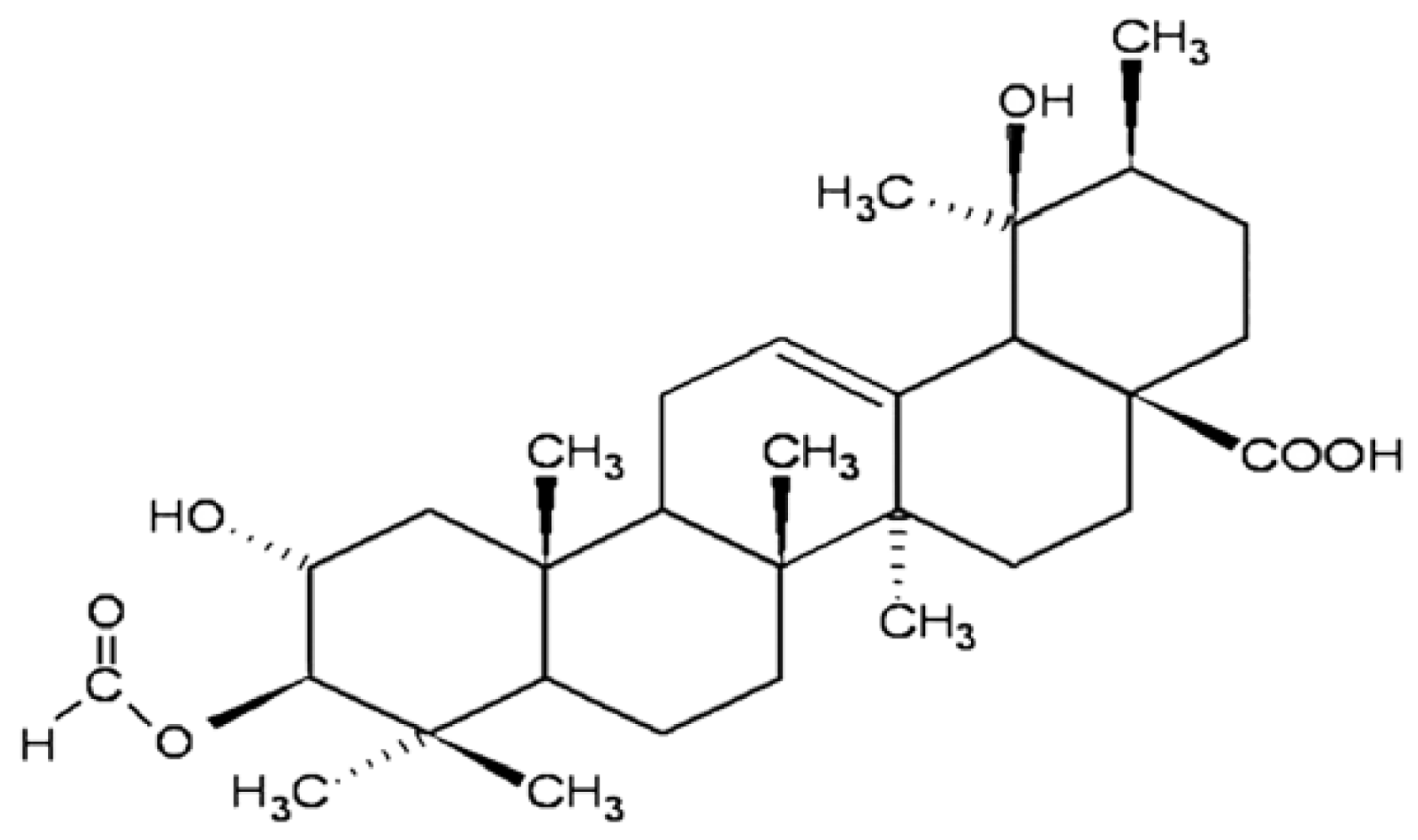
Effects of 3β-Acethyl Tormentic Acid (3ATA) on ABCC Proteins Activity

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
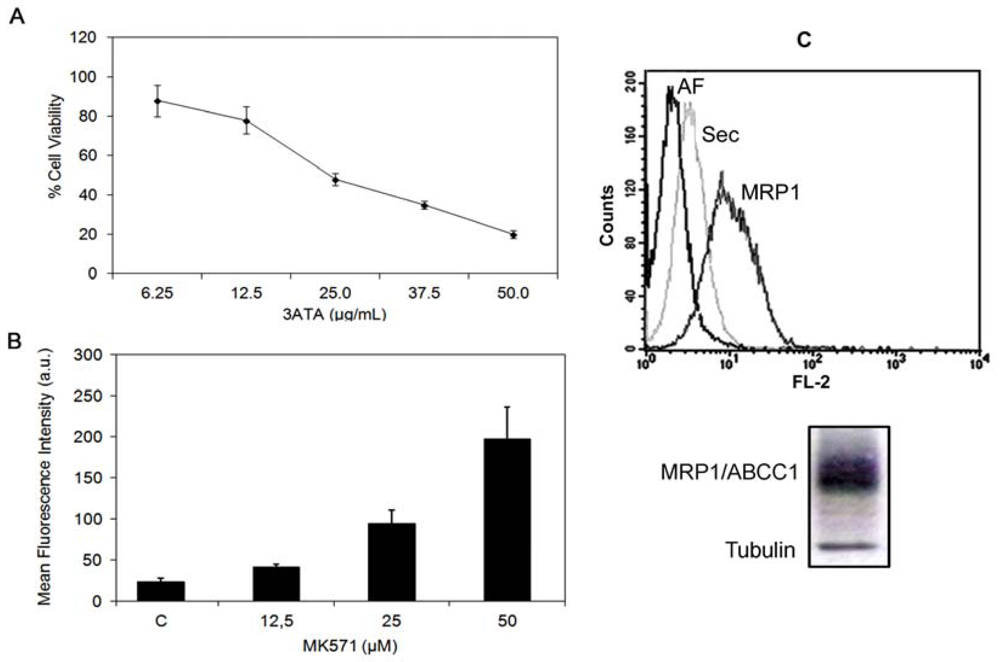
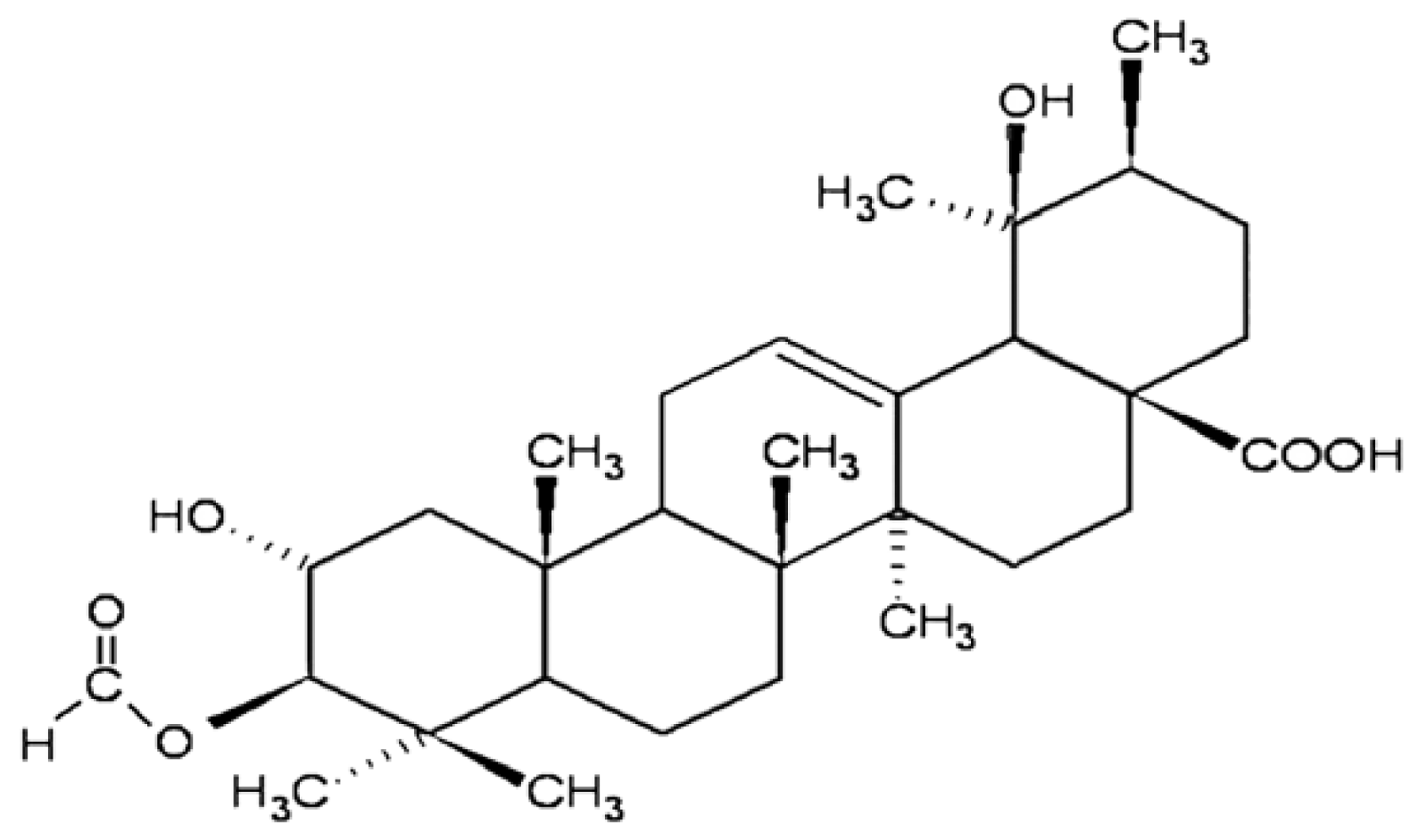
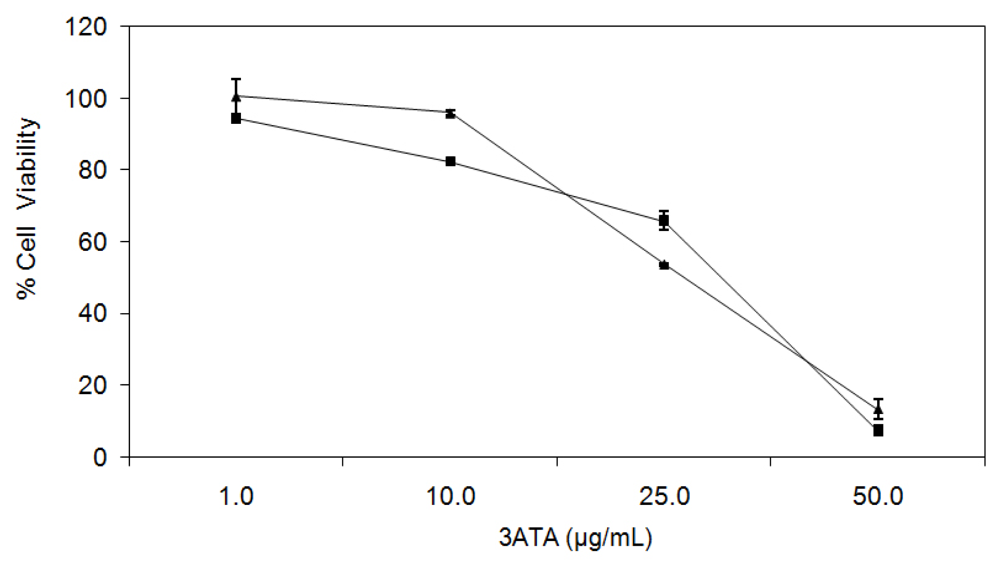
2.1.1. Cytotoxicity of 3ATA to MRP-1/ABCC1-Expressing Cell Lines
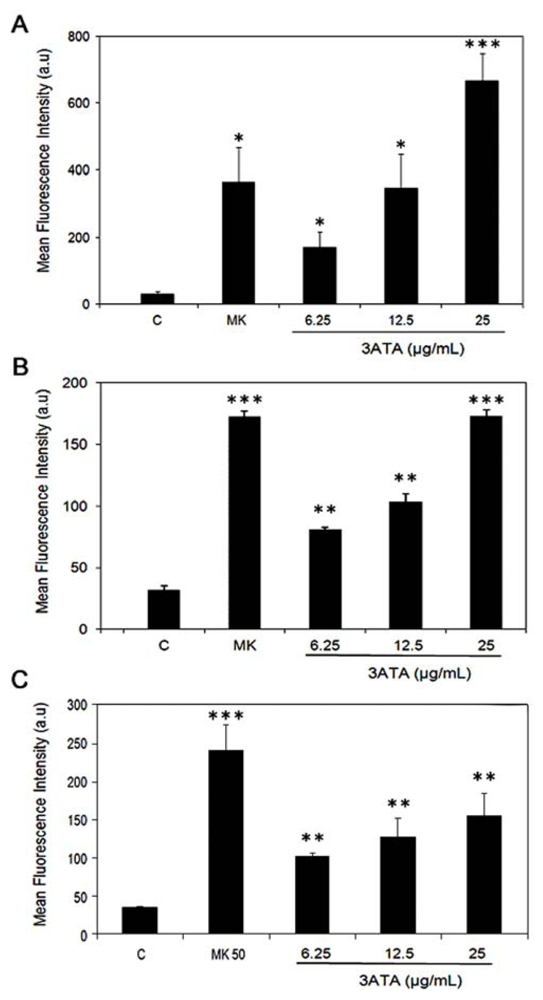
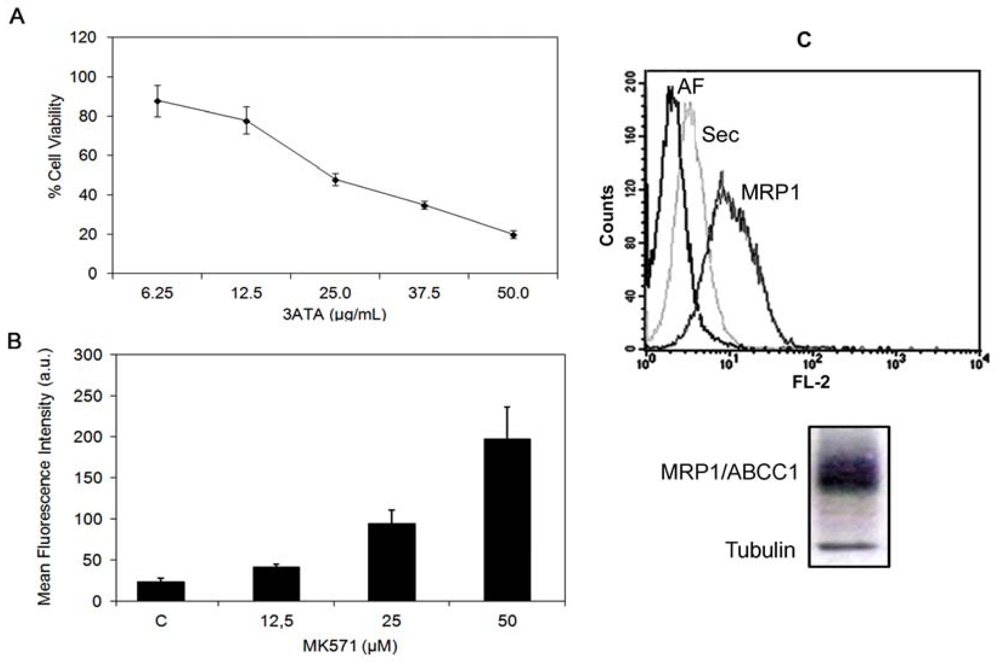
2.1.2. Modulation of MRP1/ABCC1 Activity by 3ATA
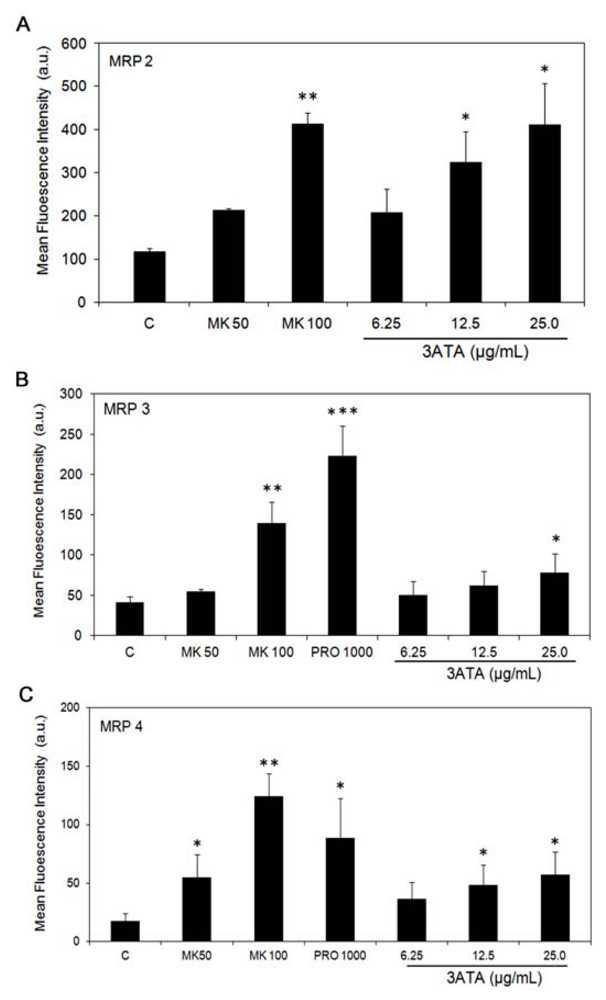
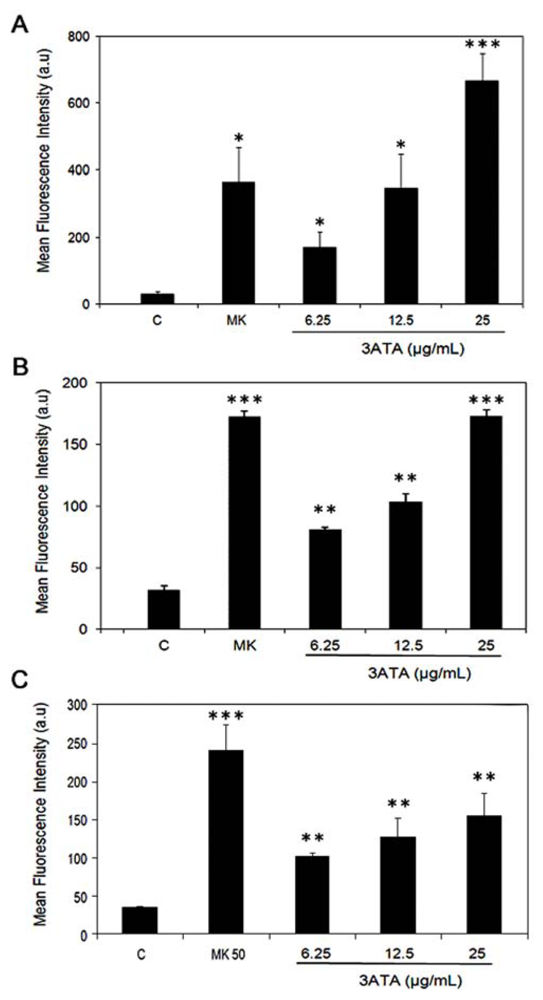
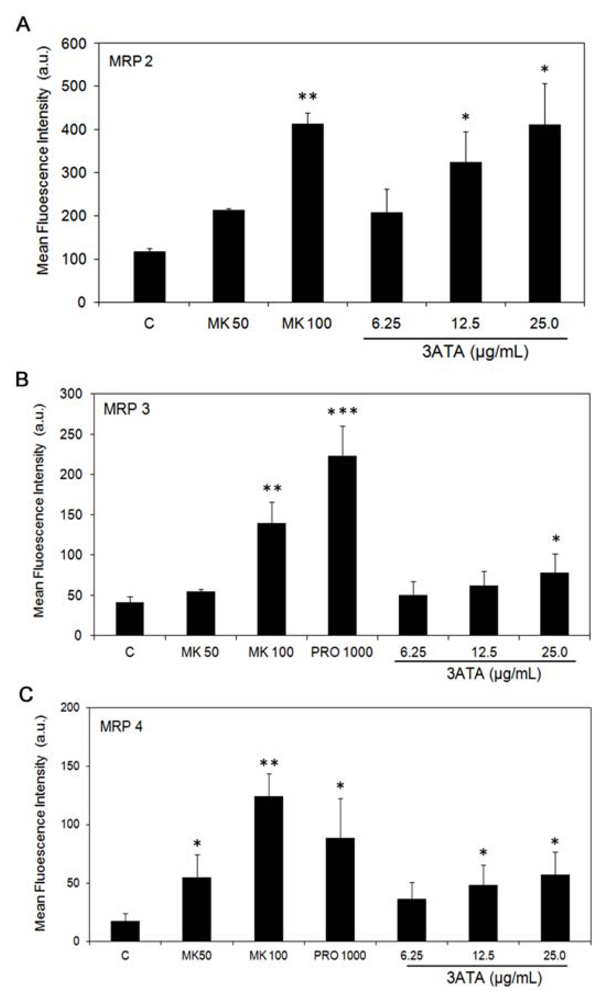
2.1.3. Effects of 3ATA on the Activity of Other MRP/ABCC Transporter Proteins
2.2. Discussion
3. Experimental Section
3.1. Chemicals and Cell Culture
3.2. MTT Assay
3.3. Activity and Expression of MRP/ABCC Proteins
3.4. Statistical Analysis
4. Conclusion
Acknowledgments
References
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP-dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar]
- Chen, Z.S.; Tiwari, A.K. Multidrug resistance proteins (MRPs/ABCCs) in cancer chemotherapy and genetic diseases. FEBS J 2011, 278, 3226–3245. [Google Scholar]
- Sharon, F.J. ABC Multidrug transporters: Structure, function and role in chemoresistance. Pharmacogenomics 2008, 9, 106–127. [Google Scholar]
- Keppler, D. Multidrug Resistance Proteins (MRPs, ABCCs): Importance for Pathophysiology and Drug Therapy. In Drug Transporters, Handbook of Experimental Pharmacology; From, M.F., Kim, R.B., Eds.; Springer-Verlag: Berlin, Germany, 2011; Volume 201, pp. 299–323. [Google Scholar]
- Slot, A.J.; Molinski, S.V.; Cole, S.P.C. Mammalian multidrug-resistance proteins (MRPs). Essays Biochem 2011, 50, 179–207. [Google Scholar]
