Differential Proteomic Analysis of Anthers between Cytoplasmic Male Sterile and Maintainer Lines in Capsicum annuum L.

Abstract
:
1. Introduction
2. Results
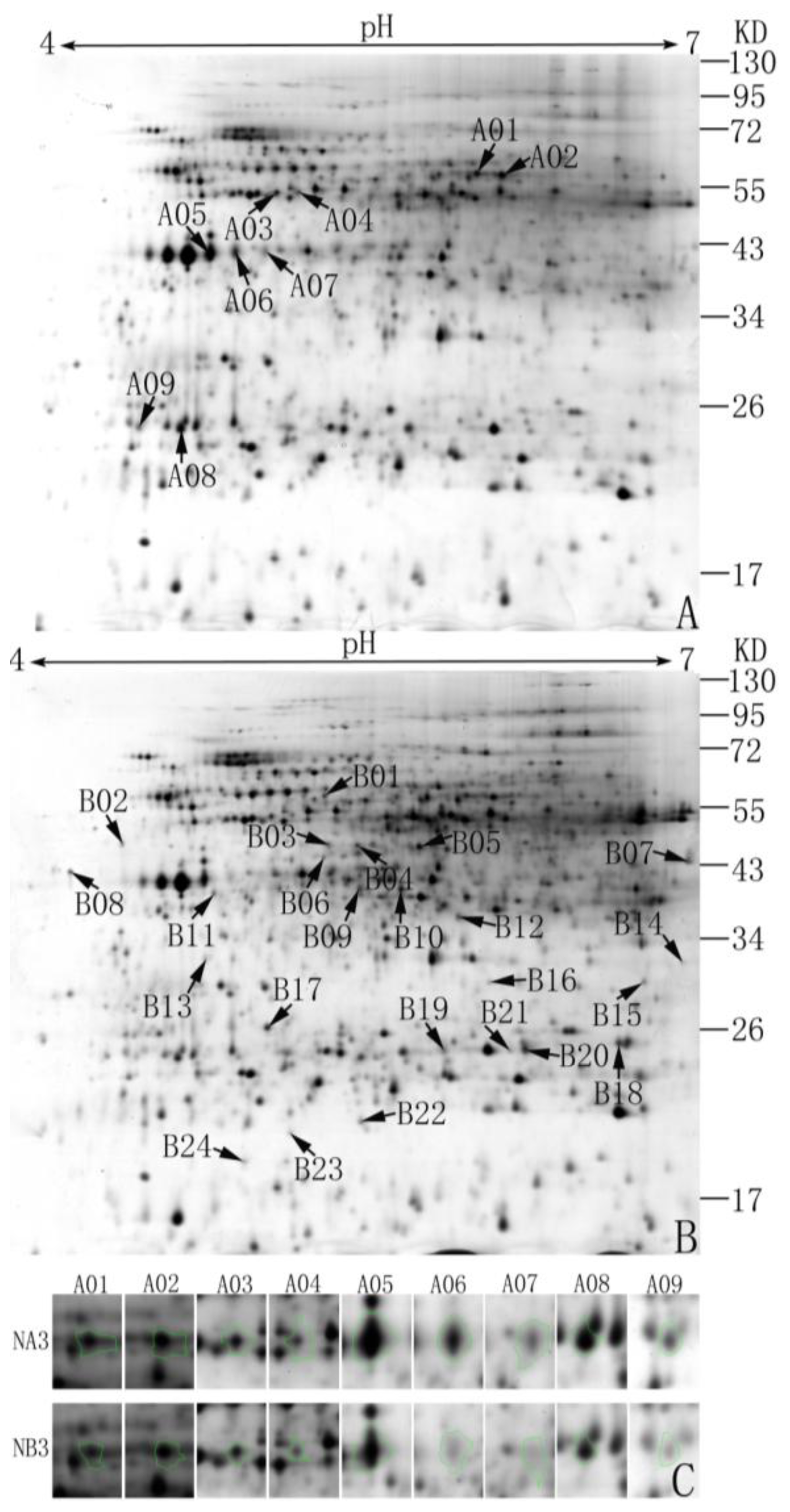
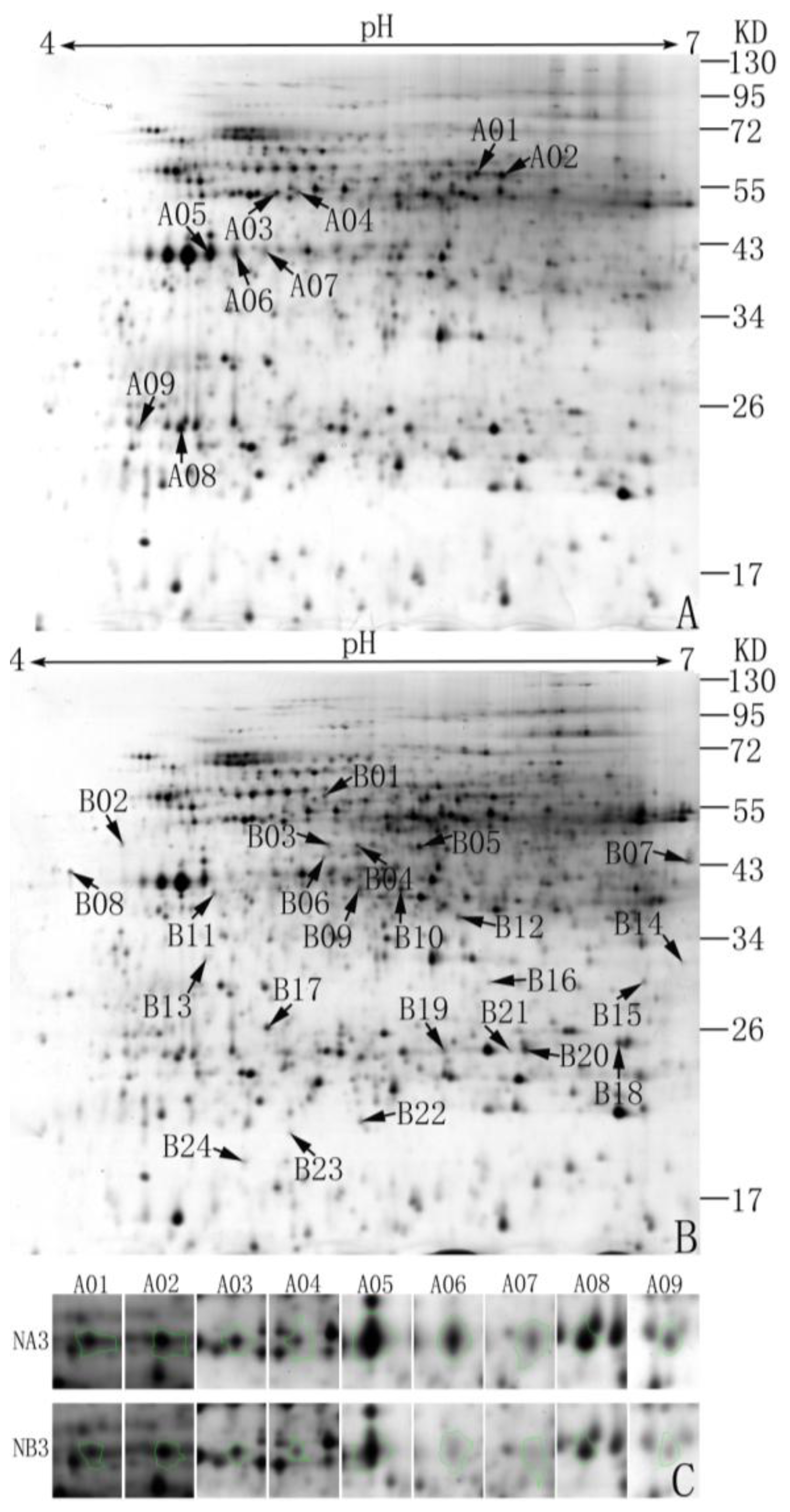
2.1. Abundance Analysis of Differentially Expressed Proteins in the Normal and Sterile Cytoplasm
2.2. Identification and Functional Classification of Differentially Expressed Proteins
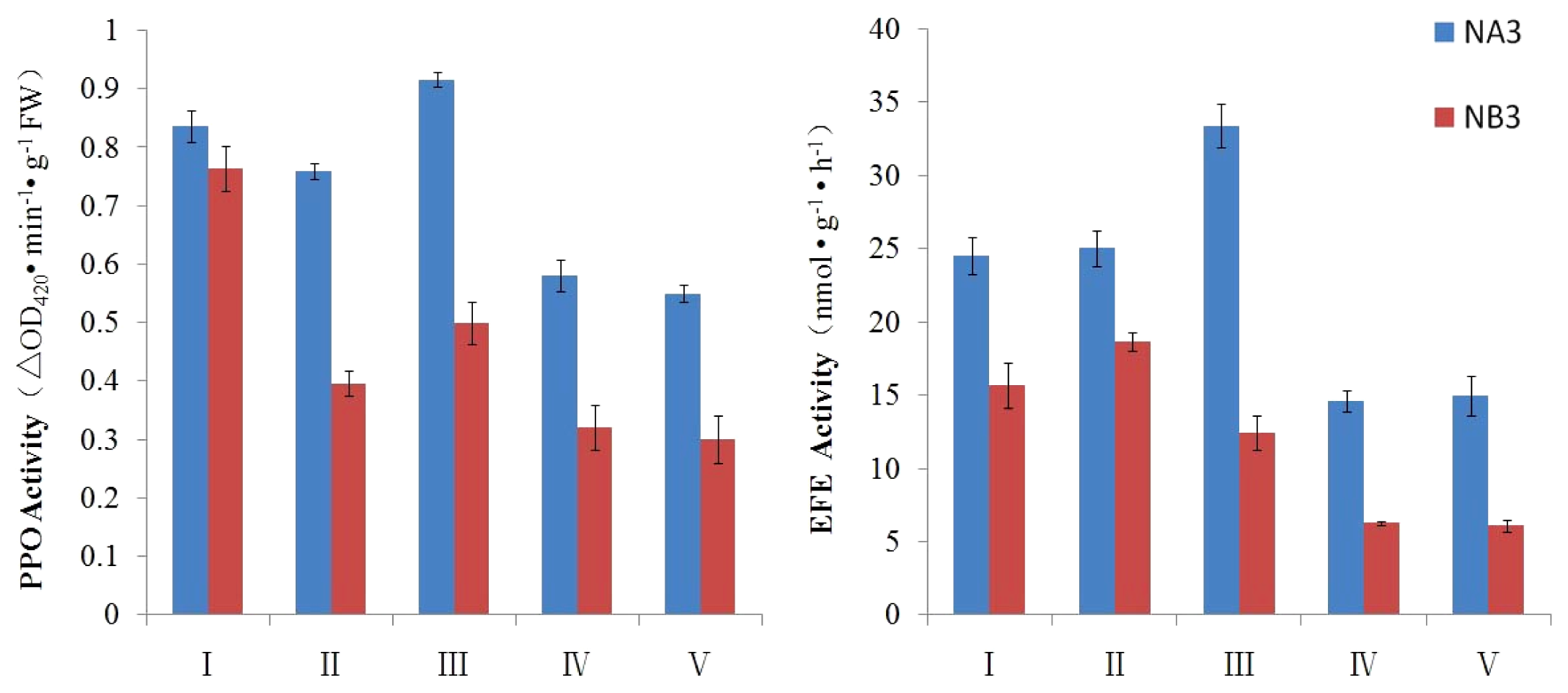
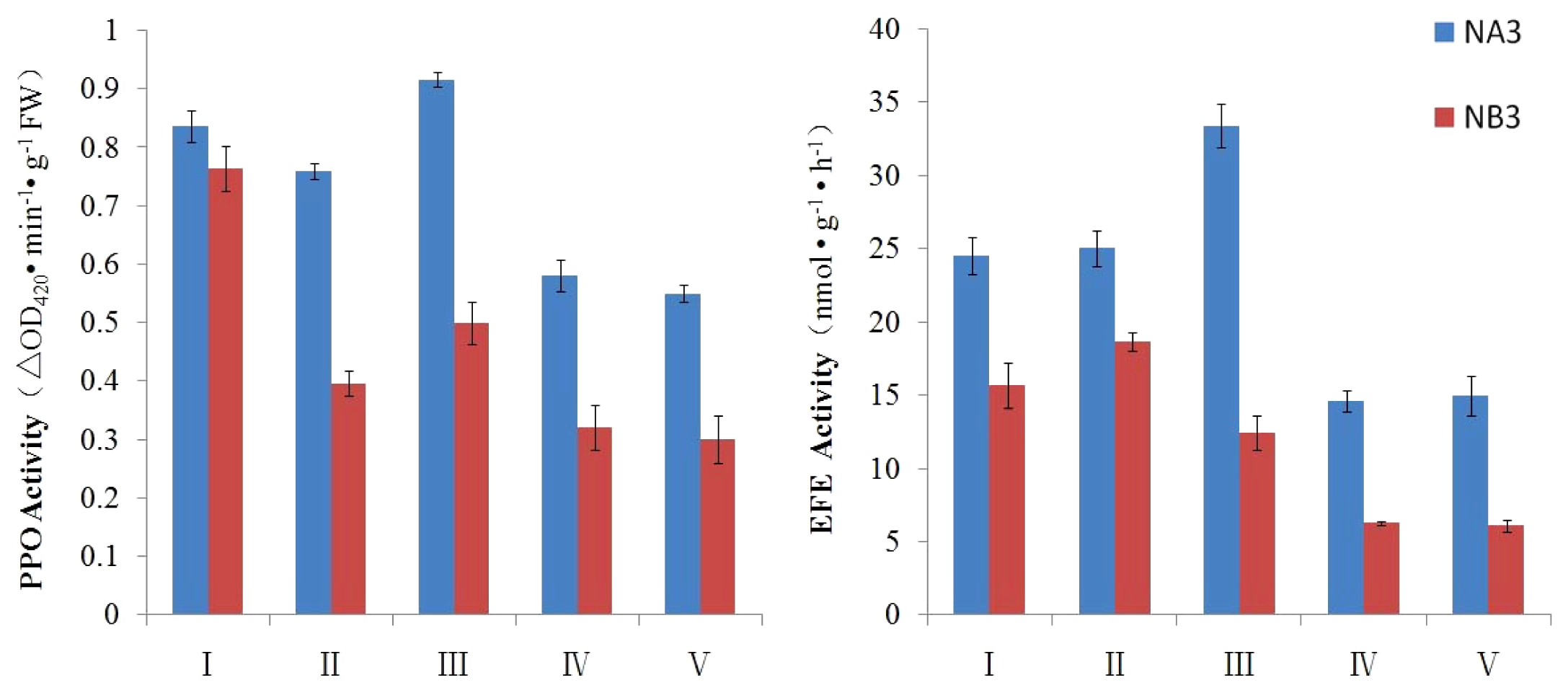
2.3. Evaluation of PPO and EFE Activities
3. Discussion
3.1. Proteins Involved in Respiration and Energy Pathway
3.2. Proteins Involved in Carbohydrate Metabolic Pathways
3.3. Proteins Involved in Photosynthesis
3.4. Proteins Involved in Antioxidative Reactions
3.5. Proteins Involved in Amino Acid Metabolism
4. Experimental Section
4.1. Plant Materials and Anther Collection
4.2. Protein Extraction and 2-DE Electrophoresis
4.3. Gel Imaging and Analysis
4.4. Trypsindigestion, Mass Spectrometry, and Protein Identification
4.5. PPO and EFE Activity Assay
5. Conclusions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. a | Protein name (Species) b | Accession No. c | kDa/pI (Theor d) | kDa/pI (Exper e) | Mascot score | Peptides matched | S.C. (%) f | A/Bspot volume ratio g |
|---|---|---|---|---|---|---|---|---|
| Respiration and energy pathway (6) | ||||||||
| A03 | ATP synthase subunit beta (NP) | gi|114421 | 59.93/5.95 | 53.95/5.10 | 370 | 13 | 33 | +1.69 |
| A04 | ATP synthase subunit beta (NP) | gi|114421 | 59.93/5.95 | 54.05/5.18 | 362 | 13 | 35 | +3.09 |
| B03 | ATP synthase CF1 alpha chain (SL) | gi|89280620 | 55.43/5.14 | 48.34/5.30 | 453 | 16 | 30 | −∞ |
| B04 | ATP synthase CF1 alpha subunit (NS) | gi|78102516 | 55.44/5.14 | 47.25/5.43 | 549 | 15 | 27 | −∞ |
