Combination BMSC and Niaspan Treatment of Stroke Enhances White Matter Remodeling and Synaptic Protein Expression in Diabetic Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Objective
Research design and methods
Results
Conclusions
1. Introduction
2. Results
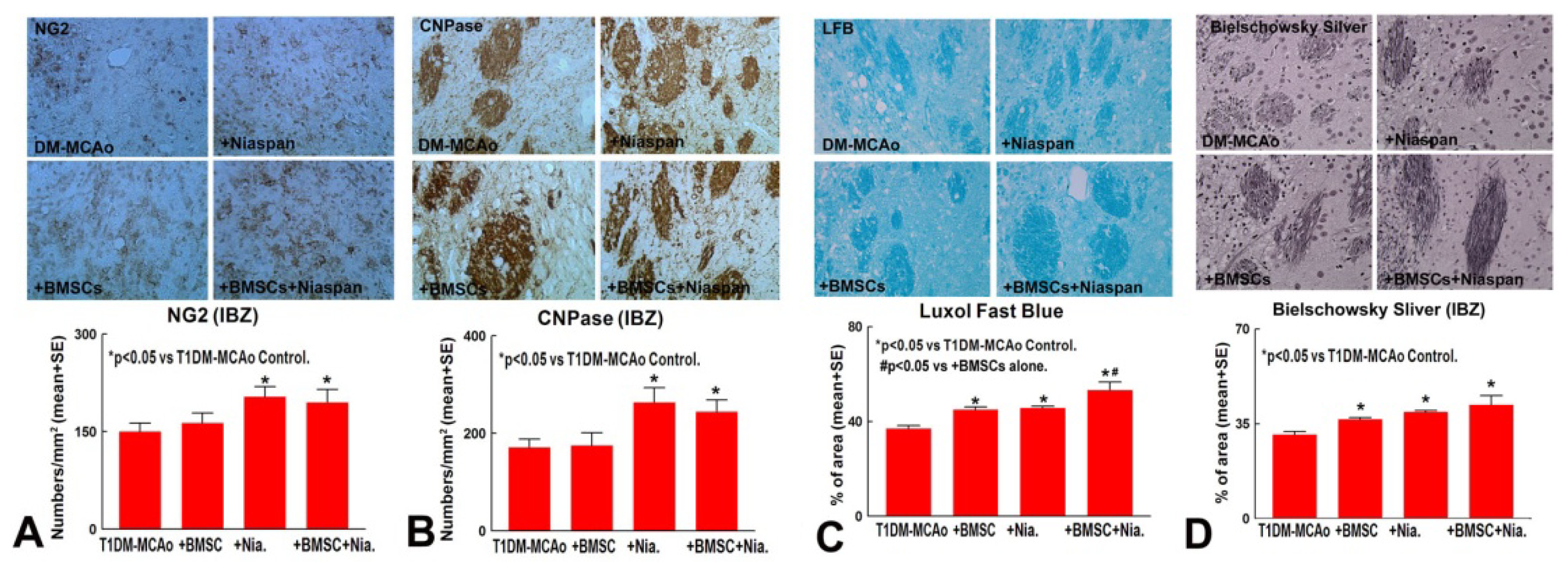
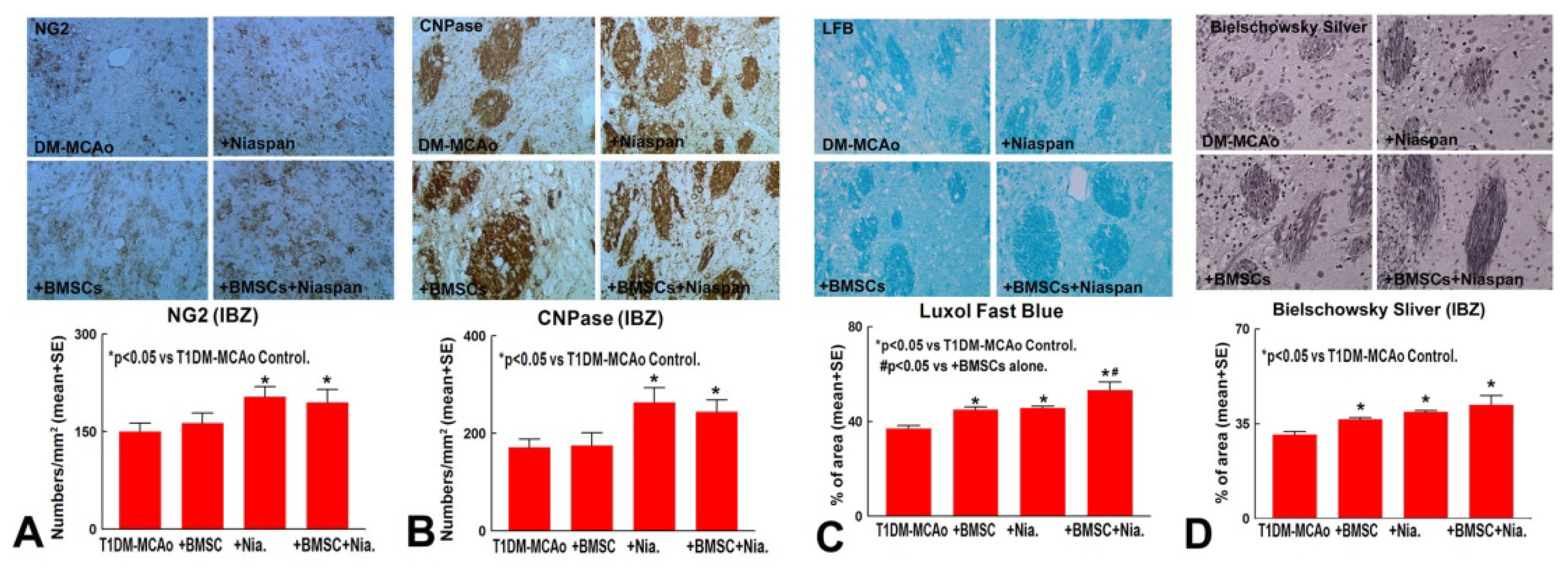
2.1. Combination BMSCs and Niaspan Treatment of Stroke Increases White Matter (WM) Remodeling
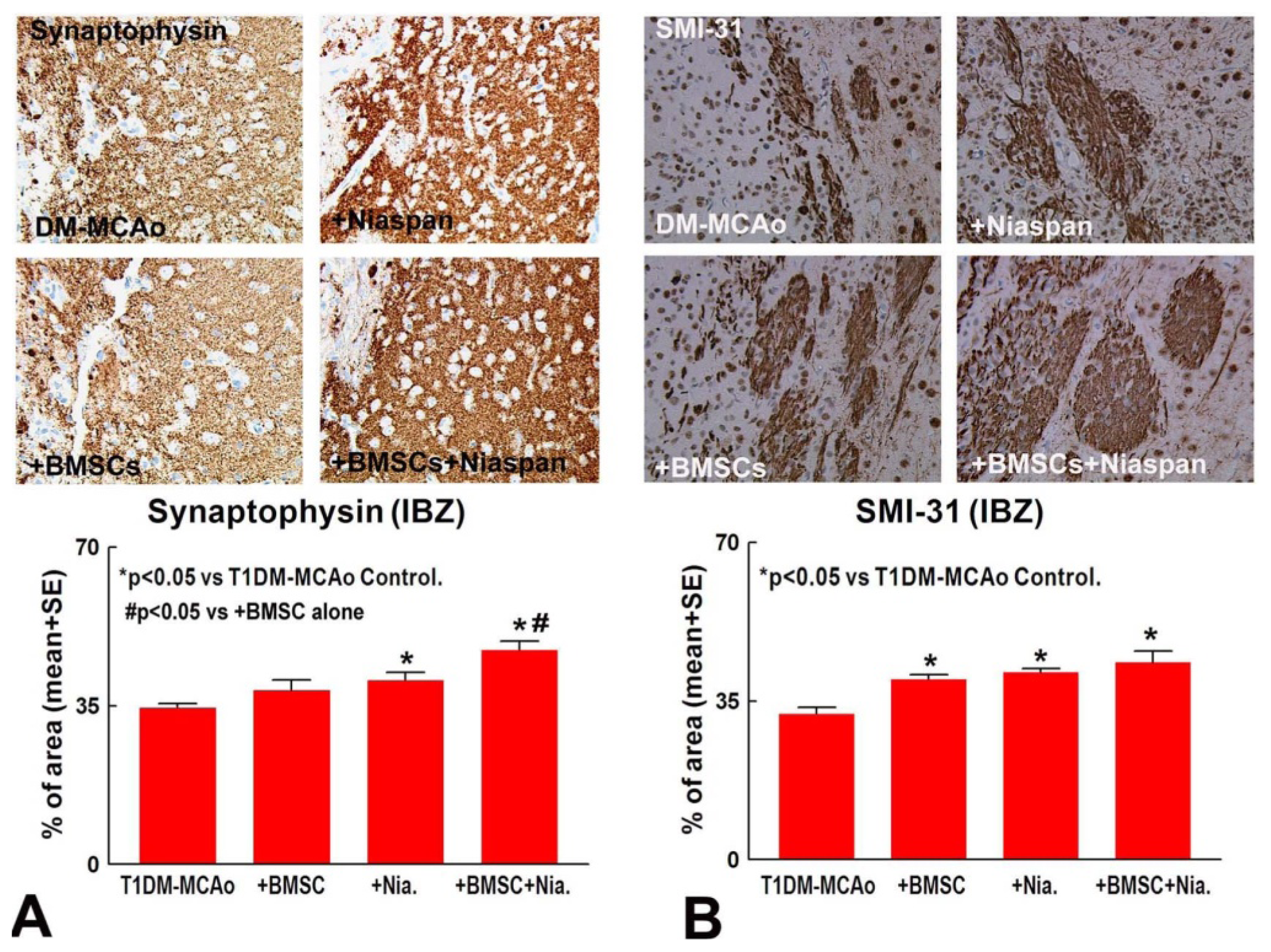
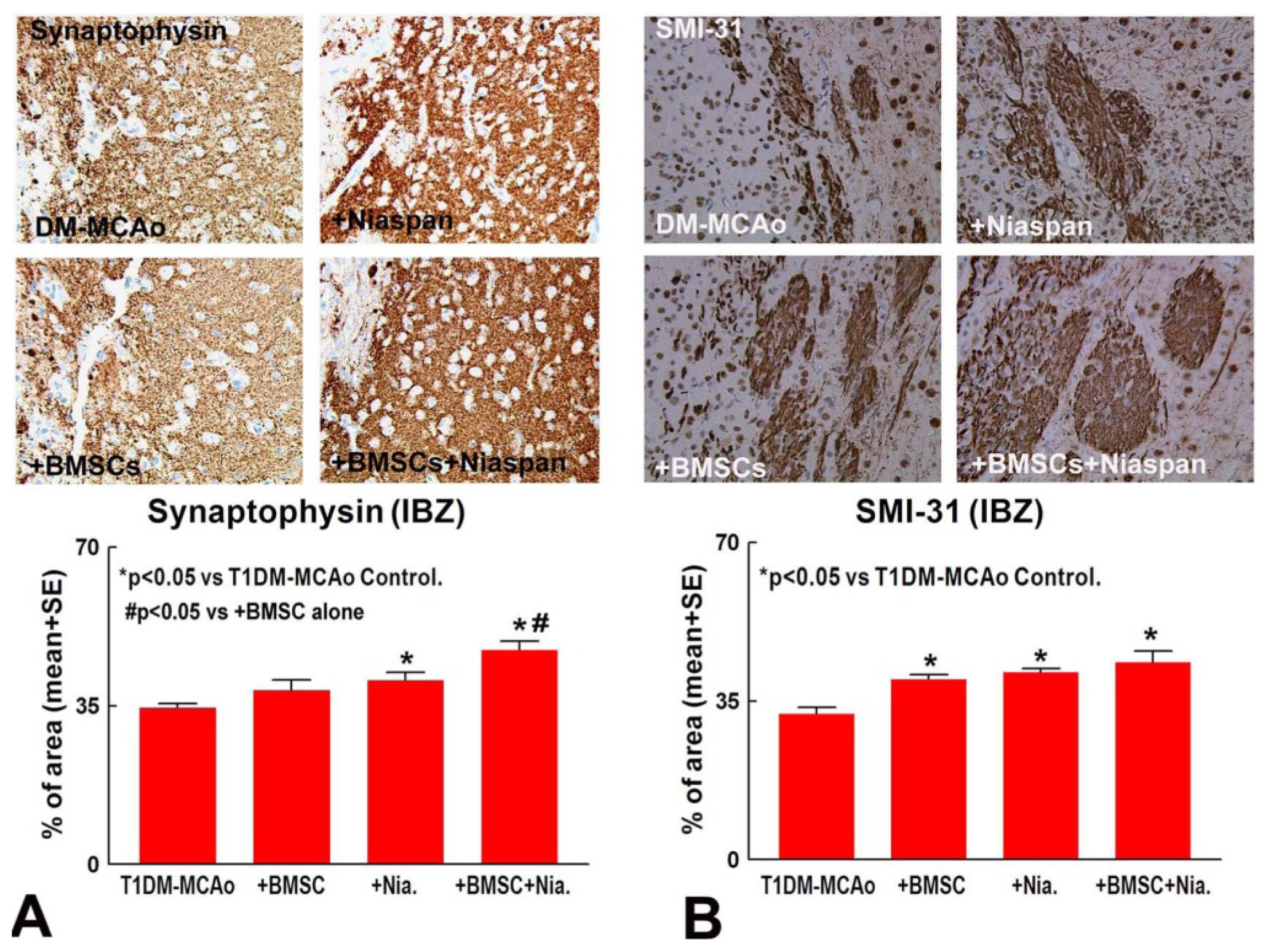
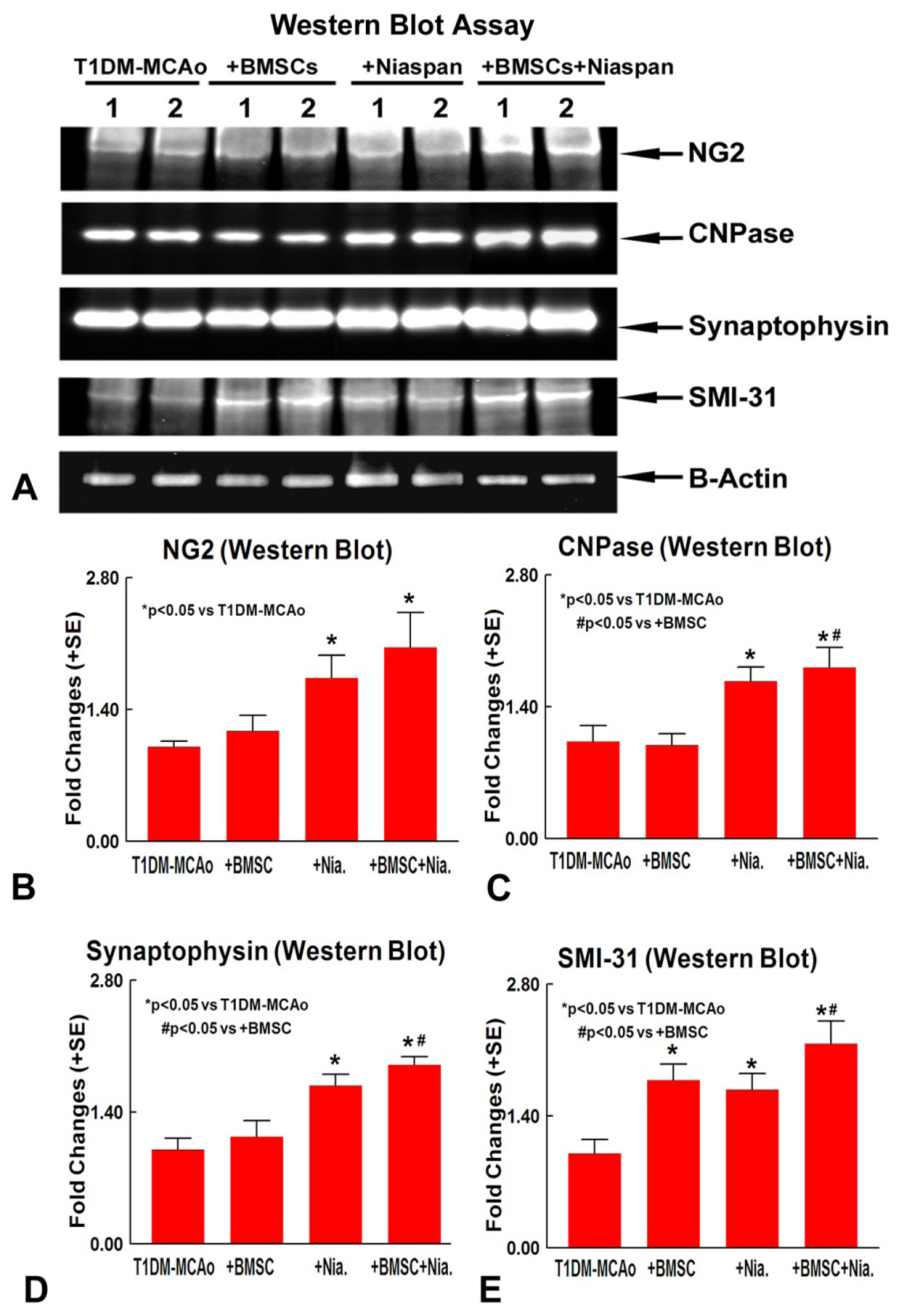
2.2. Combination BMSCs and Niaspan Treatment of Stroke Additively Enhances Synaptic Protein Expression
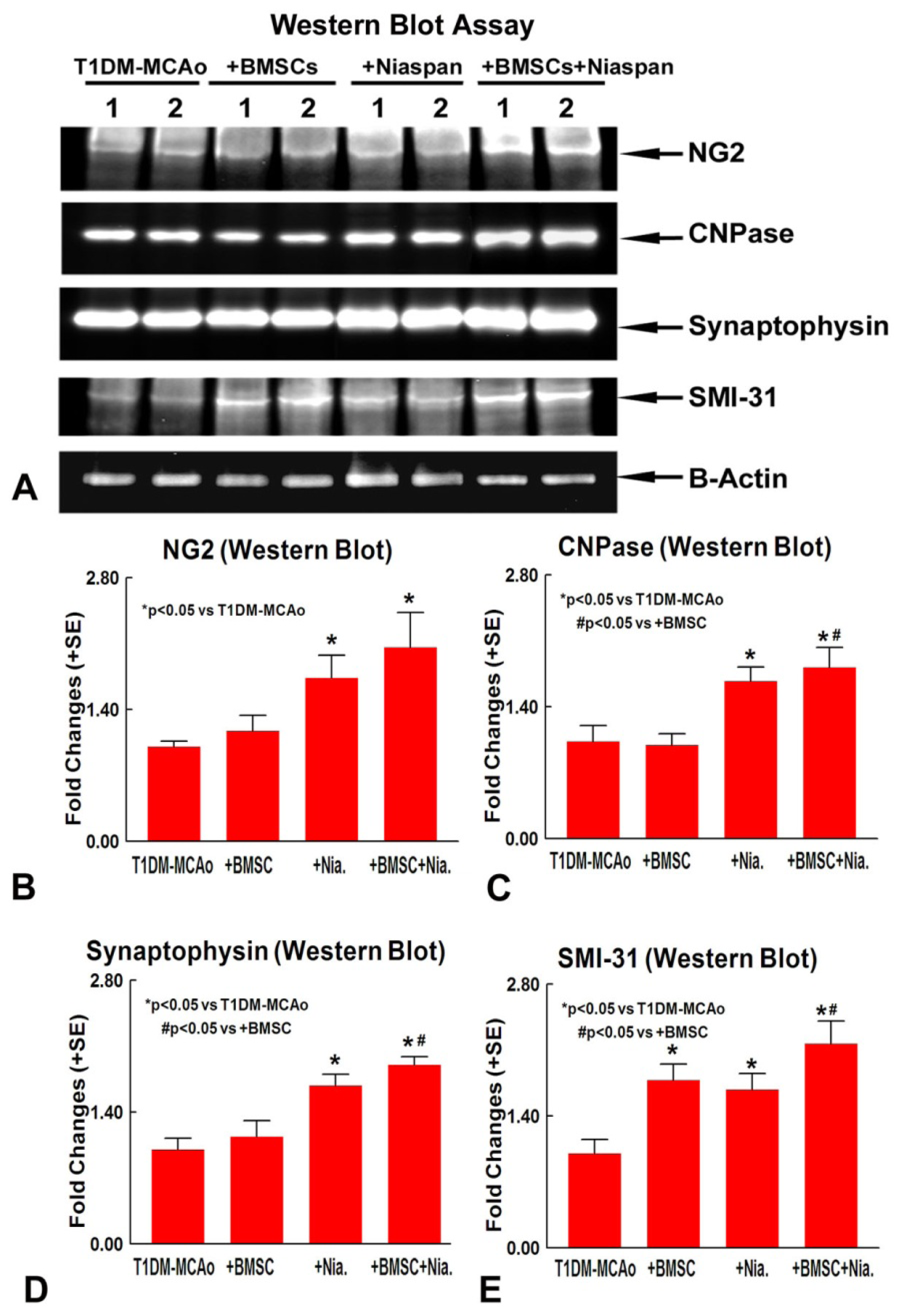
2.3. Combination BMSCs and Niaspan Treatment of Stroke Increases CNPase, Synaptophysin, and SMI-31 Expression in the Ischemic Brain
3. Discussion
4. Experimental Section
4.1. Diabetes Induction
4.2. BMSCs Culture
4.3. MCAo Model and Experiment Groups
4.4. Immunohistochemical Assessment
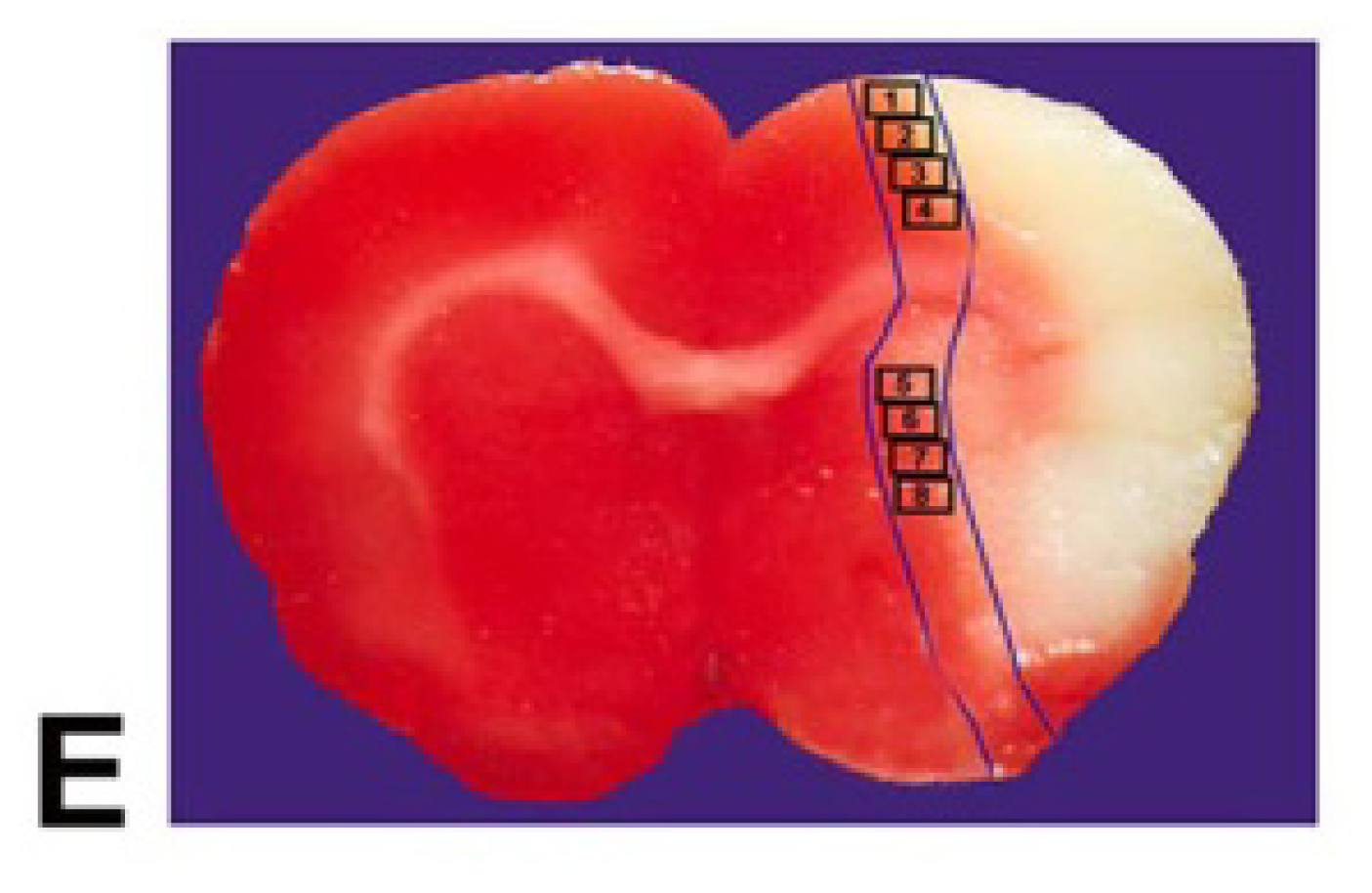
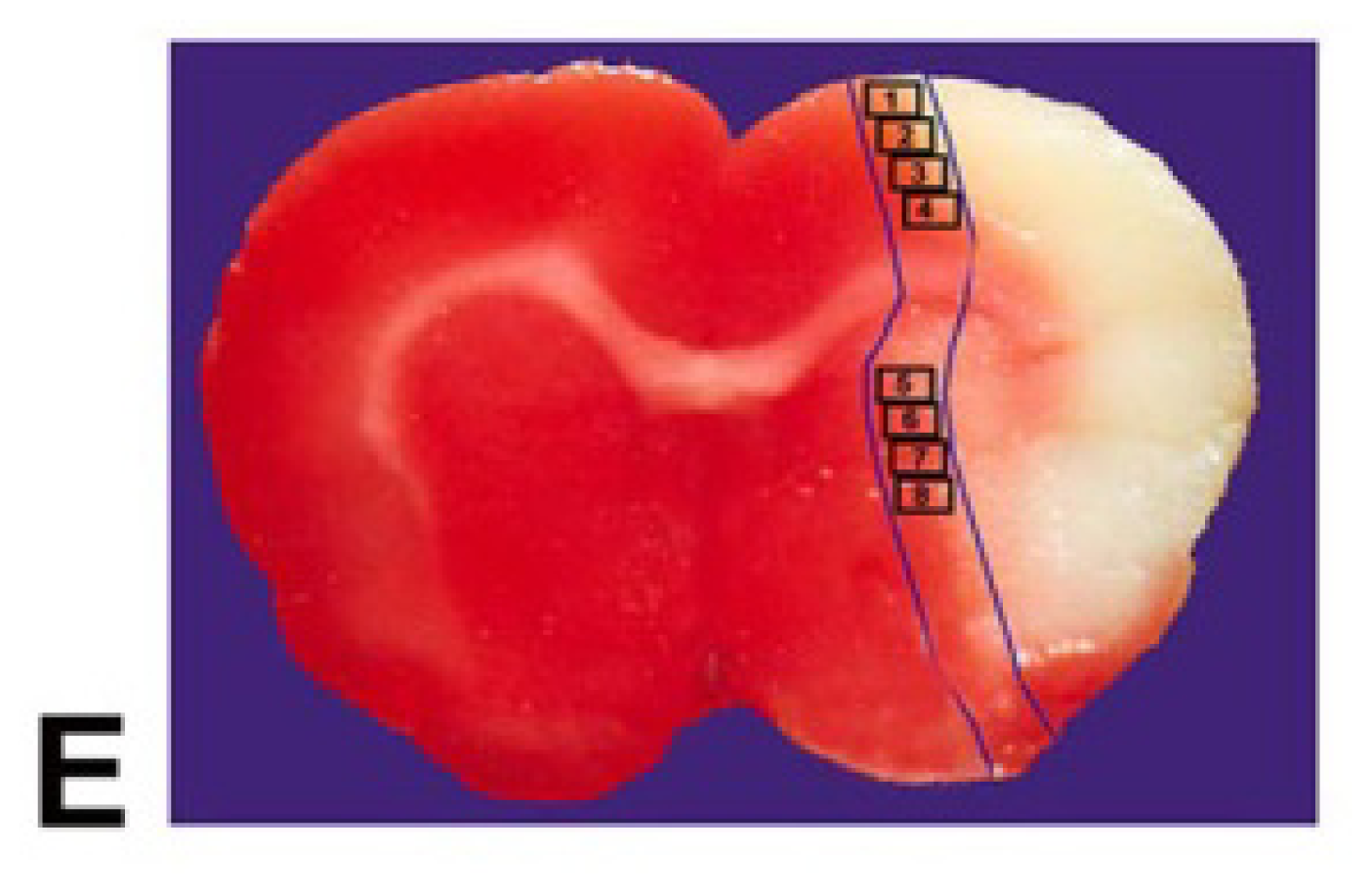
4.5. Quantitation Measurements of NG2, CNPase, Synaptophysin, BS, and LFB
4.6. Western Blot Assay
4.7. Statistical Analysis
5. Conclusions