- Chang, X.B. A molecular understanding of ATP-dependent solute transport by multidrug resistance-associated protein MRP1. Cancer Metastasis Rev 2007, 26, 15–37. [Google Scholar]
- Özvegy-Laczka, C.; Cserepes, J.; Elkind, N.B.; Sarkadi, B. Tyrosine kinase inhibitor resistance in cancer: Role of ABC multidrug transporters. Drug Resist. Updat 2005, 8, 15–26. [Google Scholar]
- Li, J.; Li, Z.N.; Yuc, L.C.; Bao, Q.L.; Wud, J.R.; Shic, S.B.; Li, X.Q. Association of expression of MRP1, BCRP, LRP and ERCC1 with outcome of patients with locally advanced non-small cell lung cancer who received neoadjuvant chemotherapy. Lung Cancer 2010, 69, 116–122. [Google Scholar]
- Penson, R.T.; Oliva, E.; Skates, S.J.; Glyptis, T.; Fuller, A.F., Jr; Goodman, A.; Seiden, M.V. Expression of multidrug resistance-1 protein inversely correlates with paclitaxel response and survival in ovarian cancer patients: A study in serial samples. Gynecol. Oncol. 2004, 93, 98–106. [Google Scholar]
- Zhang, D.; Fan, D. New insights into the mechanisms of gastric cancer multidrug resistance and future perspectives. Future Oncol 2010, 6, 527–537. [Google Scholar]
- Henderson, M.J.; Haber, M.; Porro, A.; Munoz, M.A.; Iraci, N.; Xue, C.; Murray, J.; Flemming, C.L.; Smith, J.; Fletcher, J.I.; et al. ABCC multidrug transporters in childhood neuroblastoma: Clinical and biological effects independent of cytotoxic drug efflux. J. Natl. Cancer Inst 2011, 103, 1236–1251. [Google Scholar]
