Protective Roles of Gadd45 and MDM2 in Blueberry Anthocyanins Mediated DNA Repair of Fragmented and Non-Fragmented DNA Damage in UV-Irradiated HepG2 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
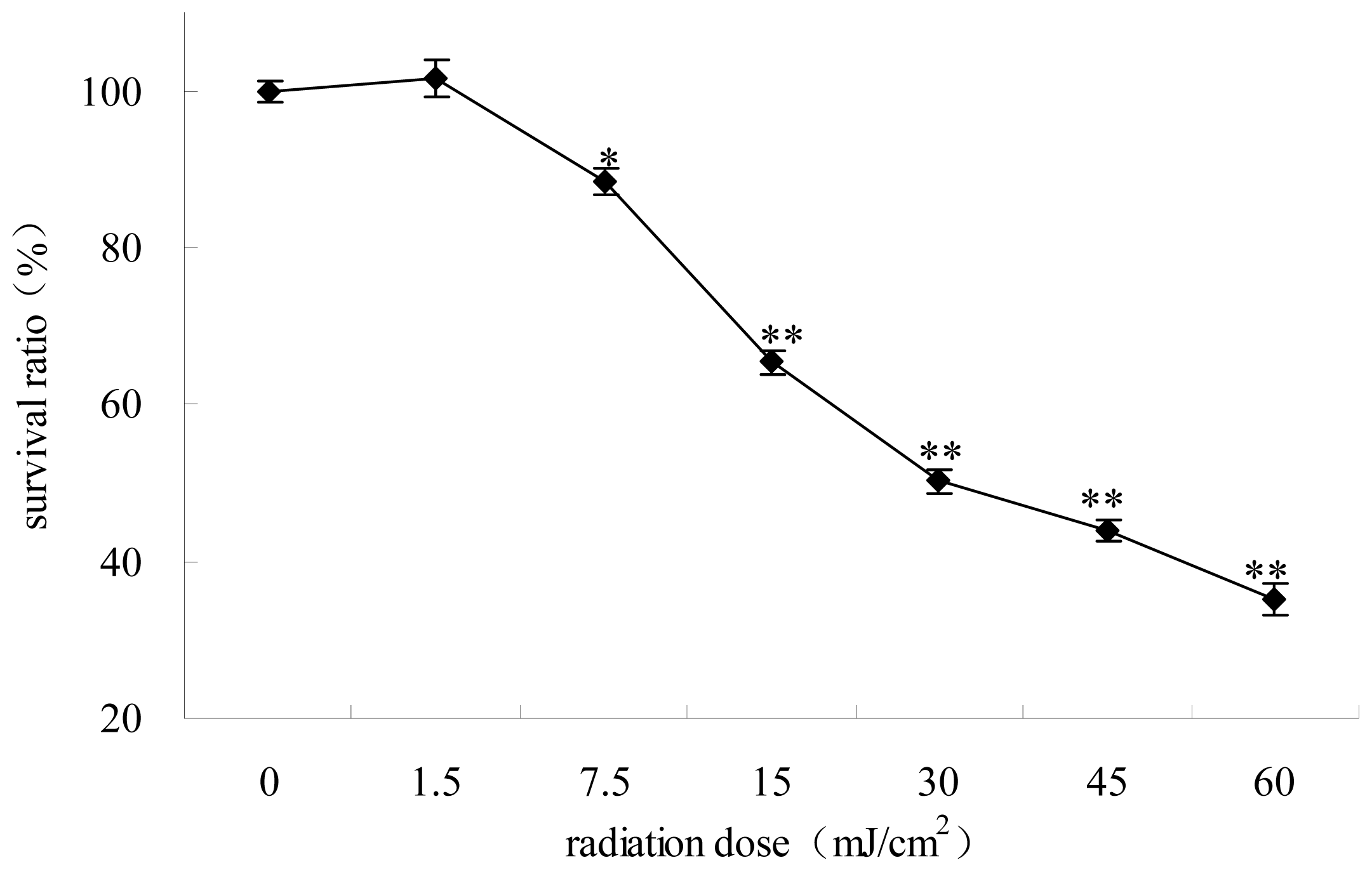
2.1. The Survival Ratios of HepG2 Cells Irradiated at Different UV Dose
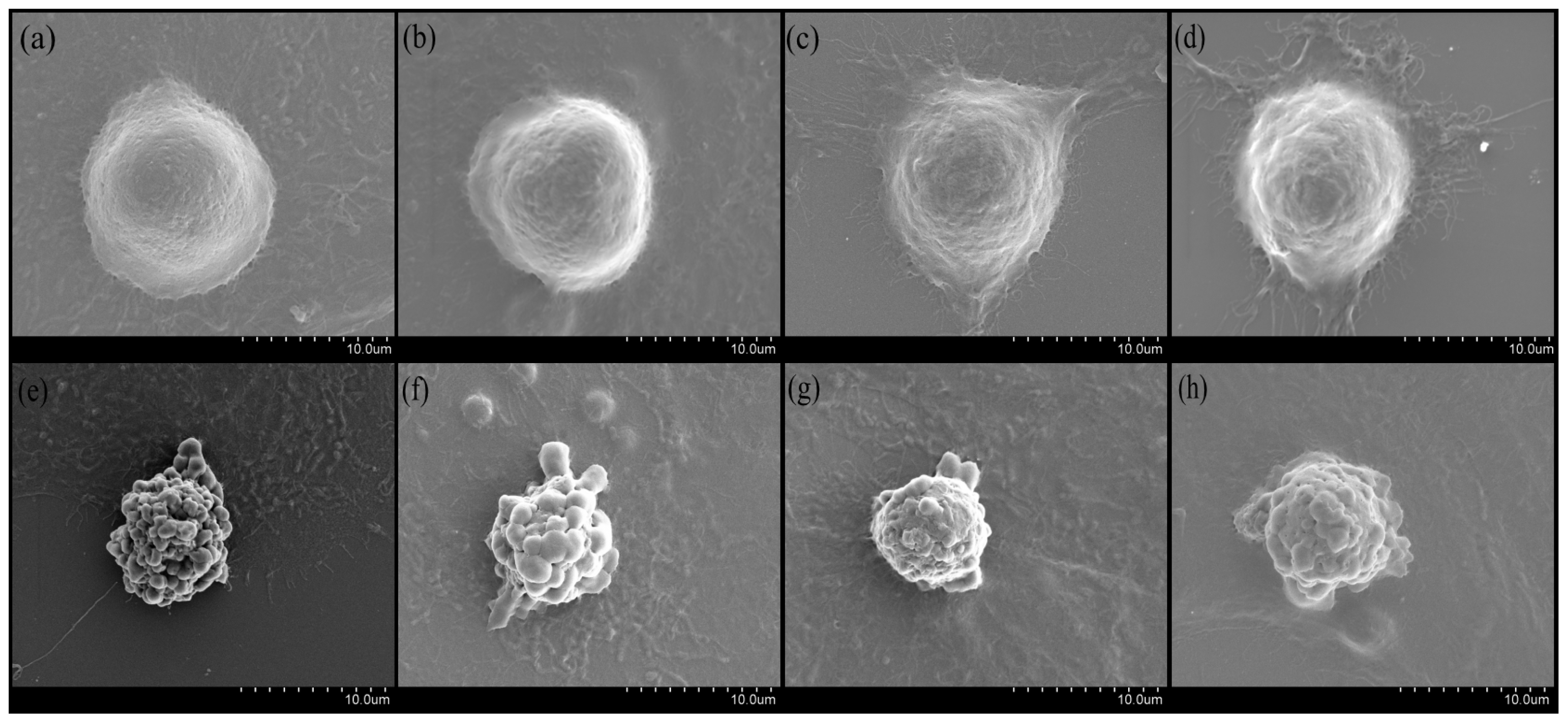
2.2. The Effects of BA on Morphology of UV-Irradiated HepG2 Cells Observed by SEM
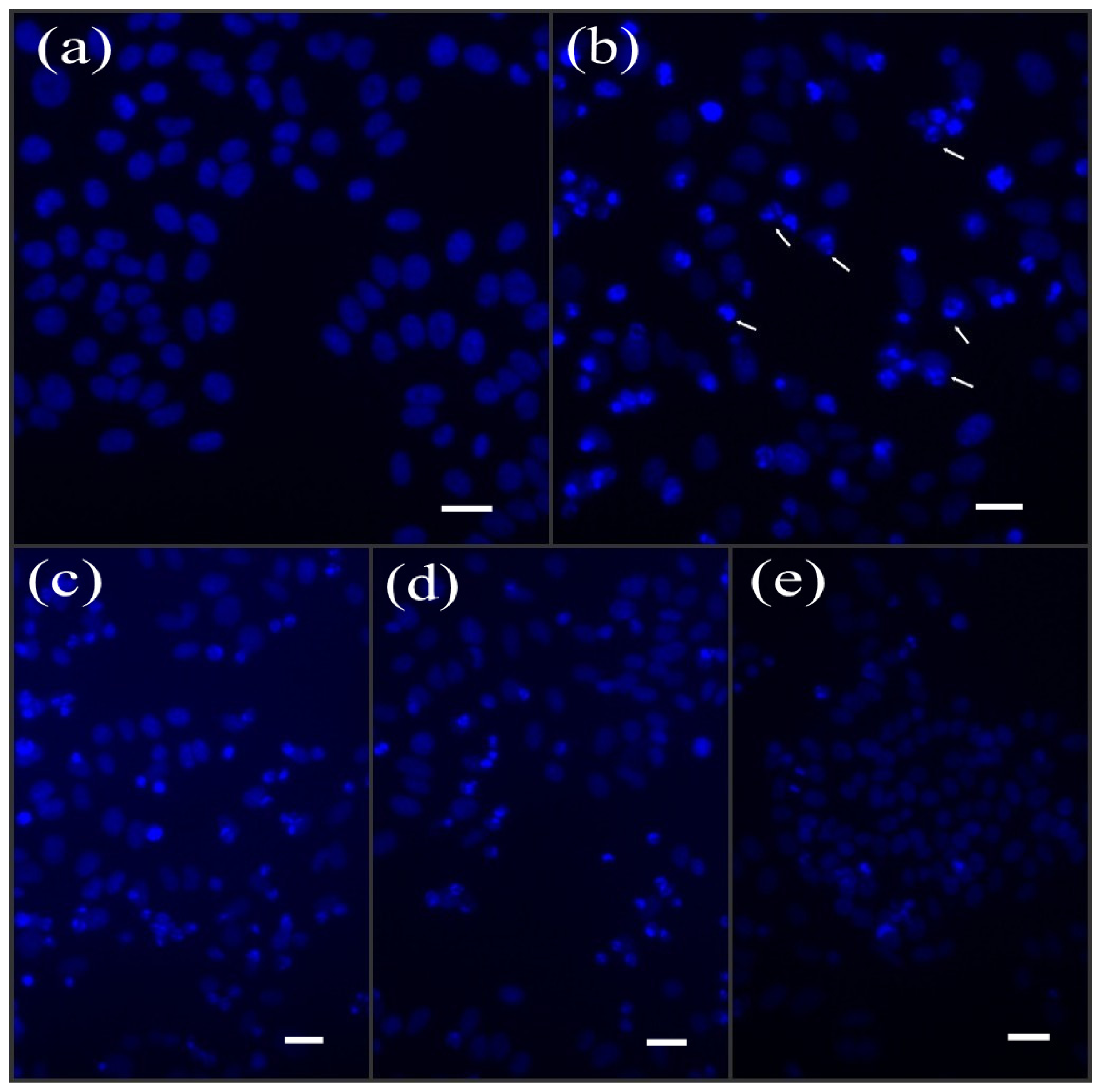
2.3. BA Restored UV-Induced DNA Fragmentation in HepG2 Cells
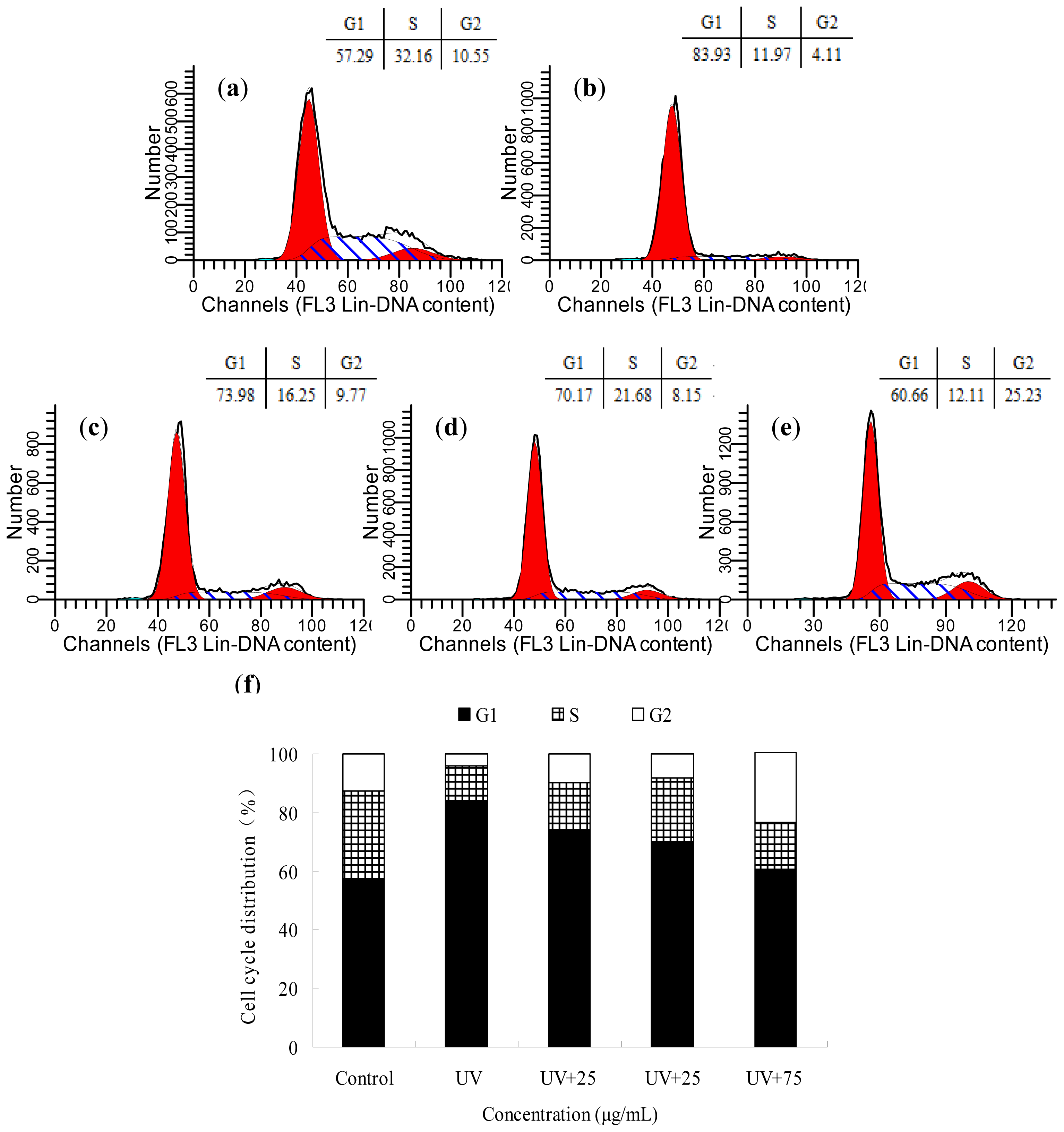
2.4. The Effects of BA on the Cell Cycle Distribution of UV-Irradiated HepG2 Cells
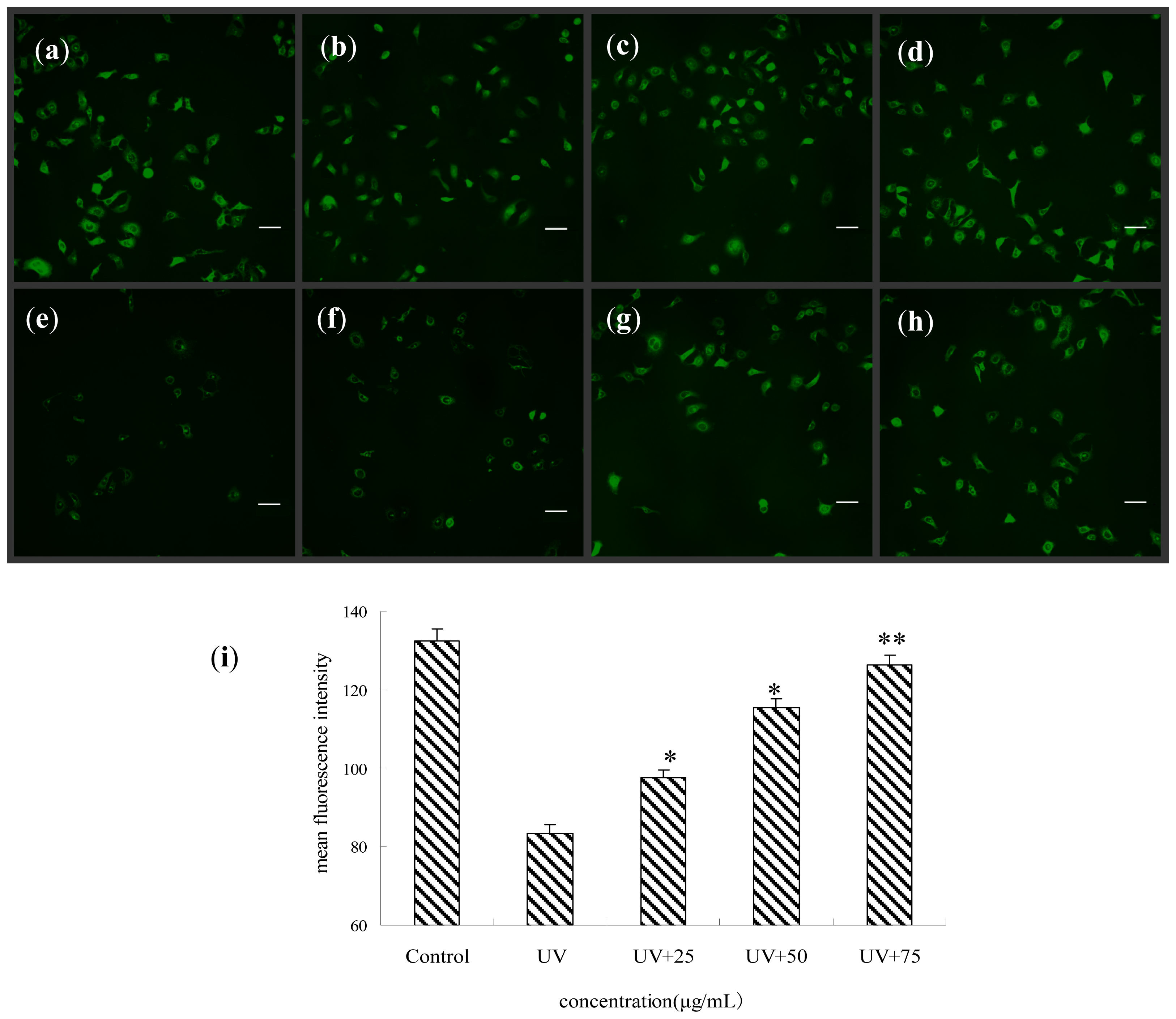
2.5. Effect of BA on the Membrane Potential of Mitochondria of UV-Irradiated HepG2 Cells
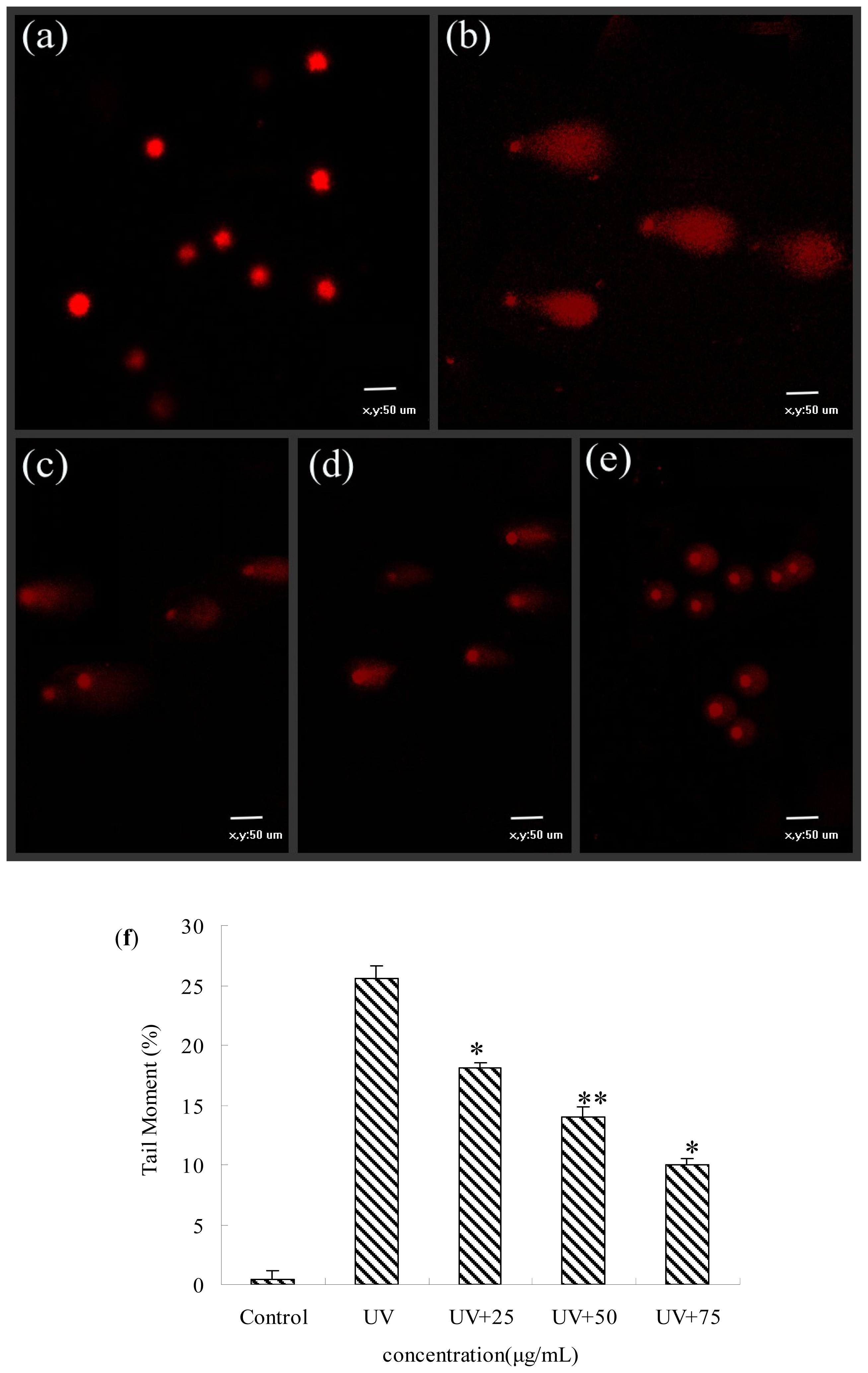
2.6. Protective Effects of BA Extracts on UV-Induced DNA Damage by Comet Assay
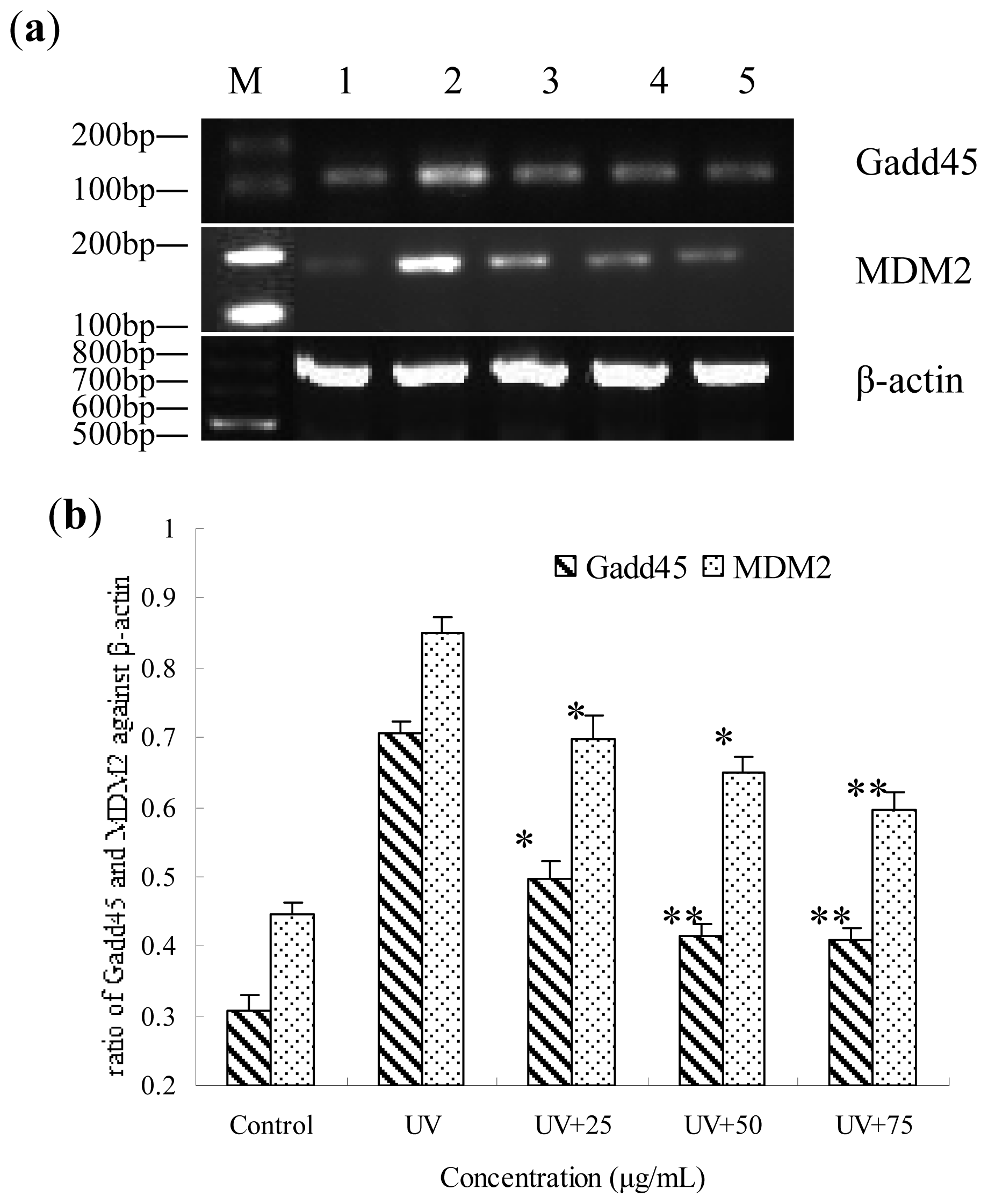
2.7. Gene Expression of Gadd45 and MDM2 Influenced by BA on UV-Irradiated HepG2 Cells
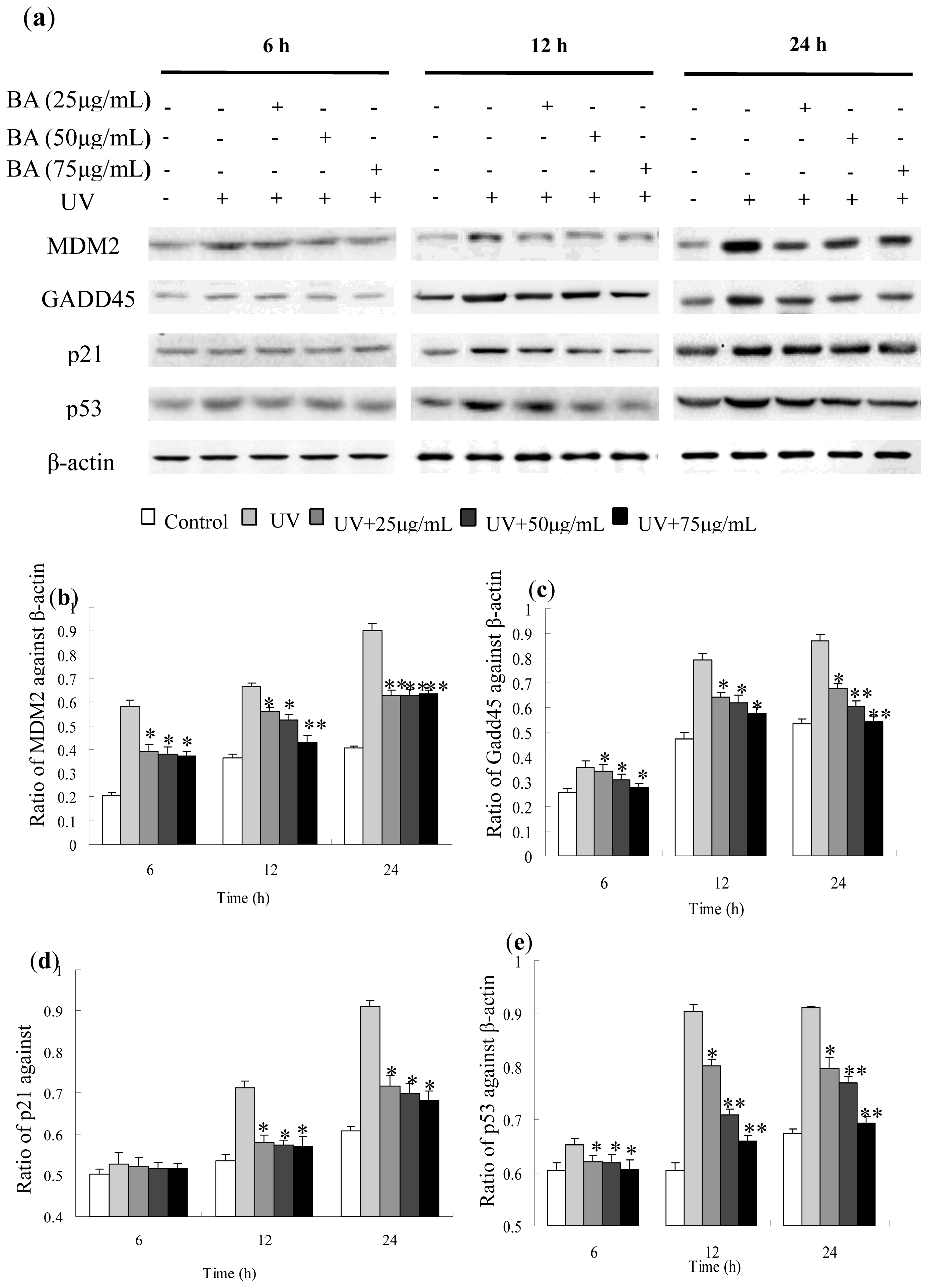
2.8. Protein Expression of Gadd45, MDM2, p53 and p21 Influenced by BA on UV-Irradiated HepG2 Cells
3. Discussion
