder, M.; McKee, K.; et al. Focused evolution of HIV-1 neutralizing antibodies revealed by structures and deep sequencing. Science 2011, 333, 1593–1602. [Google Scholar]
- Zhou, T.; Georgiev, I.; Wu, X.; Yang, Z.Y.; Dai, K.; Finzi, A.; Kwon, Y.D.; Scheid, J.F.; Shi, W.; Xu, L.; et al. Structural basis for broad and potent neutralization of HIV-1 by antibody VRC01. Science 2010, 329, 811–817. [Google Scholar]
- Lavine, C.L.; Lao, S.; Montefiori, D.C.; Haynes, B.F.; Sodroski, J.G.; Yang, X. High-mannose glycan-dependent epitopes are frequently targeted in broad neutralizing antibody responses during human immunodeficiency virus type 1 infection. J. Virol 2012, 86, 2153–2164. [Google Scholar]
- Lim, K.; Ho, J.X.; Keeling, K.; Gilliland, G.L.; Ji, X.; Ruker, F.; Carter, D.C. Three-dimensional structure of Schistosoma japonicum glutathione S-transferase fused with a six-amino acid conserved neutralizing epitope of gp41 from HIV. Protein Sci 1994, 3, 2233–2244. [Google Scholar]
- Jaworski, J.P.; Krebs, S.J.; Trovato, M.; Kovarik, D.N.; Brower, Z.; Sutton, W.F.; Waagmeester, G.; Sartorius, R.; D’Apice, L.; Caivano, A.; et al. Co-immunization with multimeric scaffolds and DNA rapidly induces potent autologous HIV-1 neutralizing antibodies and CD8+ T cells. PloS One 2012, 7. [Google Scholar] [CrossRef]
- de Berardinis, P.; Haigwood, N.L. New recombinant vaccines based on the use of prokaryotic antigen-display systems. Expert Rev. Vaccines 2004, 3, 673–679. [Google Scholar]
- Domingo, G.J.; Orru, S.; Perham, R.N. Multiple display of peptides and proteins on a macromolecular scaffold derived from a multienzyme complex. J. Mol. Biol 2001, 305, 259–267. [Google Scholar]
- Allen, M.D.; Broadhurst, R.W.; Solomon, R.G.; Perham, R.N. Interaction of the E2 and E3 components of the pyruvate dehydrogenase multienzyme complex of Bacillus stearothermophilus. Use of a truncated protein domain in NMR spectroscopy. FEBS J 2005, 272, 259–268. [Google Scholar]
- Liu, S.Q.; Liu, S.X.; Fu, Y.X. Molecular motions of human HIV-1 gp120 envelope glycoproteins. J. Mol. Model 2008, 14, 857–870. [Google Scholar]
- Wang, J.; Sen, J.; Rong, L.; Caffrey, M. Role of the HIV gp120 conserved domain 1 in processing and viral entry. J. Biol. Chem 2008, 283, 32644–32649. [Google Scholar]
- Wang, Q.; Shang, H.; Han, X.; Zhang, Z.; Jiang, Y.; Wang, Y.; Dai, D.; Diao, Y. High level serum neutralizing antibody against HIV-1 in Chinese long-term non-progressors. Microbiol. Immunol 2008, 52, 209–215. [Google Scholar]
- Humbert, M.; Antoni, S.; Brill, B.; Landersz, M.; Rodes, B.; Soriano, V.; Wintergerst, U.; Knechten, H.; Staszewski, S.; von Laer, D.; Dittmar, M.T.; Dietrich, U. Mimotopes selected with antibodies from HIV-1-neutralizing long-term non-progressor plasma. Eur. J. Immunol 2007, 37, 501–515. [Google Scholar]
- Hoxie, J.A. Toward an antibody-based HIV-1 vaccine. Ann. Rev. Med 2010, 61, 135–152. [Google Scholar]
- Puca, A.; Fiume, G.; Palmieri, C.; Trimboli, F.; Olimpico, F.; Scala, G.; Quinto, I. IkappaB-alpha represses the transcriptional activity of the HIV-1 Tat transactivator by promoting its nuclear export. J. Biol. Chem 2007, 282, 37146–37157. [Google Scholar]