Figures
Acknowledgments
Conflicts of Interest
References
- Go, A.S.; Mozaffarian, D.; Roger, V.L.; Benjamin, E.J.; Berry, J.D.; Borden, W.B.; Bravata, D.M.; Dai, S.; Ford, E.S.; Fox, C.S.; et al. Heart disease and stroke statistics—2013 update: A report from the American Heart Association. Circulation 2013, 127, e6–e245. [Google Scholar]
- Idris, I.; Thomson, G.A.; Sharma, J.C. Diabetes mellitus and stroke. Int. J. Clin. Pract 2006, 60, 48–56. [Google Scholar]
- Megherbi, S.-E.; Milan, C.; Minier, D.; Couvreur, G.; Osseby, G.-V.; Tilling, K.; Di Carlo, A.; Inzitari, D.; Wolfe, C.D.A.; Moreau, T.; et al. Association between diabetes and stroke subtype on survival and functional outcome 3 months after stroke: Data from the european biomed stroke project. Stroke 2003, 34, 688–694. [Google Scholar]
- Capes, S.E.; Hunt, D.; Malmberg, K.; Pathak, P.; Gerstein, H.C. Stress hyperglycemia and prognosis of stroke in nondiabetic and diabetic patients: A systematic overview. Stroke 2001, 32, 2426–2432. [Google Scholar]
- Cui, G.; Ye, X.; Zuo, T.; Zhao, H.; Zhao, Q.; Chen, W.; Hua, F. Chloroquine pretreatment inhibits toll-like receptor 3 signaling after stroke. Neurosci. Lett 2013, 548, 101–104. [Google Scholar]