| B24 | ATP synthase D chain (SD) | gi|48209968 | 19.80/5.34 | 19.21/5.00 | 266 | 13 | 55 | −13.53 |
| B07 | Formate dehydrogenase, mitochondrial (ST) | gi|26454627 | 42.30/6.64 | 34.00/6.50 | 202 | 10 | 33 | −1.73 |
| Carbohydrate metabolism (3) | ||||||||
| B05 | alpha-mannosidase (CA) | gi|315440801 | 47.03/6.28 | 47.09/5.67 | 284 | 14 | 12 | −4.01 |
| B19 | triose phosphate isomerase cytosolic isoform (SC) | gi|38112662 | 27.25/5.13 | 24.97/5.74 | 279 | 6 | 21 | −∞ |
| B21 | triose phosphate isomerase cytosolic isoform (SC) | gi|38112662 | 27.25/5.73 | 24.74/5.98 | 358 | 5 | 27 | −1.60 |
| Photosynthesis (3) | ||||||||
| B01 | RuBisCO large subunit-binding protein subunit beta, chloroplastic RuBisCO (VV) | gi|225442531 | 65.26/5.62 | 58.16/5.30 | 128 | 4 | 8 | −1.60 |
| B13 | chloroplast manganese stabilizing protein-II (ST) | gi|239911810 | 31.44/6.45 | 32.74/5.00 | 284 | 4 | 16 | −1.61 |
| B23 | 23 kDa polypeptide of the oxygen evolving complex of photosystem II (SA) | gi|241865142 | 22.91/5.14 | 20.41/5.16 | 131 | 2 | 9 | −∞ |
| Antioxidative reactions(3) | ||||||||
| A01 | Polyphenol oxidase F (SL) | gi|1172583 | 66.77/6.27 | 58.38/5.85 | 218 | 6 | 8 | +1.76 |
| A02 | Polyphenol oxidase F (SL) | gi|1172583 | 66.77/6.27 | 57.93/5.94 | 212 | 8 | 10 | +1.53 |
| B20 | glutathione S-transferase (AH) | gi|254798560 | 24.86/6.96 | 24.67/6.03 | 83 | 2 | 9 | −2.76 |
| Amino acid metabolism (3) | ||||||||
| B09 | glutamine synthetase(SL) | gi|541632 | 18.74/6.12 | 34.00/5.44 | 274 | 4 | 15 | −∞ |
| B10 | glutamine synthetase (AM) | gi|12802875 | 39.43/6.17 | 34.00/5.59 | 305 | 6 | 22 | −∞ |
| B12 | glutamine synthetase (SL) | gi|541632 | 18.74/6.12 | 34.00/5.80 | 128 | 5 | 15 | −∞ |