4. Experimental Section
4.1. Drugs and Antibodies
4.2. Isolation and Purification of Blueberry Anthocyanins
4.3. Cell Culture and UV Irradiation
4.4. The Survival Ratios of HepG2 Cells Irradiated at Different UV Dose
4.5. Morphologic Observations
4.6. Nuclear Morphology by Hoechst 33258 Staining
4.7. Cell Cycle Analysis
4.8. Analysis of Mitochondrial Potential (ΔΨm)
4.9. Comet Assay
4.10. Gene Expression of Gadd45 and MDM2 by RT-PCR
4.11. Protein Expression of Gadd45, MDM2, p53 and p21 by Western Blotting
4.12. Statistics Analysis
5. Conclusions








Acknowledgments
Conflicts of Interest
References
- Zhou, B.B.S.; Elledge, S.J. The DNA damage response: Putting checkpoints in perspective. Nature 2000, 408, 433–439. [Google Scholar]
- Oda, K.; Arakawa, H.; Tanaka, T. p53AIP1, a potential mediator of p53-dependent apoptosis, and its regulation by Ser-46-phosphorylated p53. Cell 2000, 102, 849–862. [Google Scholar]
- Elledge, S.J. Cell cycle checkpoints: Preventing and identity crisis. Science 1996, 274, 1664–1672. [Google Scholar]
- Kastan, M.B.; Bartek, J. Cell-cycle checkpoints and cancer. Nature 2004, 432, 316–323. [Google Scholar]