- Sacco, R.L.; Wolf, P.A.; Gorelick, P.B. Risk factors and their management for stroke prevention: outlook for 1999 and beyond. Neurology 1999, 53, S15–S24. [Google Scholar]
- Yong, M.; Kaste, M. Dynamic of hyperglycemia as a predictor of stroke outcome in the ecass-II trial. Stroke 2008, 39, 2749–2755. [Google Scholar]
- Chen, J.; Li, Y.; Wang, L.; Zhang, Z.; Lu, D.; Lu, M.; Chopp, M. Therapeutic benefit of intravenous administration of bone marrow stromal cells after cerebral ischemia in rats. Stroke 2001, 32, 1005–1011. [Google Scholar]
- Chen, J.; Zhang, Z.G.; Li, Y.; Wang, L.; Xu, Y.X.; Gautam, S.C.; Lu, M.; Zhu, Z.; Chopp, M. Intravenous administration of human bone marrow stromal cells induces angiogenesis in the ischemic boundary zone after stroke in rats. Circ. Res 2003, 92, 692–699. [Google Scholar]
- Hamano, K.; Li, T.S.; Kobayashi, T.; Kobayashi, S.; Matsuzaki, M.; Esato, K. Angiogenesis induced by the implantation of self-bone marrow cells: A new material for therapeutic angiogenesis. Cell Transplant 2000, 9, 439–443. [Google Scholar]
- Villars, F.; Bordenave, L.; Bareille, R.; Amedee, J. Effect of human endothelial cells on human bone marrow stromal cell phenotype: role of VEGF? J. Cell Biochem 2000, 79, 672–685. [Google Scholar]