| Signal transduction (2) | ||||||||
| B11 | adenosine kinase isoform 1T-like protein (ST) | gi|82400168 | 39.75/5.01 | 34.00/5.00 | 81 | 3 | 10 | −2.89 |
| B16 | annexin Cap32(CA) | gi|3979715 | 35.95/5.85 | 29.85/5.91 | 530 | 20 | 55 | −∞ |
| Cytoskeleton (2) | ||||||||
| A07 | Actin(NA) | gi|378724806 | 27.95/5.20 | 34.00/5.05 | 279 | 8 | 44 | +2.27 |
| B06 | Actin (NA) | gi|378724806 | 27.95/5.20 | 47.00/5.29 | 118 | 8 | 44 | −∞ |
| Storage protein (2) | ||||||||
| B14 | Glutelin (OS) | gi|225710 | 56.74/8.93 | 31.20/6.50 | 92 | 5 | 10 | −6.47 |
| B15 | Glutelin (OS) | gi|20217 | 56.45/9.17 | 29.23/6.50 | 158 | 6 | 16 | −∞ |
| Others (3) | ||||||||
| B02 | putative DNA repair protein RAD23-4 (AT) | gi|145334669 | 34.81/4.85 | 48.84/5.00 | 83 | 2 | 5 | −2.22 |
| B17 | putative caffeoyl-CoA 3-O-methyltransferase(CA) | gi|193290676 | 27.92/5.30 | 26.07/5.07 | 231 | 9 | 36 | −2.84 |
| B22 | allene oxide cyclase (NT) | gi|40644130 | 26.68/6.07 | 20.86/5.42 | 88 | 2 | 7 | −∞ |
Acknowledgments
Conflicts of Interest
References
- Guo, J.Z.; Ma, J.H. Determination of the combining ability of several quantitative traits constituting yield. China Veg 1984, 3, 1–4. [Google Scholar]
- Hanson, M.R.; Bentolila, S. Interaction of mitochondria and nuclear genes that affect male gametophyte development. Plant Cell 2004, 16, S154–S169. [Google Scholar]
- Schnable, P.S.; Wise, R.P. The molecular basis of cytoplasmic male sterility and fertility restoration. Trends Plant Sci 1998, 3, 175–180. [Google Scholar]
- Wang, Z.H.; Zou, Y.J.; Li, X.; Zhang, Q.; Chen, L.; Wu, H.; Su, D.; Chen, Y.; Guo, J.; Luo, D.; et al. Cytoplasmic male sterility of rice with boro II cytoplasm is caused by a cytotoxic peptide and is restored by two related PPR motif genes via distinct modes of mRNA silencing. Plant Cell 2006, 18, 676–687. [Google Scholar]
