Isolation of Multipotent Nestin-Expressing Stem Cells Derived from the Epidermis of Elderly Humans and TAT-VHL Peptide-Mediated Neuronal Differentiation of These Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
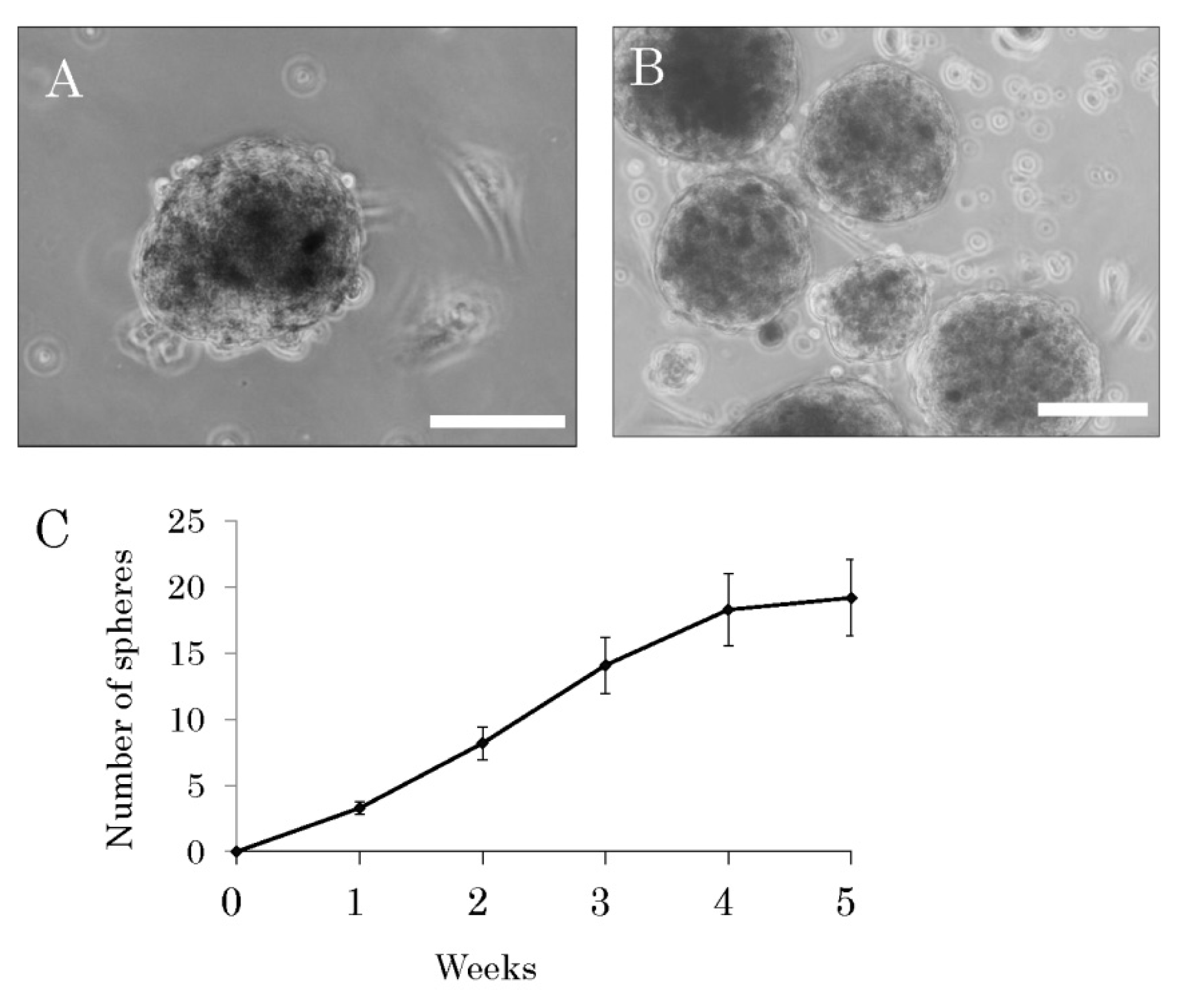
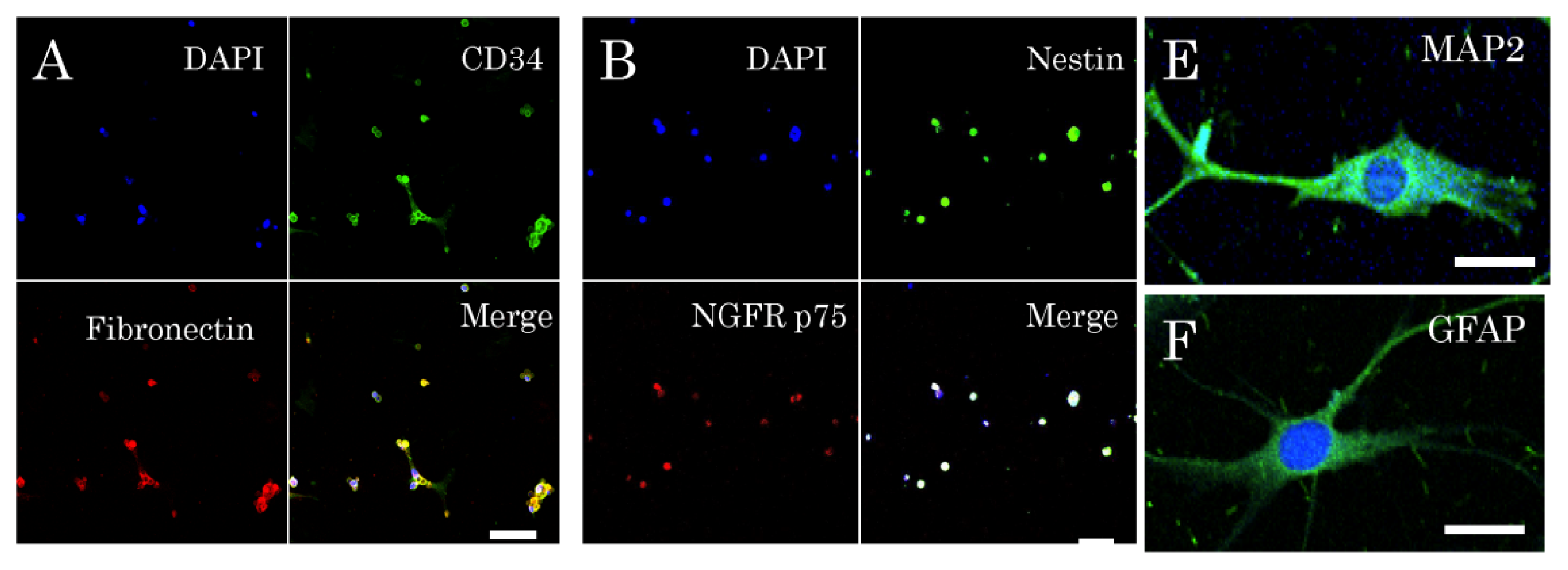
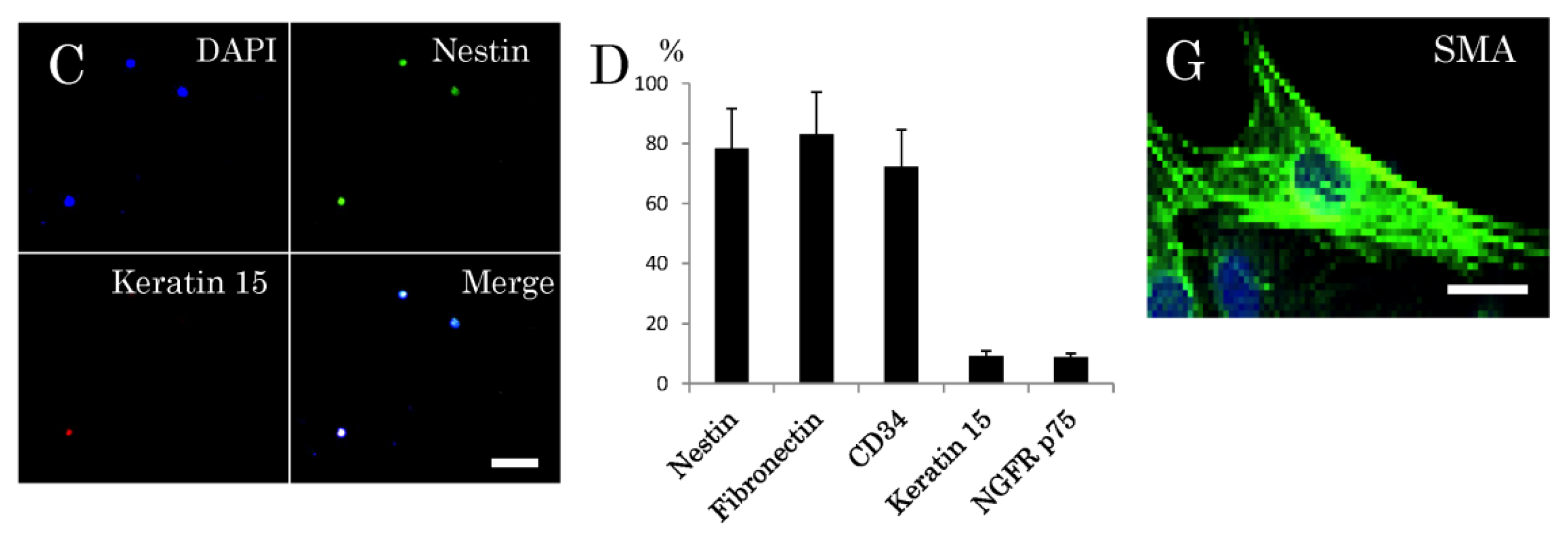
