Effect of Opuntia humifusa Supplementation and Acute Exercise on Insulin Sensitivity and Associations with PPAR-γ and PGC-1α Protein Expression in Skeletal Muscle of Rats

Abstract
:1. Introduction
2. Results
2.1. Body Weight, Food Efficiency Ratio (FER) and Fat Tissue Weight
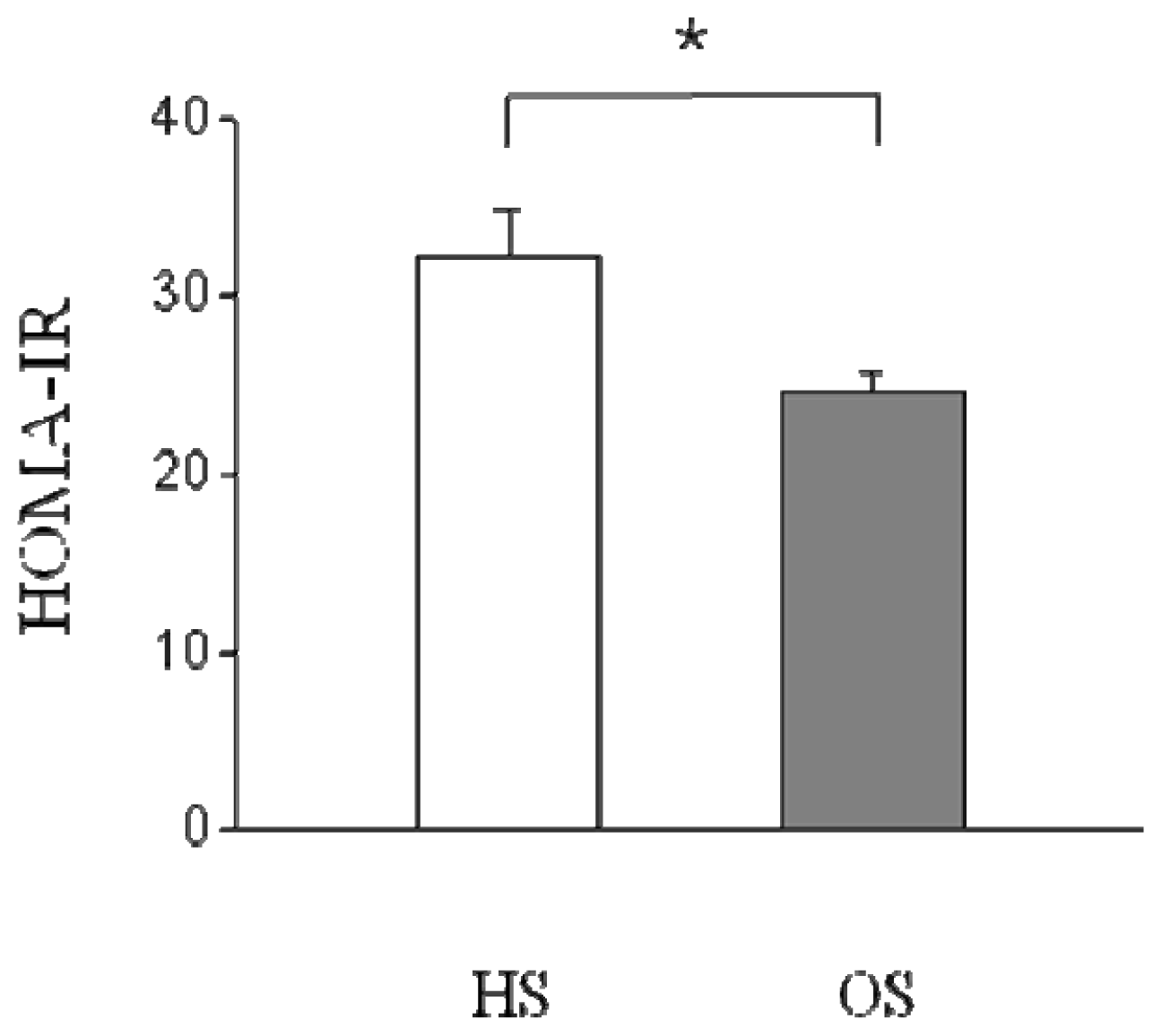
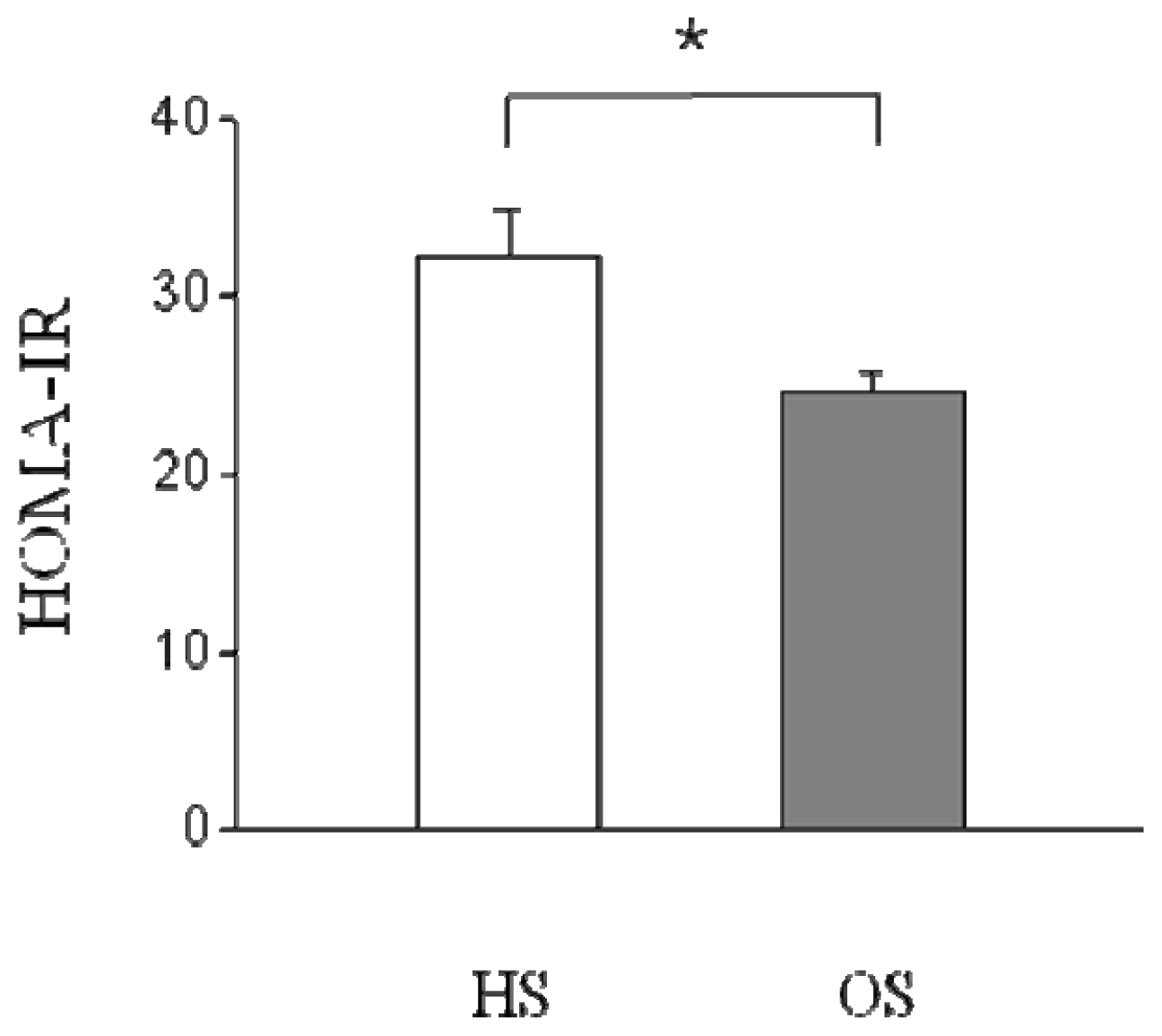
2.2. Changes in Serum Parameters and HOMA-IR
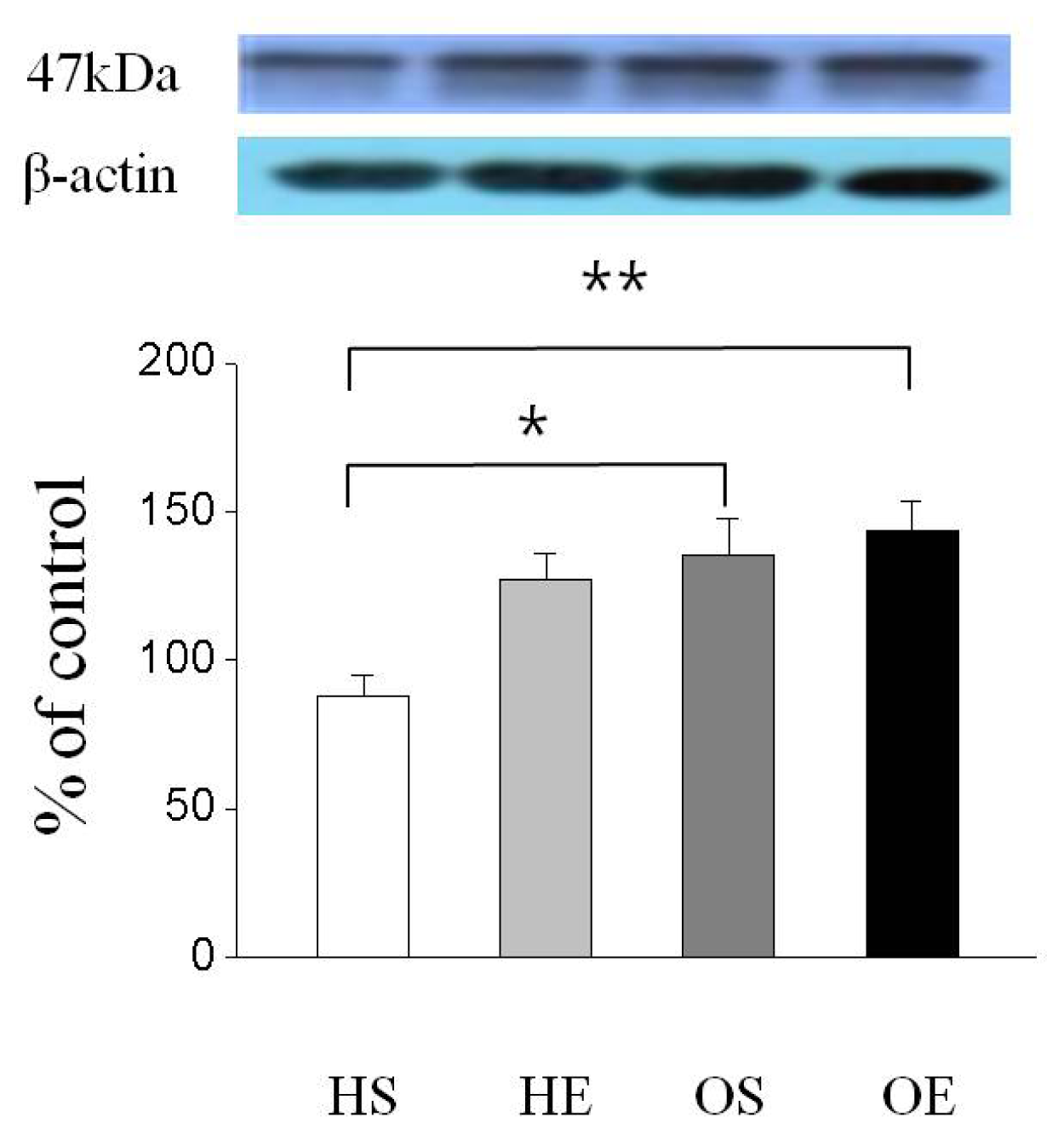
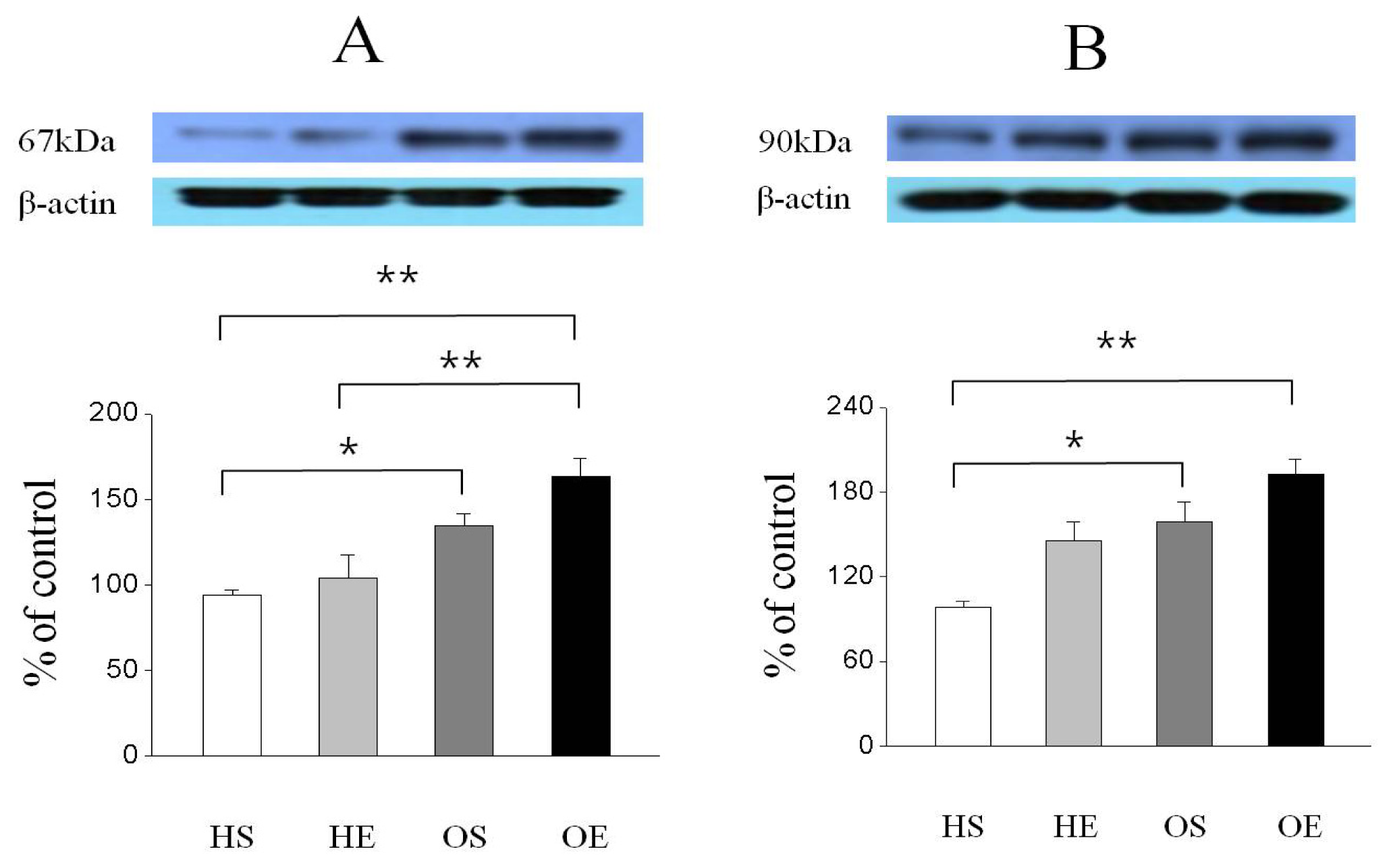
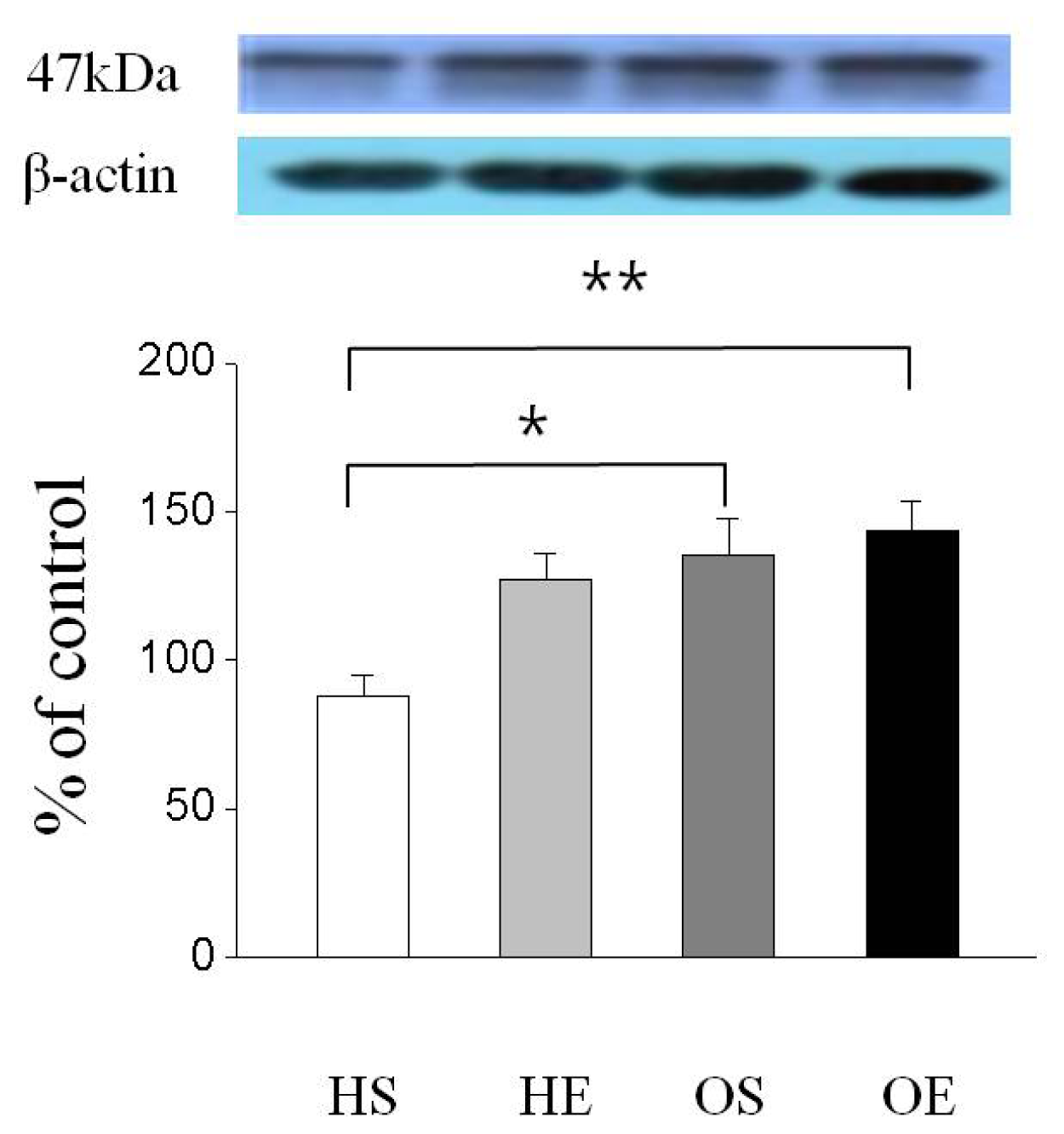
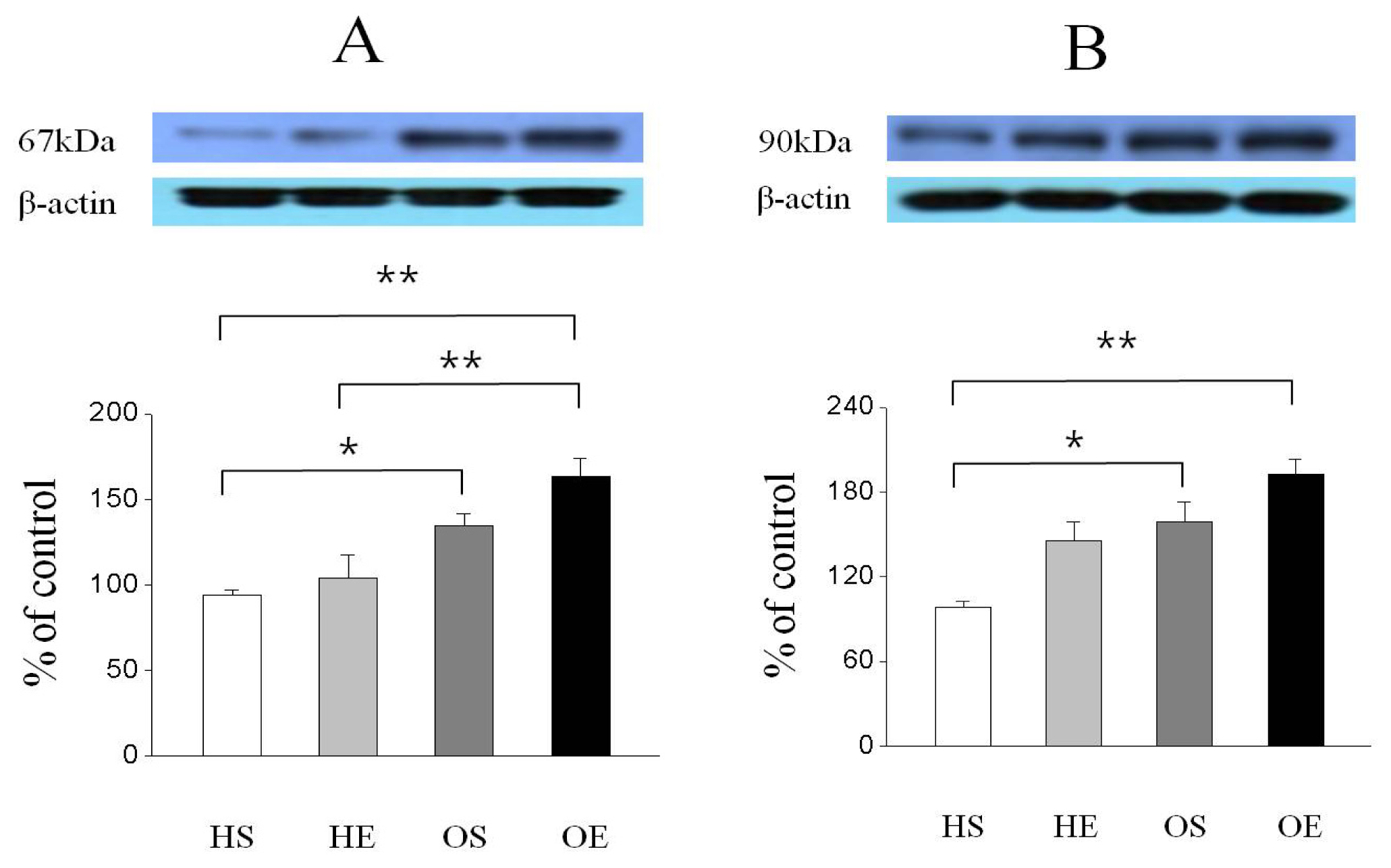
2.3. Skeletal Muscle PPAR-γ, PGC-1α, and GLUT-4 Protein Expressions
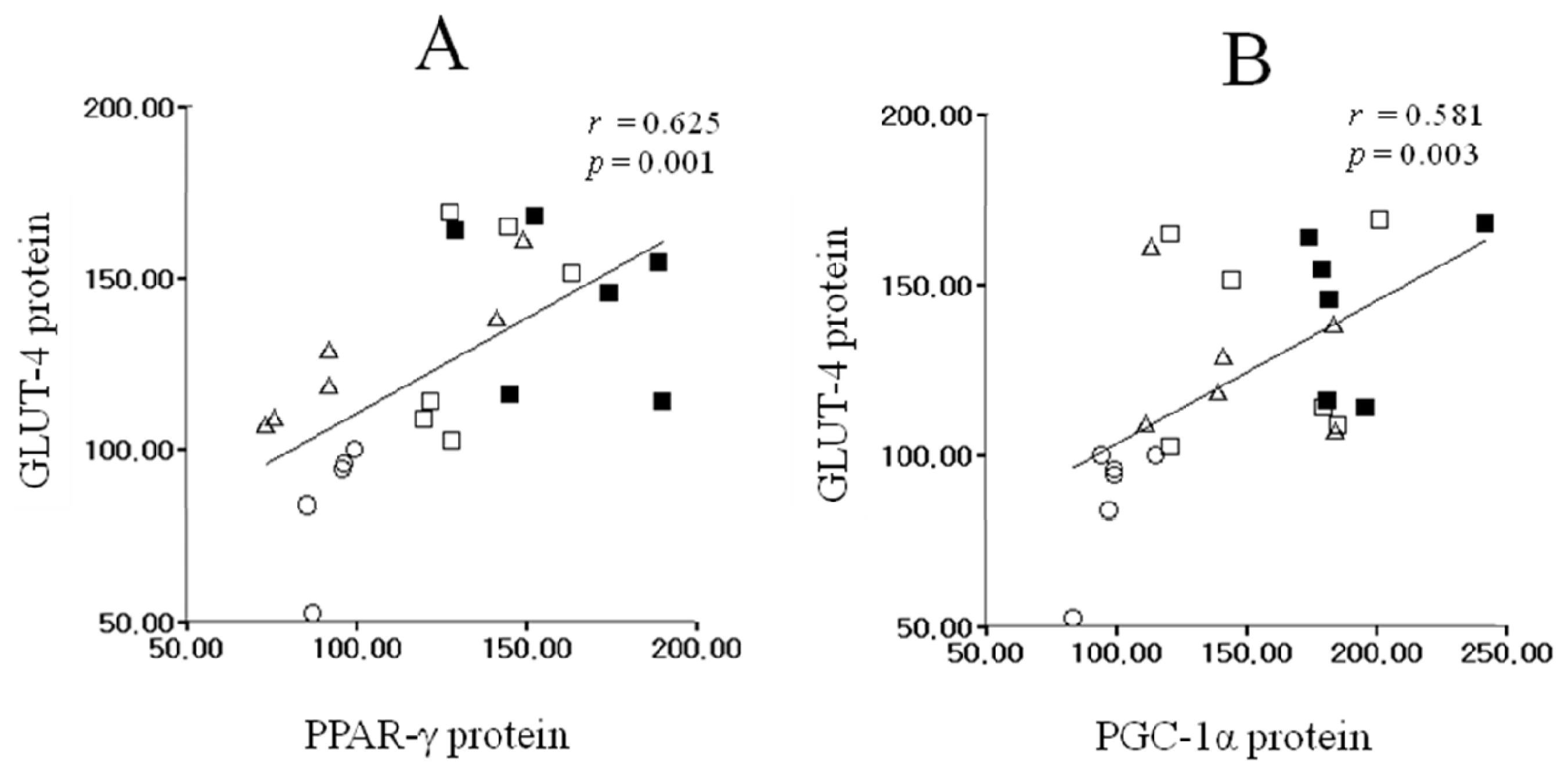
2.4. Correlation among Skeletal Muscle PPAR-γ, PGC-1α, and GLUT-4 Protein Expressions
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Exercise Protocol
4.2. Preparation of Experimental Diet
4.3. Serum Analysis
4.4. Western Blot Analysis
4.5. Statistical Analysis
5. Conclusions
Acknowledgements
References
- Haring, H.U.; Mehnert, H. Pathogenesis of Type 2 diabetes mellitus: Candidates for a signal transmitter defect causing insulin resistance of the skeletal muscle. Diabetologia 1993, 36, 176–182. [Google Scholar]
- DeFronzo, R.A.; Ferrannini, E. Insulin resistance: A multifaceted syndrome responsible for NIDDM, obesity, hypertension dyslipidemia and atherosclerotic cardiovascular disease. Diabetes Care 1991, 14, 173–194. [Google Scholar]
- DeFronzo, R.A.; Bonadonna, R.C.; Ferrannini, E. Pathogenesis of NIDDM. A balanced overview. Diabetes Care 1992, 15, 318–368. [Google Scholar]
- Ford, E.S.; Williamson, D.F.; Liu, S. Weight change and diabetes incidence: Findings from a national cohort of US adults. Am. J. Epidemiol 1997, 146, 214–222. [Google Scholar]
- Resnick, H.E.; Valsania, P.; Halter, J.B.; Lin, X. Relation of weight gain and weight loss on subsequent diabetes risk in overweight adults. J. Epidemiol. Commun. Health 2000, 54, 596–602. [Google Scholar]
- Picard, F.; Auwerx, J. PPAR (gamma) and glucose homeostasis. Annu. Rev. Nutr 2002, 22, 167–197. [Google Scholar]