- Hu, J.; Wang, K.; Huang, W.; Liu, G.; Gao, Y.; Wang, J.; Huang, Q.; Ji, Y.; Qin, X.; Wan, L.; et al. The rice pentatricopeptide repeat protein RF 5 restores fertility in Hong-Lian cytoplasmic male-sterile lines via a complex with the glycine-rich protein GRP162. Plant Cell 2012, 24, 109–122. [Google Scholar]
- Luo, D.P.; Xu, H.; Liu, Z.L.; Guo, J.X.; Li, H.Y.; Chen, L.T.; Fang, C.; Zhang, Q.Y.; Bai, M.; Yao, N.; et al. A detrimental mitochondrial-nuclear interaction causes cytoplasmic male sterility in rice. Nat. Genet 2013, 45, 573–577. [Google Scholar]
- Martin, J.; Grawford, J.H. Several types of sterility in Capsicum frutescens. Proc. Am. Soc. Hortic. Sci 1951, 57, 335–338. [Google Scholar]
- Peterson, P.A. Cytoplasmically inherited male sterility in Capsicum. Am. Nat 1958, 92, 111–119. [Google Scholar]
- Kim, D.H.; Kang, J.G.; Kim, B.D. Isolation and characterization of the cytoplasmic male sterility-associated orf456 gene of chili pepper (Capsicum annuum L.). Plant Mol. Biol 2007, 63, 519–532. [Google Scholar]
- Gulyas, G.; Shin, Y.; Kim, H.; Lee, J.S.; Hirata, Y. Altered transcript reveals an orf507 sterility-related gene in chili pepper (Capsicum annuum L.). Plant Mol. Biol. Rep 2010, 28, 605–612. [Google Scholar]
- Wen, L.; Liu, G.; Li, S.Q.; Wan, C.X.; Tao, J.; Xu, K.Y.; Zhang, Z.J.; Zhu, Y.G. Proteomic analysis of anthers from Honglian cytoplasmic male sterility line rice and its corresponding maintainer and hybrid. Bot. Stud 2007, 48, 293–309. [Google Scholar]
- Sheoran, I.S.; Ross, A.R.; Olson, D.J.; Sawhney, V.K. Differential expression of proteins in the wild type and 7B-1 male-sterile mutant anthers of tomato (Solanum lycopersicum): A proteomic analysis. J. Proteomics 2009, 71, 624–636. [Google Scholar]