- Narasaki, F.; Oka, M.; Nakano, R.; Ikeda, K.; Fukuda, M.; Nakamura, T.; Soda, H.; Nakagawa, M.; Kuwan, M.; Kohno, S. Human canalicular multispecific organic anion transporter (cMOAT) is expressedin human lung, gastric, and colorectal cancer cells. Biochem. Biophys. Res. Commun 1997, 240, 606–611. [Google Scholar]
- Steinbach, D.; Wittig, S.; Cario, G.; Viehmann, S.; Mueller, A.; Gruhn, B.; Haefer, R.; Zintl, F.; Sauerbrey, A. The multidrug resistance-associated protein 3 (MRP3) is associated with a poor outcome in childhood ALL and may account for the worse prognosis in male patients and T-cell immunophenotype. Blood 2003, 102, 4493–4498. [Google Scholar]
- Yamasaki, M.; Makino, T.; Masuzawa, T.; Kurokawa, Y.; Miyata, H.; Takiguchi, S.; Nakajima, K.; Fujiwara, Y.; Matsuura, N.; Mori, M.; et al. Role of multidrug resistance protein 2 (MRP2) in chemoresistance and clinical outcome in oesophageal squamous cell carcinoma. Br. J. Cancer 2011, 104, 707–713. [Google Scholar]
- Korita, P.V.; Wakai, T.; Shirai, Y.; Matsuda, Y.; Sakata, J.; Takamura, M.; Yano, M.; Sanpei, A.; Aoyagi, Y.; Hatakeyama, K.; et al. Multidrug resistance-associated protein 2 determines the efficacy of cisplatin in patients with hepatocellular carcinoma. Oncol. Rep 2010, 23, 965–972. [Google Scholar]
- Borst, P.; Wolf, C.; van de Wetering, K. Multidrug resistance-associated proteins 3, 4, and 5. Pfluger Arch 2007, 453, 661–673. [Google Scholar]
- Konig, J.; Hartel, M.; Nies, A.T.; Martignoni, M.E.; Guo, J.; Buchler, M.W.; Friess, H.; Keppler, D. Expression and localization of human multidrug resistance protein (ABCC) family members in pancreatic carcinoma. Int. J. Cancer 2005, 115, 359–367. [Google Scholar]
- Nies, A.T.; Konig, J.; Pfannschmidt, M.; Klar, E.; Hofmann, W.J.; Keppler, D. Expression of the multidrug resistance proteins MRP2 and MRP3 in human hepatocellular carcinoma. Int. J. Cancer 2001, 94, 492–499. [Google Scholar]