- Fiume, G.; Vecchio, E.; de Laurentiis, A.; Trimboli, F.; Palmieri, C.; Pisano, A.; Falcone, C.; Pontoriero, M.; Rossi, A.; Scialdone, A.; et al. Human immunodeficiency virus-1 Tat activates NF-kappaB via physical interaction with IkappaB-alpha and p65. Nucleic Acids Res 2012, 40, 3548–3562. [Google Scholar]
- Montefiori, D.C. Evaluating Neutralizing Antibodies Against HIV, SIV, and SHIV in Luciferase Reporter Gene Assays. In Current Protocols in Immunology; Wiley: Hoboken, NJ, USA, 2005; Volume Chapter 12. [Google Scholar]
- Fenyo, E.M.; Heath, A.; Dispinseri, S.; Holmes, H.; Lusso, P.; Zolla-Pazner, S.; Donners, H.; Heyndrickx, L.; Alcami, J.; Bongertz, V.; et al. International network for comparison of HIV neutralization assays: The NeutNet report. PloS One 2009, 4. [Google Scholar] [CrossRef]
- Montefiori, D.; Sattentau, Q.; Flores, J.; Esparza, J.; Mascola, J. Antibody-based HIV-1 vaccines: Recent developments and future directions. PLoS Med 2007, 4. [Google Scholar] [CrossRef]
- Seaman, M.S.; Leblanc, D.F.; Grandpre, L.E.; Bartman, M.T.; Montefiori, D.C.; Letvin, N.L.; Mascola, J.R. Standardized assessment of NAb responses elicited in rhesus monkeys immunized with single- or multi-clade HIV-1 envelope immunogens. Virology 2007, 367, 175–186. [Google Scholar]
- Vitagliano, L.; Fiume, G.; Scognamiglio, P.L.; Doti, N.; Cannavo, R.; Puca, A.; Pedone, C.; Scala, G.; Quinto, I.; Marasco, D. Structural and functional insights into IkappaB-alpha/HIV-1 Tat interaction. Biochimie 2011, 93, 1592–1600. [Google Scholar]
- Schiavone, M.; Quinto, I.; Scala, G. Perspectives for a protective HIV-1 vaccine. Adv. Pharmacol 2008, 56, 423–452. [Google Scholar]
- Chavan, M. AIDS vaccine fails. Natl. Med. J. India 2007, 20, 273. [Google Scholar]
- Qiu, C.; Xu, J.Q. HIV-1/AIDS vaccine development: Are we in the darkness. Chin. Med. J 2008, 121, 939–945. [Google Scholar]
- Walker, L.M.; Burton, D.R. Rational antibody-based HIV-1 vaccine design: Current approaches and future directions. Curr. Opin. Immunol 2010, 22, 358–366. [Google Scholar]
- Kwong, P.D.; Wilson, I.A. HIV-1 and influenza antibodies: Seeing antigens in new ways. Nat. Immunol 2009, 10, 573–578. [Google Scholar]
- Burton, D.R.; Desrosiers, R.C.; Doms, R.W.; Koff, W.C.; Kwong, P.D.; Moore, J.P.; Nabel, G.J.; Sodroski, J.; Wilson, I.A.; Wyatt, R.T. HIV vaccine design and the neutralizing antibody problem. Nat. Immunol 2004, 5, 233–236. [Google Scholar]
- Dormitzer, P.R.; Ulmer, J.B.; Rappuoli, R. Structure-based antigen design: A strategy for next generation vaccines. Trends Biotechnol 2008, 26, 659–667. [Google Scholar]
- Huang, Y.; Krasnitz, M.; Rabadan, R.; Witten, D.M.; Song, Y.; Levine, A.J.; Ho, D.D.; Robins, H. A recoding method to improve the humoral immune response to an HIV DNA vaccine. PloS One 2008, 3. [Google Scholar] [CrossRef]
- Hsu, S.T.; Bonvin, A.M. Atomic insight into the CD4 binding-induced conformational changes in HIV-1 gp120. Proteins 2004, 55, 582–593. [Google Scholar]
- Biscone, M.J.; Miamidian, J.L.; Muchiri, J.M.; Baik, S.S.; Lee, F.H.; Doms, R.W.; Reeves, J.D. Functional impact of HIV coreceptor-binding site mutations. Virology 2006, 351, 226–236. [Google Scholar]