- Sheoran, I.S.; Sawhneya, V.K. Proteome analysis of the normal and Ogura (ogu) CMS anthers of Brassica napus to identify proteins associated with male sterility. Botany 2010, 88, 217–230. [Google Scholar]
- Zheng, R.; Yue, S.; Xu, X.; Liu, J.; Xu, Q.; Wang, X.L.; Han, L.; Yu, D.Y. Proteome analysis of the wild and YX-1 male sterile mutant anthers of wolfberry (Lycium barbarum L.). PLoS One 2012, 7, e41861. [Google Scholar]
- Qi, J.M.; Ma, H.B.; Xu, J.T.; Chen, M.X.; Zhou, D.X.; Wang, T.; Chen, S.H. Proteomic analysis of bud differentiation between cytoplasmic male-sterile line and maintainer in tobacco. Acta Agron. Sin 2012, 38, 1232–1239. [Google Scholar]
- Lee, S.J.; Warmke, H.E. Organelle size and number in fertile and T-cytoplasmic male-sterile corn. Am. J. Bot 1979, 66, 141–148. [Google Scholar]
- Yui, R.; Iketani, S.; Mikami, T.; Kubo, T. Antisense inhibition of mitochondrial pyruvate dehydrogenase subunit in anther tapetum causes male sterility. Plant J 2003, 34, 57–66. [Google Scholar]
- Siedow, J.N.; Umbach, A.L. Plant mitochondrial electron transfer and molecular biology. Plant Cell 1995, 7, 821–831. [Google Scholar]
- Xu, K.; Cao, M.J.; Zhu, Y.G.; Pan, G.T.; Rong, T.Z. Differential expression of mitochondrial proteins between C-type cytoplasmic male sterility line C48-2 and its maintainer line in maize. Acta Agron. Sin 2008, 2, 232–237. [Google Scholar]
- Sabar, M.; Gagliardi, D. ORFB is a subunit of F1F0-ATPsynthase: In sight into the basis of cytoplasmic male sterility in sunflower. EMBO Rep 2003, 4, 381–386. [Google Scholar]