- Wilson, T.M.; Brown, P.J.; Sternbach, D.D.; Henke, B.R. The PPARs: From orphan receptors to drug discovery. J. Med. Chem 2000, 43, 527–550. [Google Scholar]
- Miyazaki, Y.; He, H.; Mandarino, L.J.; de Fronzo, R.A. Resiglitazone improves downstream insulin receptor signaling in type 2 diabetic patients. Diabetes 2003, 52, 1943–1950. [Google Scholar]
- Kawaguchi, H.; Akune, T.; Yamaguchi, M.; Ohba, S.; Ogata, N.; Chung, U.I.; Kubota, N.; Terauchi, Y.; Kadowaki, T.; Nakamura, K. Distinct effects of PPAR gamma insufficiency on bone marrow cells, osteoblast, and osteoclastic cells. J. Bone Miner. Metab 2005, 23, 275–279. [Google Scholar]
- Lefebvre, A.M.; Chen, I.; Desteumaux, P.; Najib, J.; Fruchart, J.C.; Geboes, K.; Btiggs, M.; Heyman, R.; Auwerx, J. Activation of the peroxisome proliferator-activated receptor gamma promotes the development of colon tumors in C57BL/6JAPCMin/+mice. Nat. Med 1998, 4, 1053–1057. [Google Scholar]
- Song, Y.J.; Sawamura, M.; Ikeda, K.; Igawa, S.; Yamori, Y. Soluble dietary fibre improves insulin sensitivity by increasing muscle glut-4 content in stroke-prone spontaneously hypertensive rats. Clin. Exp. Pharmacol. Physiol 2000, 27, 41–45. [Google Scholar]
- Lee, S.H.; Lee, H.J.; Lee, Y.H.; Lee, B.W.; Cha, B.S.; Kang, E.S.; Ahn, C.W.; Park, J.S.; Kim, H.J.; Lee, E.Y.; et al. Korean red ginseng (Panax ginseng) improves insulin sensitivity in high fat fed Sprague-Dawley rats. Phytother. Res 2012, 26, 142–147. [Google Scholar]
- Goldstein, G.; Nobel, P.S. Water relations and low temperature acclimation for cactus species varying in freezing tolerance. Plant Physiol 1994, 104, 675–681. [Google Scholar]
- Kang, J.; Park, J.; Choi, S.H.; Igawa, S.; Song, Y. Opuntia humifusa supplementation increased bone density by regulating parathyroid hormone and osteocalcin in male growing rats. Int. J. Mol. Sci 2012, 13, 6747–6756. [Google Scholar]