- Young, L.C.; Campling, B.G.; Cole, S.P.C.; Deeley, R.G.; Gerlach, J.H. Multidrug Resistance proteins MRP3, MRP1, and MRP2 in lung cancer: Correlation of protein levels with drug response and messenger RNA levels. Clin. Cancer Res 2001, 7, 1798–1804. [Google Scholar]
- Ohishi, Y.; Oda, Y.; Uchiumi, T.; Kobayashi, H.; Hirakawa, T.; Miyamoto, S.; Kinukawa, N.; Nakano, H.; Kuwano, M.; Tsuneyoshi, M. ATP-binding cassette superfamily transporter gene expression in human primary ovarian carcinoma. Clin. Cancer Res 2002, 8, 3767–3775. [Google Scholar]
- Russel, F.G.; Koenderink, J.B.; Masereeuw, R. Multidrug resistance protein 4 (MRP4/ABCC4): A versatile efflux transporter for drugs and signalling molecules. Trends Pharmacol. Sci 2008, 29, 200–207. [Google Scholar]
- Norris, M.D.; Smith, J.; Tanabe, K.; Tobin, P.; Flemming, C.; Scheffer, G.L.; Wielinga, P.; Cohn, S.L.; London, W.B.; Marshall, G.M.; et al. Expression of multidrug transporter MRP4/ABCC4 is a marker of poor prognosis in neuroblastoma and confers resistance to irinotecan in vitro. Mol. Cancer Ther 2005, 4, 547–553. [Google Scholar]
- Fernandes, J.; Castilho, R.O.; Costa, M.R.; Wagner-Souza, K.; Kaplan, M.A.; Gattass, C.R. Pentacyclic triterpenes from Chrysobalanaceae species: Cytotoxicity on multidrug resistant and sensitive leukemia cell lines. Cancer Lett. 2003, 190, 165–169. [Google Scholar]
- Vasconcelos, F.C.; Gattass, C.G.R.; Rumjanek, V.M.; Maia, R.C. Pomolic acid-induced apoptosis in cells from patients with chronic myeloid leukemia exhibiting different drug resistance profile. Invest. New Drugs 2007, 25, 525–533. [Google Scholar]