- D’Apice, L.; Sartorius, R.; Caivano, A.; Mascolo, D.; Del Pozzo, G.; Di Mase, D.S.; Ricca, E.; Li Pira, G.; Manca, F.; Malanga, D.; de Palma, R.; de Berardinis, P. Comparative analysis of new innovative vaccine formulations based on the use of procaryotic display systems. Vaccine 2007, 25, 1993–2000. [Google Scholar]
- de Berardinis, P.; Sartorius, R.; Caivano, A.; Mascolo, D.; Domingo, G.J.; del Pozzo, G.; Gaubin, M.; Perham, R.N.; Piatier-Tonneau, D.; Guardiola, J. Use of fusion proteins and procaryotic display systems for delivery of HIV-1 antigens: Development of novel vaccines for HIV-1 infection. Curr. HIV Res 2003, 1, 441–446. [Google Scholar]
- Zwick, M.B.; Komori, H.K.; Stanfield, R.L.; Church, S.; Wang, M.; Parren, P.W.; Kunert, R.; Katinger, H.; Wilson, I.A.; Burton, D.R. The long third complementarity-determining region of the heavy chain is important in the activity of the broadly neutralizing anti-human immunodeficiency virus type 1 antibody 2F5. J. Virol 2004, 78, 3155–3161. [Google Scholar]
- Nelson, J.D.; Kinkead, H.; Brunel, F.M.; Leaman, D.; Jensen, R.; Louis, J.M.; Maruyama, T.; Bewley, C.A.; Bowdish, K.; Clore, G.M.; et al. Antibody elicited against the gp41 N-heptad repeat (NHR) coiled-coil can neutralize HIV-1 with modest potency but non-neutralizing antibodies also bind to NHR mimetics. Virology 2008, 377, 170–183. [Google Scholar]
- Huang, Y.; Chen, A.; Li, X.; Chen, Z.; Zhang, W.; Song, Y.; Gurner, D.; Gardiner, D.; Basu, S.; Ho, D.D.; Tsuji, M. Enhancement of HIV DNA vaccine immunogenicity by the NKT cell ligand, alpha-galactosylceramide. Vaccine 2008, 26, 1807–1816. [Google Scholar]
- Vcelar, B.; Stiegler, G.; Wolf, H.M.; Muntean, W.; Leschnik, B.; Mehandru, S.; Markowitz, M.; Armbruster, C.; Kunert, R.; Eibl, M.M.; Katinger, H. Reassessment of autoreactivity of the broadly neutralizing HIV antibodies 4E10 and 2F5 and retrospective analysis of clinical safety data. AIDS 2007, 21, 2161–2170. [Google Scholar]
- Haynes, B.F.; Fleming, J.; St Clair, E.W.; Katinger, H.; Stiegler, G.; Kunert, R.; Robinson, J.; Scearce, R.M.; Plonk, K.; Staats, H.F.; et al. Cardiolipin polyspecific autoreactivity in two broadly neutralizing HIV-1 antibodies. Science 2005, 308, 1906–1908. [Google Scholar]
- Matyas, G.R.; Beck, Z.; Karasavvas, N.; Alving, C.R. Lipid binding properties of 4E10, 2F5, and WR304 monoclonal antibodies that neutralize HIV-1. Biochim. Biophys. Acta 2009, 1788, 660–665. [Google Scholar]
- Martinez, V.; Diemert, M.C.; Braibant, M.; Potard, V.; Charuel, J.L.; Barin, F.; Costagliola, D.; Caumes, E.; Clauvel, J.P.; Autran, B.; Musset, L. Anticardiolipin antibodies in HIV infection are independently associated with antibodies to the membrane proximal external region of gp41 and with cell-associated HIV DNA and immune activation. Clin. Infect. Dis 2009, 48, 123–132. [Google Scholar]
- Chakraborty, K.; Shivakumar, P.; Raghothama, S.; Varadarajan, R. NMR structural analysis of a peptide mimic of the bridging sheet of HIV-1 gp120 in methanol and water. Biochem. J 2005, 390, 573–581. [Google Scholar]