- Zhang, H.; Li, S.Q.; Yi, P.; Wan, C.X. A honglian CMS line of rice displays aberrant F0 of F1F0-ATPase. Plant Cell Rep 2007, 26, 1065–1071. [Google Scholar]
- Ji, J.J.; Huang, W.; Yin, C.C.; Gong, Z.H. Mitochondrial cytochrome c oxidase and F1F0-ATPase dysfunction in peppers (Capsicum annuum L.) with cytoplasmic male sterility and its association with orf507 and Ψatp6-2 genes. Int. J. Mol. Sci 2013, 14, 1050–1068. [Google Scholar]
- Li, J.J.; Pandeya, D.; Jo, Y.D.; Liu, W.Y.; Kang, B.C. Reduced activity of ATP synthase in mitochondria causes cytoplasmic male sterility in chili pepper. Planta 2013, 237, 1097–1109. [Google Scholar]
- Lolis, E.; Alber, T.; Davenport, R.C.; Rose, D.; Hartman, F.C.; Petsko, G.A. Structure of yeast triosephosphate isomerase at 1.9-A resolution. Biochemistry 1990, 29, 6609–6618. [Google Scholar]
- Deng, M.H.; Wen, J.F.; Huo, J.L.; Zhu, H.S.; Dai, X.Z.; Zhang, Z.Q.; Zhou, H.; Zou, X.X. Relationship of metabolism of reactive oxygen species with cytoplasmic male sterility in pepper (Capsicum annuum L.). Sci. Hortic 2012, 134, 232–236. [Google Scholar]
- Raghavendra, A.S.; Padmasree, K. Beneficial interactions of mitochondrial metabolism with photosynthetic carbon assimilation. Trends Plant Sci 2003, 8, 546–553. [Google Scholar]
- Gilmore, A.M.; Bjorkman, O. Temperature-sensitive coupling and uncoupling of ATPase-mediated, nonradiative energy dissipation-similarities between chloroplast sand leaves. Planta 1995, 197, 646–654. [Google Scholar]
- Kromer, S. Respiration during photosynthesis. Annu. Rev. Plant Physiol 1995, l46, 45–70. [Google Scholar]
- Sabar, M.; de Paepe, R.; de Kouchkovsky, Y. Complex I impairment, respiratory compensations, and photosynthetic decrease in nuclear and mitochondrial male sterile mutants of Nicotiana sylvestris. Plant Physiol 2000, 124, 1239–1250. [Google Scholar]