- Yoshida, N.; Takada, T.; Yamamura, Y.; Adachi, I.; Suzuki, H.; Kawakami, J. Inhibitory effects of terpenoids on multidrug resistance-associated protein 2- and breast cancer resistance protein mediated transport. Drug Metab. Dispos 2008, 36, 1206–1211. [Google Scholar]
- Lúcio, K.A.; Rocha, G.G.; Monção-Ribeiro, L.C.; Fernandes, J.; Takiya, C.M.; Gattass, C.R. Oleanolic acid initiates apoptosis in non-small cell lung cancer cell lines and reduces metastasis of a B16F10 melanoma model in vivo. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Rocha, G.G.; Simões, M.; Oliveira, R.R.; Kaplan, M.A.C.; Gattass, C.R. 3β-acetyl tormentic acid induces apoptosis of resistant leukemia cells independently of P-gp/ABCB1 activity or expression. Invest. New Drugs 2012, 30, 105–113. [Google Scholar]
- Hooijberg, J.H.; Broxterman, H.J.; Kool, M.; Assaraf, Y.G.; Peters, G.J.; Noordhuis, P.; Scheper, R.J.; Borst, P.; Pinedo, H.M.; Jansen, G. Antifolate Resistance mediated by the multidrug resistance proteins MRP1 and MRP2. Cancer Res 1999, 9, 2532–2535. [Google Scholar]
- Zeng, H.; Bain, L.J.; Belinsky, M.G.; Kruh, G.D. Expression of multidrug resistance protein-3 (multispecific organic anion transporter-D) in human embryonic kidney 293 cells confers resistance to anticancer agents. Cancer Res 1999, 59, 5964–5967. [Google Scholar]
- Lee, K.; Klein-Szanto, A.J.P.; Kruh, G.D. Analysis of the MRP4 drug resistance profile in transfected NIH3T3 cells. J. Nat. Cancer Inst 2000, 92, 1934–1940. [Google Scholar]