- Cho, J.Y.; Park, S.C.; Kim, T.W.; Kim, K.S.; Song, J.C.; Kim, S.K.; Lee, H.M.; Sung, H.J.; Park, H.J.; Song, Y.B.; et al. Radical scavenging and anti-inflammatory activity of extracts from Opuntia humifusa Raf. J. Pharm. Pharmacol 2006, 58, 113–119. [Google Scholar]
- Park, E.H.; Kahng, J.H.; Lee, S.H.; Shin, K.H. An anti-inflammatory principles from cactus. Fitoterapia 2001, 72, 288–290. [Google Scholar]
- Chavez-Santoscoy, R.A.; Gutierrez-Uribe, J.A.; Serna-Saldivar, S.O. Phenolic composition, antioxidant capacity and in vitro cancer cell cytotoxicity of nine prickly pear (Opuntia spp.) juices. Plant Foods Hum. Nutr 2009, 64, 146–152. [Google Scholar]
- Butera, D.; Tesoriere, L.; Di Gaudio, F.; Bongiorno, A.; Allegra, M.; Pintaudi, A.M.; Kohen, R.; Livrea, M.A. Antioxidant activities of sicilian prickly pear (Opuntia ficus indica) fruit extracts and reducing properties of its betalains: Betanin and indicaxanthin. J. Agric. Food Chem 2002, 50, 6895–6901. [Google Scholar]
- Hahm, S.W.; Park, J.; Son, Y.S. Opuntia humifusa stems lower blood glucose and cholesterol levels in streptozotocin-induced diabetic rats. Nutr. Res 2011, 31, 479–487. [Google Scholar]
- Hamada, T.; Arias, E.B.; Cartee, G.D. Increased submaximal insulin-stimulated glucose uptake in mouse skeletal muscle after treadmill exercise. J. Appl. Physiol 2006, 101, 1368–1376. [Google Scholar]
- Ruschke, K.; Fishbein, L.; Dietrich, A.; Klöting, N.; Tönjes, A.; Oberbach, A.; Fasshauer, M.; Jenkner, J.; Schön, M.R.; Stumvoll, M.; et al. Gene expression of PPARgamma and PGC-1alpha in human omental and subcutaneous adipose tissues is related to insulin resistance markers and mediates beneficial effects of physical training. Eur. J. Endocrinol 2010, 162, 515–523. [Google Scholar]
- Hevener, A.L.; He, W.; Barak, Y.; Le, J.; Bandyopadhyay, G.; Olson, P.; Wilkes, J.; Evans, R.M.; Olefsky, J. Muscle-specific Pparg deletion causes insulin resistance. Nat. Med 2003, 9, 1491–1497. [Google Scholar]
- Shih, C.C.; Lin, C.H.; Lin, W.L.; Wu, J.B. Momordica charantia extract on insulin resistance and the skeletal muscle GLUT4 protein in fructose-fed rats. J. Ethnopharmacol 2009, 123, 82–90. [Google Scholar]