- Zacharek, A.; Chen, J.; Cui, X.; Li, A.; Li, Y.; Roberts, C.; Feng, Y.; Gao, Q.; Chopp, M. Angiopoietin1/Tie2 and VEGF/Flk1 induced by MSC treatment amplifies angiogenesis and vascular stabilization after stroke. J. Cereb. Blood Flow Metab 2007, 27, 1684–1691. [Google Scholar]
- Li, Y.; Chen, J.; Wang, L.; Lu, M.; Chopp, M. Treatment of stroke in rat with intracarotid administration of marrow stromal cells. Neurology 2001, 56, 1666–1672. [Google Scholar]
- Lu, D.; Mahmood, A.; Wang, L.; Li, Y.; Lu, M.; Chopp, M. Adult bone marrow stromal cells administered intravenously to rats after traumatic brain injury migrate into brain and improve neurological outcome. Neuroreport 2001, 12, 559–563. [Google Scholar]
- Elam, M.B.; Hunninghake, D.B.; Davis, K.B.; Garg, R.; Johnson, C.; Egan, D.; Kostis, J.B.; Sheps, D.S.; Brinton, E.A. Effect of niacin on lipid and lipoprotein levels and glycemic control in patients with diabetes and peripheral arterial disease: The ADMIT study: A randomized trial. Arterial Disease Multiple Intervention Trial. JAMA 2000, 284, 1263–1270. [Google Scholar]