- Kops, G.J.P.L.; Weaver, B.A.A.; Cleveland, D.W. On the road to cancer: Aneuploidy and the mitotic checkpoint. Nat. Rev. Cancer 2005, 5, 773–785. [Google Scholar]
- Shen, Y.H.; Utama, B.; Wang, J. Human cytomegalovirus causes endothelial injury through the ataxia telangiectasia mutant and p53 DNA damage signaling pathways. Circ. Res 2004, 94, 1310–1317. [Google Scholar]
- Roos, W.P.; Kaina, B. DNA damage-induced cell death by apoptosis. Trends Mol. Med 2006, 12, 440–450. [Google Scholar]
- Bartek, J.; Lukas, J. DNA damage checkpoints: From initiation to recovery or adaptation. Curr. Opin. Cell. Biol 2007, 2, 238–245. [Google Scholar]
- Reed, E. Damage and repair in translational oncology: An overview. Clin. Cancer Res 2010, 18, 4511–4516. [Google Scholar]
- Zhan, Q.; Chen, I.; Antinore, M.J.; Fornace, A.J. Tumor suppressor p53 can participate in transcriptional induction of the GADD45 promoter in the absence of direct DNA binding. Mol. Cell Biol 1998, 5, 2768–2778. [Google Scholar]
- Moskalev, A.A.; Smit-McBride, Z.; Shaposhnikov, M.V. Gadd45 proteins: Relevance to aging, longevity and age -related pathologies. Ageing Res. Rev 2011, 11, 51–66. [Google Scholar]