- Braga, F.; Ayres-Saraiva, D.; Gattass, C.R.; Capella, M.A.M. Oleanolic acid inhibits the activity of the multidrug resistance protein ABCC1 (MRP1) but not of the ABCB1 (P-glycoprotein): Possible use in cancer chemotherapy. Cancer Lett 2007, 248, 147–152. [Google Scholar]
- Torky, A.R.W.; Stehfest, E.; Viehweger, K.; Taege, C.; Foth, H. Immuno-histochemical detection of MRPs in human lung cells in culture. Toxicology 2005, 207, 437–450. [Google Scholar]
- Leslie, E.M.; Deeley, R.G.; Cole, S.P.C. Multidrug resistance proteins: Role of P-glycoprotein, MRP1, MRP2, and BCRP (ABCG2) in tissue defense. Toxicol. Appl. Pharmacol 2005, 204, 216–237. [Google Scholar]
- Choi, C.H. ABC transporters as multidrug resistance mechanisms and the development of chemosensitizers for their reversal. Cancer Cell Int 2005, 5, 1–13. [Google Scholar]
- Gekeler, V.; Ise, W.; Sanders, K.H.; Ulrich, W.R.; Beck, J. The leukotriene LTD(4) receptor antagonist MK571 specifically modulates MRP associated multidrug-resistance. Biochem. Biophys. Res. Comm 1995, 208, 345–352. [Google Scholar]
- Norman, B.H.; Gruber, J.M.; Hollinshead, S.P.; Wilson, J.W.; Starling, J.J.; Law, K.L.; Self, T.D.; Tabas, L.B.; Williams, D.C.; Paul, D.C.; et al. Cyclohexyl-linked tricyclic isoxazoles are potent and selective modulators of the multidrug resistance protein (MRP1). Bioorg. Med. Chem. Lett 2005, 15, 5526–5530. [Google Scholar]
- Zhou, S.F.; Wang, L.L.; Di, Y.M.; Xue, C.C.; Duan, W.; Li, C.G.; Li, Y. Substrates and inhibitors of human multidrug resistance associated proteins and the implications in drug development. Curr. Med. Chem 2008, 15, 1981–2039. [Google Scholar]
- Deeley, R.G.; Cole, S.P.C. Substrate recognition and transport by multidrug resistance protein 1 (ABCC1). FEBS Lett 2006, 580, 1103–1111. [Google Scholar]