- Patti, M.E.; Butte, A.J.; Crunkhorn, S.; Cusi, K.; Berria, R.; Kashyap, S.; Miyazaki, Y.; Kohane, I.; Costello, M.; Saccone, R.; et al. Coordinated reduction of genes of oxidative metabolism in humans with insulin resistance and diabetes: Potential role of PGC1 and NRF1. Proc. Natl. Acad. Sci. USA 2003, 100, 8466–8471. [Google Scholar]
- Michael, L.F.; Wu, Z.; Cheatham, R.B.; Puigserver, P.; Adelmant, G.; Lehman, J.J.; Kelly, D.P.; Spiegelman, B.M. Restoration of insulin-sensitive glucose transporter (GLUT4) gene expression in muscle cells by the transcriptional coactivator PGC-1. Proc. Natl. Acad. Sci. USA 2001, 98, 3820–3825. [Google Scholar]
- De Munter, J.S.; Hu, F.B.; Spiegelman, D.; Franz, M.; van Dam, R.M. Whole grain, bran, and germ intake and risk of type 2 diabetes: A prospective cohort study and systematic review. PLoS Med 2007, 4, 1385–1395. [Google Scholar]
- Pittas, A.G.; Lau, J.; Hu, F.B.; Dawson-Hughes, B. The role of vitamin D and calcium in type 2 diabetes: A systemic review and meta-analysis. J. Clin. Endocrinol. Metab 2007, 92, 2017–2029. [Google Scholar]
- Takaya, J.; Higashino, H.; Kobayashi, Y. Intracellular magnesium and insulin resistance. Magnes. Res 2004, 17, 126–136. [Google Scholar]
- Lopez-Ridaura, R.; Willett, W.C.; Rimm, E.B.; Liu, S.; Stampfer, M.J.; Manson, J.E.; Hu, F.B. Magnesium intake and risk of type 2 diabetes in men and women. Diabetes Care 2004, 27, 134–140. [Google Scholar]
- Suarez, A.; Pulido, N.; Casla, A.; Casanova, B.; Arrieta, F.J.; Rovira, A. Impaired tyrosin-kinase activity of muscle insulin receptors from hypomagnesaemic rats. Diabetologia 1995, 38, 1262–1270. [Google Scholar]
- Berger, J.; Bailey, P.; Biswas, C.; Cullinan, C.A.; Doebber, T.W.; Hayes, N.S.; Saperstein, R.; Smith, R.G.; Leibowitz, M.D. Thiazolidinediones produce a conformational change in peroxisomal proliferator-activated receptor-gamma: Binding and activation correlate with antidiabetic actions in db/db mice. Endocrinology 1996, 137, 4189–4195. [Google Scholar]