- Chen, J.; Cui, X.; Zacharek, A.; Jiang, H.; Roberts, C.; Zhang, C.; Lu, M.; Kapke, A.; Feldkamp, C.S.; Chopp, M. Niaspan increases angiogenesis and improves functional recovery after stroke. Ann. Neurol 2007, 62, 49–58. [Google Scholar]
- Meyers, C.D.; Kamanna, V.S.; Kashyap, M.L. Niacin therapy in atherosclerosis. Curr. Opin. Lipidol 2004, 15, 659–665. [Google Scholar]
- Ye, X.; Chopp, M.; Cui, X.; Zacharek, A.; Cui, Y.; Yan, T.; Shehadah, A.; Roberts, C.; Liu, X.; Lu, M.; et al. Niaspan enhances vascular remodeling after stroke in type 1 diabetic rats. Exp. Neurol 2011, 232, 299–308. [Google Scholar]
- Yan, T.; Chopp, M.; Ye, X.; Liu, Z.; Zacharek, A.; Cui, Y.; Roberts, C.; Buller, B.; Chen, J. Niaspan increases axonal remodeling after stroke in type 1 diabetes rats. Neurobiol. Dis 2012, 46, 157–164. [Google Scholar]
- Zhang, Z.G.; Chopp, M. Neurorestorative therapies for stroke: Underlying mechanisms and translation to the clinic. Lancet Neurol 2009, 8, 491–500. [Google Scholar]
- Cho, S.; Kim, E.; Qin, L.; Bhosle, S.; Bao, Y.; Alam, N.; Prusky, G.; Ratan, R. Daidzein promotes functional recovery and enhances glial-directed synaptogenesis in an experimental animal model of stroke. Stroke 2010, 41, E210. [Google Scholar]