- Letschert, K.; Komatsu, M.; Hummel-Eisenbeiss, J.; Keppler, D. Vectorial transport of the peptide CCK-8 by double-transfected MDCKII cells stably expressing the organic anion transporter OATP1B3 (OATP8) and the export pump ABCC2. J. Pharmacol. Exp. Ther 2005, 313, 549–556. [Google Scholar]
- Rius, M.; Nies, A.T.; Hummel-Eisenbeiss, J.; Jedlitschky, G.; Keppler, D. Cotransport of reduced glutathione with bile salts by MRP4 (ABCC4) localized to the basolateral hepatocyte membrane. Hepatology 2003, 38, 374–384. [Google Scholar]
- Reid, G.; Wielinga, P.; Zelcer, N.; van der Heijden, I.; Kuil, A.; de Haas, M.; Wijnholds, J.; Borst, P. The human multidrug resistance protein MRP4 functions as a prostaglandin efflux transporter and is inhibited by nonsteroidal antiinflammatory drugs. Proc. Natl. Acad. Sci. USA 2003, 100, 9244–9249. [Google Scholar]
- Rocha, G.G.; Simões, M.; Lúcio, K.A.; Oliveira, R.R.; Kaplan, M.A.C.; Gattass, C.R. Natural triterpenoids from Cecropia lyratiloba are cytotoxic to both sensitive and multidrug resistant leukemia cell lines. Bioorg. Med. Chem 2007, 15, 7355–7360. [Google Scholar]
- Rabindran, S.K.; Ross, D.D.; Doyle, L.A.; Yang, W.; Greenberger, L.M. Fumitremorgin C reverses multidrug resistance in cells transfected with the breast cancer resistance protein. Cancer Res 2000, 60, 47–50. [Google Scholar]
- Chearwae, W.; Anuchapreeda, S.; Nandigama, K.; Ambudkar, S.V.; Limtrakul, P. Biochemical mechanism of modulation of human P-glycoprotein (ABCB1) by curcumin I, II, and III purified from Turmeric powder. Biochem. Pharmacol 2004, 68, 2043–2052. [Google Scholar]