- Guerrero-Romero, F.; Rodríguez-Morán, M. Pioglitazone increases serum magnesium levels in glucose-intolerant subjects. A randomized, controlled trial. Exp. Clin. Endocrinol. Diabetes 2003, 111, 91–96. [Google Scholar]
- Emoto, M.; Nishizawa, Y. Diabetes Mellitus and Magnesium. In New Perspectives in Magnesium Research; Nishizawa, Y., Morii, H., Durlach, J., Eds.; Springer-Verlag: London, UK, 2007; pp. 197–212. [Google Scholar]
- Garvey, W.T.; Maianu, L.; Hancock, J.A.; Golichowski, A.M.; Baron, A. Gene expression of GLUT4 in skeletal muscle from insulin-resistant patients with obesity, IGT, GDM, and NIDDM. Diabetes 1992, 41, 465–475. [Google Scholar]
- Begum, N.; Leitner, W.; Reusch, J.E.; Sussman, K.E.; Draznin, B. GLUT-4 phosphorylation and its intrinsic activity. Mechanism of Ca2+ induced inhibition of insulin-stimulated glucose transport. J. Biol. Chem 1993, 268, 3352–3356. [Google Scholar]
- Hellman, B.; Berne, C.; Grapengiesser, E.; Grill, V.; Gylfe, E.; Lund, P.E. The cytoplasmic Ca2+ response to glucose as an indicator of impairment of the pancreatic beta-cell function. Eur. J. Clin. Invest 1990, 20, S10–S17. [Google Scholar]
- Pershadsingh, H.A.; Shade, D.L.; Delfert, D.M.; McDonald, J.M. Chelation of intracellular calcium blocks insulin action in the adipocyte. Proc. Natl. Acad. Sci. USA 1987, 84, 1025–1029. [Google Scholar]
- Pikilidou, M.I.; Lasaridis, A.N.; Sarafidism, P.A.; Befani, C.D.; Koliakos, G.G.; Tziolas, I.M.; Kazakos, K.A.; Yovos, J.G.; Nilsson, P.M. Insulin sensitivity increase after calcium supplementation and change in intraplatelet calcium and sodium-hydrogen exchange in hypertensive patients with Type 2 diabetes. Diabet. Med 2009, 26, 211–219. [Google Scholar]
- Sánchez, M.; de la Sierra, A.; Coca, A.; Poch, E.; Giner, V.; Urbano-Márquez, A. Oral calcium supplementation reduces intraplatelet free calcium concentration and insulin resistance in essential hypertensive patients. Hypertension 1997, 29, 531–536. [Google Scholar]