- Shih, R.S.M.; Wong, S.H.K.; Schoene, N.W.; Zhang, J.J.; Lei, K.Y. Enhanced Gadd45 expression and delayed G2/M progression are p53-dependent in zinc-supplemented human bronchial epithelial cells. Exp. Biol. Med 2010, 8, 932–940. [Google Scholar]
- Warmerdama, D.O.; Kanaar, R. Dealing with DNA damage: Relationships between checkpoint and repair pathways. Mutat. Res 2010, 1, 2–11. [Google Scholar]
- Kim, S.H.; Hur, Y.J.; Lee, S.J.; Kim, S.J.; Park, C.G.; Oh, Y.K.; Jung, W.W.; Seo, J.B.; Nam, M.H.; Choi, I. E6 and E7 fusion immunoglobulin from human papilloma virus 16 induces dendritic cell maturation and antigen specific activation of T helper 1 response. Biotechnol. Lett 2011, 4, 663–671. [Google Scholar]
- Nyberg, K.A.; Michelson, R.J.; Putnam, C.W.; Weinert, T.A. Toward maintaining the genome: DNA damage and replication checkpoints. Annu. Rev. Genet 2002, 36, 617–656. [Google Scholar]
- Khosravi, R.; Maya, R.; Gottlieb, T.; Oren, M.; Shiloh, Y.; Shkedy, D. Rapid ATM-dependent phosphorylation of MDM2 precedes p53 accumulation in response to DNA damage. Proc. Natl. Acad. Sci. USA 1999, 26, 14973–14977. [Google Scholar]
- Iliakis, G.; Wang, Y.; Guan, J.; Wang, H. DNA damage checkpoint control in cells exposed to ionizing radiation. Oncogene 2003, 22, 5834–5847. [Google Scholar]