- Tanaka, K.; Nogawa, S.; Suzuki, S.; Dembo, T.; Kosakai, A. Upregulation of oligodendrocyte progenitor cells associated with restoration of mature oligodendrocytes and myelination in peri-infarct area in the rat brain. Brain Res 2003, 989, 172–179. [Google Scholar]
- Yan, T.; Ye, X.; Chopp, M.; Zacharek, A.; Ning, R.; Venkat, P.; Roberts, C.; Lu, M.; Chen, J. Niaspan attenuates the adverse effects of bone marrow stromal cell treatment of stroke in type one diabetic rats. PLoS One 2013, in press. [Google Scholar]
- Dewar, D.; Underhill, S.M.; Goldberg, M.P. Oligodendrocytes and ischemic brain injury. J. Cereb. Blood Flow Metab 2003, 23, 263–274. [Google Scholar]
- Zhang, L.; Chopp, M.; Zhang, R.L.; Wang, L.; Zhang, J.; Wang, Y.; Toh, Y.; Santra, M.; Lu, M.; Zhang, Z.G. Erythropoietin amplifies stroke-induced oligodendrogenesis in the rat. PLoS One 2010, 5, e11016. [Google Scholar]
- Arai, K.; Lo, E.H. Oligovascular signaling in white matter stroke. Biol. Pharm. Bull 2009, 32, 1639–1644. [Google Scholar]
- Nowak, D.A.; Grefkes, C.; Ameli, M.; Fink, G.R. Interhemispheric Competition After Stroke: Brain Stimulation to Enhance Recovery of Function of the Affected Hand. Neurorehabil. Neural Repair 2009, 23, 641–656. [Google Scholar]