- Hessell, A.J.; Poignard, P.; Hunter, M.; Hangartner, L.; Tehrani, D.M.; Bleeker, W.K.; Parren, P.W.; Marx, P.A.; Burton, D.R. Effective, low-titer antibody protection against low-dose repeated mucosal SHIV challenge in macaques. Nat. Med 2009, 15, 951–954. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ELISA Antigen Titer after Four iImmunizations * | |||||
|---|---|---|---|---|---|
| Animal a | Immunogen | Bridging Sheet | gp120-SF162 | gp140DV2-SF162 | E2 Protein |
| Mouse1 | E2-BS1 | 100,000 | <20 | -- | 500,000 |
| Mouse2 | E2-BS1 | 100,000 | <20 | -- | 1,000,000 |
| Mouse3 | E2-BS1 | 50,000 | <20 | -- | 500,000 |
| Mouse4 | E2 | <20 | <20 | -- | 1,000,000 |
| Mouse5 | E2 | <20 | <20 | -- | 1,000,000 |
| Mouse6 | E2 | <20 | <20 | -- | 500,000 |
| Rabbit1 (#E2-BS60 male) | E2-BS1 | 500,000 | 400 b | -- | 1,000,000 |
| Rabbit2 (#E2-BS60 female) | E2-BS1 | 2000 | 200 b | -- | 500,000 |
| Rabbit3 (#E2-BS male) | E2-BS1 | 500,000 | 400 c | -- | 1,000,000 |
| Rabbit4 (#E2-BS female) | E2-BS1 | 150,000 | 20,000 c | -- | 1,000,000 |
| Rabbit5 (#5/76) | E2-BS1 | 100,000 | 5000 c | 3000 c | 500,000 |
| Rabbit6 (#6/76) | E2-BS1 | 100 | <20 | <20 | 75,000 |
| Rabbit7 (#1/76) | E2 | <20 | <20 | <20 | 500,000 |
| Rabbit8 (#3/76) | E2 | <20 | <20 | <20 | 1,000,000 |
| Rabbit9 (#E2-wt female) | E2 | <20 | <20 | -- | 1,000,000 |
| Rabbit10 (#E2-wt male) | E2 | <20 | <20 | -- | 500,000 |
| A | ||||
|---|---|---|---|---|
| ID50 in TZM-bl Cells 1 | ||||
| Animal (rabbits) | Bleed | SF162.LS | HIV-0012466-2.52 | NL-ADArs |
| First set of assays | ||||
| Rabbit 5 | post-immune | 114 | 30 | 64 |
| Rabbit 8 | post-immune | 27 | 28 | 22 |
| Second set of assays | ||||
| Rabbit 5 | post-immune | 109 | 53 | 155 |
| Rabbit 8 | post-immune | <20 | 30 | <20 |
| naive rabbit pool | Naïve | <20 | <20 | <20 |
| B | ||||
| ID50 in TZM-bl Cells 1 | ||||
| Animal (Rabbits) | Bleed | HIV-1 MN | SF162.LS | SIVmac239CS.23 |
| Rabbit 9 | Post-immune | <20 | <20 | <20 |
| Rabbit 10 | Post-immune | <20 | <20 | <20 |
| Rabbit 1 | Post-immune | <20 | <20 | <20 |
| Rabbit 2 | Post-immune | <20 | <20 | <20 |
| Rabbit 3 | Post-immune | <20 | <20 | <20 |
| Rabbit 4 | Post-immune | <20 | <20 | <20 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Schiavone, M.; Fiume, G.; Caivano, A.; De Laurentiis, A.; Falcone, C.; Masci, F.F.; Iaccino, E.; Mimmi, S.; Palmieri, C.; Pisano, A.; et al. Design and Characterization of a Peptide Mimotope of the HIV-1 gp120 Bridging Sheet. Int. J. Mol. Sci. 2012, 13, 5674-5699. https://doi.org/10.3390/ijms13055674
Schiavone M, Fiume G, Caivano A, De Laurentiis A, Falcone C, Masci FF, Iaccino E, Mimmi S, Palmieri C, Pisano A, et al. Design and Characterization of a Peptide Mimotope of the HIV-1 gp120 Bridging Sheet. International Journal of Molecular Sciences. 2012; 13(5):5674-5699. https://doi.org/10.3390/ijms13055674
Chicago/Turabian StyleSchiavone, Marco, Giuseppe Fiume, Antonella Caivano, Annamaria De Laurentiis, Cristina Falcone, Francesca Fasanella Masci, Enrico Iaccino, Selena Mimmi, Camillo Palmieri, Antonio Pisano, and et al. 2012. "Design and Characterization of a Peptide Mimotope of the HIV-1 gp120 Bridging Sheet" International Journal of Molecular Sciences 13, no. 5: 5674-5699. https://doi.org/10.3390/ijms13055674