- Dutilleul, C.; Driscoll, S.; Cornic, G.; de Paepe, R.; Foyer, C.H.; Noctor, G. Functional mitochondrial complex I is required by tobacco leaves for optimal photosynthetic performance in photo respiratory conditions and during transients. Plant Physiol 2003, 131, 264–275. [Google Scholar]
- Vaughn, K.C.; Lax, A.R.; Duke, S.O. Polyphenol oxidase: The chloroplast oxidase with no established function. Physiol. Plant 1988, 72, 659–665. [Google Scholar]
- Tingey, S.V.; Tsai, F.Y.; Edwards, J.W.; Walker, E.L.; Coruzzi, G.M. Chloroplast and cytosolic glutamine synthetase are encoded by homologous nuclear genes which are differentially expressed in vivo. J. Biol. Chem. 1988, 263, 9651–9657. [Google Scholar]
- Moffatt, B.A.; Stevens, Y.Y.; Allen, M.S.; Snider, J.D.; Pereira, L.A.; Todorova, M.I.; Summers, P.S.; Weretilnyk, E.A.; Martin-McCaffrey, L.; Wagner, C. Adenosine kinase deficiency is associated with developmental abnormalities and reduced transmethylation. Plant Physiol 2002, 128, 812–821. [Google Scholar]
- Sun, Z.J.; Xu, J.Q.; Song, M.; Wei, J.G.; Tang, Q.L.; Wang, Z.M.; Wang, X.J. Molecular cloning and expression analysis of annexingene from cabbage. Acta Hortic. Sin 2012, 39, 1567–1574. [Google Scholar]
- Hause, B.; Stenzel, I.; Miersch, O.; Maucher, H.; Kramell, R.; Ziegler, J.; Wasternack, C. Tissue-specific oxylipin signature of tomato flowers: Allene oxide cyclase is highly expressed in distinct flower organs and vascular bundles. Plant J 2000, 24, 113–126. [Google Scholar]
- Browse, J. Jasmonate passes muster: A receptor and targets for the defense hormone. Annu. Rev. Plant Biol 2009, 60, 183–205. [Google Scholar]