- Wu, C.P.; Calcagno, A.M.; Hladky, S.B.; Ambudkar, S.V.; Barrand, M.A. Modulatory effects of plant phenols on human multidrug-resistance proteins 1, 4 and 5 (ABCC1, 4 and 5). FEBS J 2005, 272, 4725–4740. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibition Ratio | ||||||
|---|---|---|---|---|---|---|
| Treatment | B16F10 | Ma104 | A549 | 2008/MRP2 | HEK/MRP3 | 3T3/MRP4 |
| MK50 | 10.5 ± 1.7 | 5.4 ± 0.3 | 7.0 ± 1.1 | 1.8 ± 0.3 | 1.5 ± 0.2 | 3.7 ± 0.7 |
| MK100 | - | - | - | 3.6 ± 0.3 | 3.7 ± 0.1 | 6.1 ± 1.4 |
| PRO 1000 | - | - | - | - | 6.0 ± 0.1 | 7.7 ± 1.3 |
| 3ATA 6.25 | 5.6 ± 1.2 | 2.5 ± 0.1 | 2.9 ± 0.2 | 1.8 ± 0.2 | 1.2 ± 0.3 | 2.3 ± 0.4 |
| 3ATA 12.5 | 11.4 ± 1.3 | 3.2 ± 0.1 | 3.0 ± 0.3 | 2.7 ± 0.1 | 1.4 ± 0.3 | 3.1 ± 0.5 |
| 3ATA 25.0 | 20.9 ± 3.4 | 5.4 ± 0.2 | 4.0 ± 0.4 | 3.5 ± 0.2 | 1.7 ± 0.4 | 3.5 ± 0.3 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Da Graça Rocha, G.; Simões, M.; Oliveira, R.R.; Kaplan, M.A.C.; Gattass, C.R. Effects of 3β-Acethyl Tormentic Acid (3ATA) on ABCC Proteins Activity. Int. J. Mol. Sci. 2012, 13, 6757-6771. https://doi.org/10.3390/ijms13066757
Da Graça Rocha G, Simões M, Oliveira RR, Kaplan MAC, Gattass CR. Effects of 3β-Acethyl Tormentic Acid (3ATA) on ABCC Proteins Activity. International Journal of Molecular Sciences. 2012; 13(6):6757-6771. https://doi.org/10.3390/ijms13066757
Chicago/Turabian StyleDa Graça Rocha, Gleice, Marisol Simões, Rodrigo Rodrigues Oliveira, Maria Auxiliadora Coelho Kaplan, and Cerli Rocha Gattass. 2012. "Effects of 3β-Acethyl Tormentic Acid (3ATA) on ABCC Proteins Activity" International Journal of Molecular Sciences 13, no. 6: 6757-6771. https://doi.org/10.3390/ijms13066757