- Dudley, G.A.; Terjung, R.L. Influence of acidosis on AMP deaminase activity in contracting fast-twitch muscle. Am. J. Physiol 1985, 248, C43–C50. [Google Scholar]
- Hutber, C.A.; Hardie, D.G.; Winder, W.W. Electrical stimulation inactivates muscle acetyl-CoA carboxylase and increases AMP-activated protein kinase. Am. J. Physiol 1997, 272, E262–E266. [Google Scholar]
- Tarada, S.; Kawanaka, K.; Goto, M.; Shimokawa, T.; Tabata, I. Effect of high-intensity intermittent swimming on PGC-1α protein expression in rat skeletal muscle. Acta Physiol. Scand 2005, 184, 59–65. [Google Scholar]
- Calvo, J.A.; Daniels, T.G.; Wang, X.; Paul, A.; Lin, J.; Spiegelman, B.M.; Stevenson, S.C.; Rangwala, S.M. Muscle specific expression of PPARγ coactivator-1α improves exercise performance and increases peak oxygen uptake. J. Appl. Physiol 2008, 104, 1304–1324. [Google Scholar]
- Kawamura, T.; Yoshida, K.; Sugawara, A.; Nagasaka, M.; Mori, N.; Takeuchi, K.; Kohzuki, M. Regulation of skeletal muscle peroxisome proliferator-activated receptor gamma expression by exercise and angiotensin-converting enzyme inhibition in fructose-fed hypertensive rats. Hypertens. Res 2004, 27, 61–70. [Google Scholar]
- Russell, A.P.; Feilchenfeldt, J.; Schreiber, S.; Praz, M.; Crettenand, A.; Gobelet, C.; Meier, C.A.; Bell, D.R.; Kralli, A.; Giacobino, J.P.; et al. Endurance training in humans leads to fiber type specific increases in levels of peroxisome proliferator activated receptor gamma coactivator 1 and peroxisome proliferator activated receptor alpha in skeletal muscle. Diabetes 2003, 52, 2874–2881. [Google Scholar]
- De Filippis, E.; Alvarez, G.; Berria, R.; Cusi, K.; Everman, S.; Meyer, C.; Mandarino, L.J. Insulin-resistant muscle is exercise resistant: Evidence for reduced response of nuclear-encoded mitochondrial genes to exercise. Am. J. Physiol. Endocrinol. Metab 2008, 294, 607–614. [Google Scholar]
- Baar, K.; Wende, A.R.; Jones, T.E.; Marison, M.; Nolte, L.A.; Chen, M.; Kelly, D.P.; Holloszy, J.O. Adaptations of skeletal muscle to exercise: Rapid increase in the transcriptional coactivator PGC-1. FASEB J 2002, 16, 1879–1886. [Google Scholar]