- Yang, J.; Yu, Y.; Hamrick, H.E.; Duerksen-Hughes, P.J. ATM, ATR and DNA-PK: Initiators of the cellular genotoxic stress responses. Carcinogenesis 2003, 10, 1571–1580. [Google Scholar]
- Liu, W.; Lu, X.Y.; He, G.Y.; Gao, X.; Li, M.X.; Wu, J.H.; Li, Z.J.; Wu, J.H.; Wang, J.C.; Luo, C. Cytosolic protection against ultraviolet induced DNA damage by blueberry anthocyanins and anthocyanidins in hepatocarcinoma HepG2 cells. Biotechnol. Lett 2013, 35, 491–498. [Google Scholar]
- Long, X.L.; Goldenthal, M.J.; Marín-García, J. Oxidative stress enhances phosphorylation of p53 in neonatal rat cardiomyocytes. Mol. Cell. Biochem 2007, 1–2, 167–174. [Google Scholar]
- Serranoa, M.C.; Pagania, R.; Manzanob, M.; Comasc, J.V.; Portolésa, M.T. Mitochondrial membrane potential and reactive oxygen species content of endothelial and smooth muscle cells cultured on poly (ɛ-caprolactone) films. Biomaterials 2006, 27, 4706–4714. [Google Scholar]
- Gadbois, D.M.; Crissman, H.A.; Nastasi, A.; Habbersett, R.; Wang, S.; Chen, D.; Lehnert, B.E. Alterations in the progression of cells through the cell cycle after exposure to alpha particles or gamma rays. Radiat. Res 1996, 146, 414–424. [Google Scholar]