- Stumpe, M.; Göbel, C.; Faltin, B.; Beike, A.K.; Hause, B.; Himmelsbach, K.; Bode, J.; Kramell, R.; Wasternack, C.; Frank, W.; et al. The moss Physcomitrella patens contains cyclopentenones but no jasmonates: Mutations in allene oxide cyclase lead to reduced fertility and altered sporophyte morphology. New Phytol 2010, 188, 740–749. [Google Scholar]
- Shen, H.L.; Jiang, J.Z.; Wang, Z.Y.; Geng, S.S. Studies on the breeding and inheritance of male sterile lines of pepper (Capsicum annuum). Acta Agric. Univ. Pekin 1994, 20, 25–30. [Google Scholar]
- Leja, M.; Mareczeka, A.; Benb, J. Antioxidant properties of two apple cultivars during long-term storage. Food Chem 2003, 80, 303–307. [Google Scholar]
- Wei, J.; Kenji, T. Post-harvest changes of EFE activity, ACC and polyamine contents in cortex and pith of pear fruits and its relation to ethylene production. Acta Phytophisiol. Sin 1995, 21, 136–142. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, Z.; Cheng, J.; Qin, C.; Hu, Z.; Yin, C.; Hu, K. Differential Proteomic Analysis of Anthers between Cytoplasmic Male Sterile and Maintainer Lines in Capsicum annuum L. Int. J. Mol. Sci. 2013, 14, 22982-22996. https://doi.org/10.3390/ijms141122982
Wu Z, Cheng J, Qin C, Hu Z, Yin C, Hu K. Differential Proteomic Analysis of Anthers between Cytoplasmic Male Sterile and Maintainer Lines in Capsicum annuum L. International Journal of Molecular Sciences. 2013; 14(11):22982-22996. https://doi.org/10.3390/ijms141122982
Chicago/Turabian StyleWu, Zhiming, Jiaowen Cheng, Cheng Qin, Zhiqun Hu, Caixia Yin, and Kailin Hu. 2013. "Differential Proteomic Analysis of Anthers between Cytoplasmic Male Sterile and Maintainer Lines in Capsicum annuum L." International Journal of Molecular Sciences 14, no. 11: 22982-22996. https://doi.org/10.3390/ijms141122982