- Russell, A.P.; Hessenlink, M.K.; Lo, S.K.; Schrauwen, P. Regulation of metabolic transcriptional co-activators and transcription factors with acute exercise. FASEB J 2005, 19, 986–988. [Google Scholar]
- Tunstall, R.J.; Mehan, K.A.; Wadley, G.D.; Collier, G.R.; Bonen, A.; Hargreaves, M.; Cameron-Smith, D. Exercise training increases lipid metabolism gene expression in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab 2002, 283, E66–E72. [Google Scholar]
- Loviscach, M.; Rehman, N.; Carter, L.; Mudaliar, S.; Mohadeen, P.; Ciaraldi, T.P.; Veerkamp, J.H.; Henry, R.R. Distribution of peroxisome proliferator-activated receptors (PPARs) in human skeletal muscle and adipose tissue: Relation to insulin action. Diabetologia 2000, 43, 304–311. [Google Scholar]
- Baron, A.D.; Brechtel, G. Insulin differentially regulates systemic and skeletal muscle vascular resistance. Am. J. Physiol 1993, 265, E61–E67. [Google Scholar]
- Puigserver, P.; Spiegelman, B.M. Peroxisome proliferator-activated receptor-gamma coactivator 1alpha (PGC-1 alpha): Transcriptional coactivator and metabolic regulator. Endocr. Rev 2003, 24, 78–90. [Google Scholar]
- Ling, C.; Poulsen, P.; Carlsson, E.; Ridderstråle, M.; Almgren, P.; Wojtaszewski, J.; Beck-Nielsen, H.; Groop, L.; Vaag, A. Multiple environmental and genetic factors influence skeletal muscle PGC-1α and PGC-1β gene expression in twins. J. Clin. Invest 2004, 114, 1518–1526. [Google Scholar]
- Crunkhorn, S.; Dearie, F.; Mantzoros, C.; Gami, H.; da Silva, W.S.; Espinoza, D.; Faucette, R.; Barry, K.; Bianco, A.C.; Patti, M.E. Peroxisome proliferator activator receptor gamma coactivator-1 expression is reduced in obesity: Potential pathogenic role of saturated fatty acids and p38 mitogen-activated protein kinase activation. J. Biol. Chem 2007, 282, 15439–15450. [Google Scholar]
- Association of Official Analytical Chemists (AOAC), Official Methods of Analysis of the Association of Analytical Chemists, 16th ed; AOAC: Washington DC, USA, 1998.