- Bae, I.; Fan, S.; Bhatia, K.; Kohn, K.W.; Fornace, A.J.; O’Connor, P.M. Relationships between G1 arrest and stability of the p53 and p21Cip1/Waf1 proteins following γ-irradiation of human lymphoma cells. Cancer Res 1995, 55, 2387–2394. [Google Scholar]
- Chen, C.Y.; Oliner, J.D.; Zhan, Q.; Fornace, A.J.; Vogelstein, J.B.; Kastan, M.B. Interactions between p53 and MDM2 in a mammalian cell cycle checkpoint pathway. Proc. Natl. Acad. Sci. USA 1994, 7, 2684–2688. [Google Scholar]
- Nicoue, E.E.; Savard, S.; Belkacemi, K. Anthocyanins in wild blueberries of Quebec: Extraction and identification. J. Agric. Food Chem 2007, 55, 5626–5635. [Google Scholar]
- Zhou, K.Q.; Yu, L.L. Effects of extraction solvent on wheat bran antioxidant activity estimation. Food Sci. Technol 2004, 37, 717–721. [Google Scholar]
- Prior, R.L.; Lazarus, S.A.; Cao, G.H.; Muccitelli, H.; Hammerstone, J.F. Identification of procyanidins and anthocyanins in blueberries and cranberries (Vaccinium Spp.) using high-performance liquid chromatography/mass spectrometry. J. Agric. Food Chem 2001, 49, 1270–1276. [Google Scholar]