- Weidner, N.; Ner, A.; Salimi, N.; Tuszynski, M.H. Spontaneous corticospinal axonal plasticity and functional recovery after adult central nervous system injury. Proc. Natl. Acad. Sci. USA 2001, 98, 3513–3518. [Google Scholar]
- Font, M.A.; Arboix, A.; Krupinski, J. Angiogenesis, neurogenesis and neuroplasticity in ischemic stroke. Current Cardiol. Rev 2010, 6, 238–244. [Google Scholar]
- Calabresi, P.; Centonze, D.; Pisani, A.; Cupini, L.M.; Bernardi, G. Synaptic plasticity in the ischaemic brain. Lancet Neurol 2003, 2, 622–629. [Google Scholar]
- Ye, X.; Chopp, M.; Liu, X.; Zacharek, A.; Cui, X.; Yan, T.; Roberts, C.; Chen, J. Niaspan reduces high-mobility group box 1/receptor for advanced glycation endproducts after stroke in type-1 diabetic rats. Neuroscience 2011, 190, 339–345. [Google Scholar]
- Ding, J.; Cheng, Y.; Gao, S.; Chen, J. Effects of nerve growth factor and Noggin-modified bone marrow stromal cells on stroke in rats. J. Neurosci. Res 2011, 89, 222–230. [Google Scholar]
- Irving, E.A.; Bentley, D.L.; Parsons, A.A. Assessment of white matter injury following prolonged focal cerebral ischaemia in the rat. Acta Neuropathol 2001, 102, 627–635. [Google Scholar]