- Kim, Y.J.; Park, T.S. Genes are differentially expressed in the epididymal fat of rats rendered obese by a high-fat diet. Nutr. Res 2008, 28, 414–422. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HS | HE | OS | OE | |
|---|---|---|---|---|
| Initial body weight (g) | 254.8 ± 3.66 | 255.4 ± 5.52 | 255.3 ± 2.46 | 254.7 ± 5.45 |
| Final body weight (g) | 445.9 ± 13.21 | 454.7 ± 12.33 | 448.3 ± 9.70 | 444.9 ± 6.99 |
| Food efficiency ratio | 0.23 ± 0.010 | 0.24 ± 0.010 | 0.23 ± 0.009 | 0.23 ± 0.011 |
| Abdominal fat tissue (g) | 9.1 ± 0.80 | 8.2 ± 0.62 | 9.4 ± 0.66 | 8.7 ± 0.66 |
| Epididymal fat tissue (g) | 8.5 ± 0.75 | 8.0 ± 0.83 | 8.6 ± 0.61 | 8.4 ± 0.43 |
| HS | HE | OS | OE | |
|---|---|---|---|---|
| Glucose (mg/dL) | 157.8 ± 5.05 | 122.1 ± 2.63 # | 119.1 ± 2.16 ** | 107.2 ± 5.99 §§ |
| Insulin (ng/mL) | 3.7 ± 0.26 | 0.8 ± 0.12 ## | 2.5 ± 0.37 *,§§ | 0.4 ± 0.05 ##,§§ |
| TG (mg/dL) | 52.4 ± 4.66 | 46.2 ± 3.42 | 30.9 ± 2.66 * | 77.0 ± 5.88 **,##,§§ |
| FFA (mEq/L) | 319.4 ± 8.55 | 438.1 ± 13.81 # | 328.6 ± 21.80 §§ | 560.8 ± 29.48 **,##,§§ |
| TC (mg/dL) | 77.1 ± 2.06 | 82.1 ± 5.80 | 68.5 ± 3.12 | 87.9 ± 4.63 # |
| HDLC (mg/dL) | 23.4 ± 0.39 | 23.9 ± 0.70 | 23.9 ± 1.29 | 22.8 ± 0.94 |
| Ingredients | Contents |
|---|---|
| moisture (% w/w) | 2.9 |
| ash (% w/w) | 13.8 |
| carbohydrate (g/100 g) | 46.6 |
| crude protein (g/100 g) | 4.9 |
| crude fat (g/100 g) | 3.1 |
| fiber (g/100 g) | 28.9 |
| Fe2+ (mg/g) | 5.8 |
| Ca2+ (mg/100 g) | 2931.3 |
| Mg2+ (mg/100 g) | 1227.9 |
| K+ (mg/100 g) | 2155.5 |
| Na+ (mg/100 g) | 30.9 |
| P2+ (mg/100 g) | 653.2 |
| Ingredients | High fat diet | 5% O. humifusa added diet |
|---|---|---|
| Casein | 200 | 197.6 |
| Starch | 111 | 87.8 |
| Sucrose | 370 | 370 |
| Lard | 170 | 170 |
| Corn oil | 30 | 28.4 |
| Cellulose | 50 | 35.6 |
| Vitamin mix. | 12 | 12 |
| Mineral mix. | 42 | 42 |
| Cholesterol | 10 | 10 |
| d,l-methionine | 3 | 3 |
| Choline barbiturate | 2 | 2 |
| Tert-butylhydroquinone | 0.04 | 0.04 |
| O. humifusa | · | 50 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kang, J.; Lee, J.; Kwon, D.; Song, Y. Effect of Opuntia humifusa Supplementation and Acute Exercise on Insulin Sensitivity and Associations with PPAR-γ and PGC-1α Protein Expression in Skeletal Muscle of Rats. Int. J. Mol. Sci. 2013, 14, 7140-7154. https://doi.org/10.3390/ijms14047140
Kang J, Lee J, Kwon D, Song Y. Effect of Opuntia humifusa Supplementation and Acute Exercise on Insulin Sensitivity and Associations with PPAR-γ and PGC-1α Protein Expression in Skeletal Muscle of Rats. International Journal of Molecular Sciences. 2013; 14(4):7140-7154. https://doi.org/10.3390/ijms14047140
Chicago/Turabian StyleKang, Junyong, Junghun Lee, Daekeun Kwon, and Youngju Song. 2013. "Effect of Opuntia humifusa Supplementation and Acute Exercise on Insulin Sensitivity and Associations with PPAR-γ and PGC-1α Protein Expression in Skeletal Muscle of Rats" International Journal of Molecular Sciences 14, no. 4: 7140-7154. https://doi.org/10.3390/ijms14047140