- Williams, G.C.; Liu, A.; Knipp, G.; Sinko, P.J. Direct evidence that saquinavir is transported by multidrug resistance-associated protein (MRP1) and canalicular multispecific organic anion transporter (MRP2). Antimicrob. Agents Chemother 2002, 46, 3456–3462. [Google Scholar]
- Lu, X.Y.; Liu, W.; Wu, J.H.; Li, M.X.; Wang, J.C.; Wu, J.H.; Luo, C. A polysaccharide fraction of adlay seed (Coixlachryma-jobi L.) induces apoptosis in human non-small cell lung cancer A549 cells. Biochem. Biophys. Res. Commun 2012, 430, 846–851. [Google Scholar]
- Heemels, M.T.; Dhand, R.; Allen, L. The biochemistry of apoptosis. Nature 2000, 350, 770–776. [Google Scholar]
- Lemasters, J.J.; Nieminen, A.L. Mitochondrial oxygen radical formation during reductive and oxidative stress to intact hepatocytes. Biosci. Rep 1997, 17, 281–291. [Google Scholar]
- Sancar, A.; Lindsey-Boltz, L.A.; Ünsal-Kaçmaz, K.; Linn, S. Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints. Annu. Rev. Biochem 2004, 73, 39–85. [Google Scholar]
- Mozaffarieh, M.; Schoetzau, A.; Sauter, M.; Grieshaber, M.; Orgul, S.; Golubnitschaja, O.; Flammer, J. Comet assay analysis of single-stranded DNA breaks in circulating leukocytes of glaucoma patients. Mol. Vis 2008, 14, 1584–1588. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, W.; Lu, X.; He, G.; Gao, X.; Xu, M.; Zhang, J.; Li, M.; Wang, L.; Li, Z.; Wang, L.; et al. Protective Roles of Gadd45 and MDM2 in Blueberry Anthocyanins Mediated DNA Repair of Fragmented and Non-Fragmented DNA Damage in UV-Irradiated HepG2 Cells. Int. J. Mol. Sci. 2013, 14, 21447-21462. https://doi.org/10.3390/ijms141121447
Liu W, Lu X, He G, Gao X, Xu M, Zhang J, Li M, Wang L, Li Z, Wang L, et al. Protective Roles of Gadd45 and MDM2 in Blueberry Anthocyanins Mediated DNA Repair of Fragmented and Non-Fragmented DNA Damage in UV-Irradiated HepG2 Cells. International Journal of Molecular Sciences. 2013; 14(11):21447-21462. https://doi.org/10.3390/ijms141121447
Chicago/Turabian StyleLiu, Wei, Xiangyi Lu, Guangyang He, Xiang Gao, Maonian Xu, Jingkai Zhang, Meiling Li, Lifeng Wang, Zhenjing Li, Likui Wang, and et al. 2013. "Protective Roles of Gadd45 and MDM2 in Blueberry Anthocyanins Mediated DNA Repair of Fragmented and Non-Fragmented DNA Damage in UV-Irradiated HepG2 Cells" International Journal of Molecular Sciences 14, no. 11: 21447-21462. https://doi.org/10.3390/ijms141121447