- Cui, G.; Zuo, T.; Zhao, Q.; Hu, J.; Jin, P.; Zhao, H.; Jing, J.; Zhu, J.; Chen, H.; Liu, B.; et al. ROCK mediates the inflammatory response in thrombin induced microglia. Neurosci. Lett 2013, 554, 82–87. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ye, X.; Yan, T.; Chopp, M.; Zacharek, A.; Ning, R.; Venkat, P.; Roberts, C.; Chen, J. Combination BMSC and Niaspan Treatment of Stroke Enhances White Matter Remodeling and Synaptic Protein Expression in Diabetic Rats. Int. J. Mol. Sci. 2013, 14, 22221-22232. https://doi.org/10.3390/ijms141122221
Ye X, Yan T, Chopp M, Zacharek A, Ning R, Venkat P, Roberts C, Chen J. Combination BMSC and Niaspan Treatment of Stroke Enhances White Matter Remodeling and Synaptic Protein Expression in Diabetic Rats. International Journal of Molecular Sciences. 2013; 14(11):22221-22232. https://doi.org/10.3390/ijms141122221
Chicago/Turabian StyleYe, Xinchun, Tao Yan, Michael Chopp, Alex Zacharek, Ruizhuo Ning, Poornima Venkat, Cynthia Roberts, and Jieli Chen. 2013. "Combination BMSC and Niaspan Treatment of Stroke Enhances White Matter Remodeling and Synaptic Protein Expression in Diabetic Rats" International Journal of Molecular Sciences 14, no. 11: 22221-22232. https://doi.org/10.3390/ijms141122221