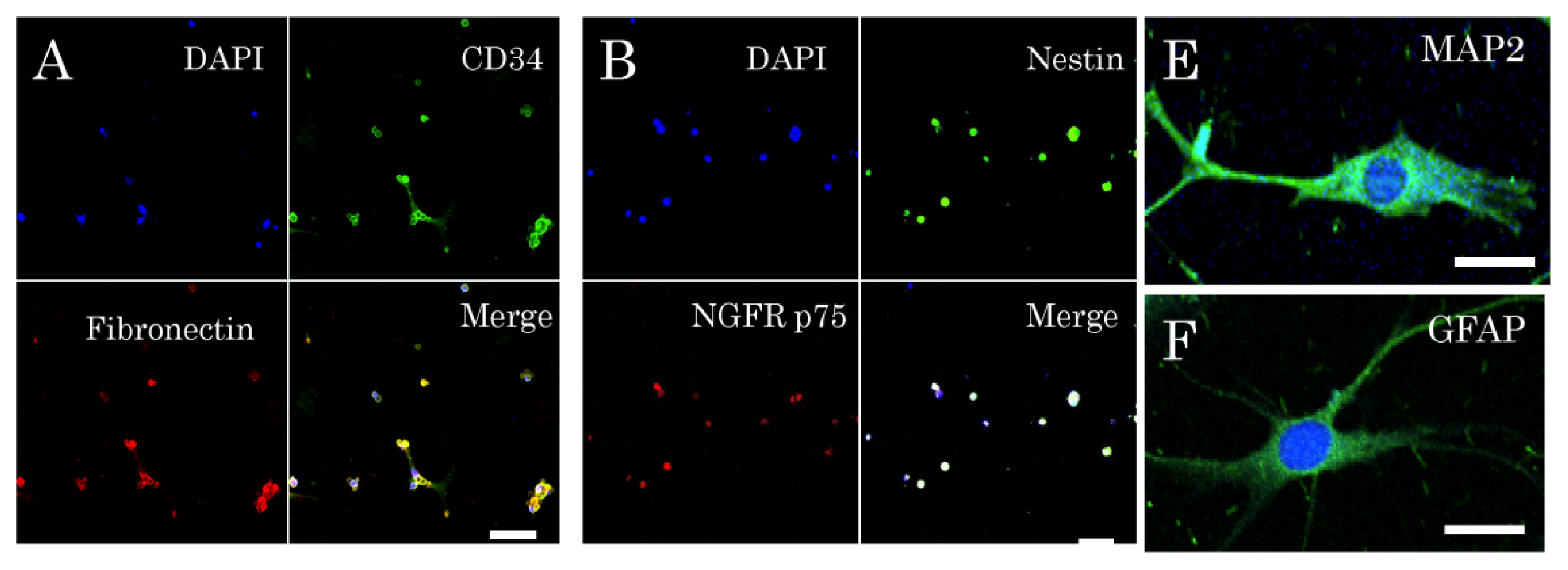
2.1. Isolation and Characterization of Multipotent Nestin-Expressing Stem Cells Derived from the Epidermis of Elderly Patients
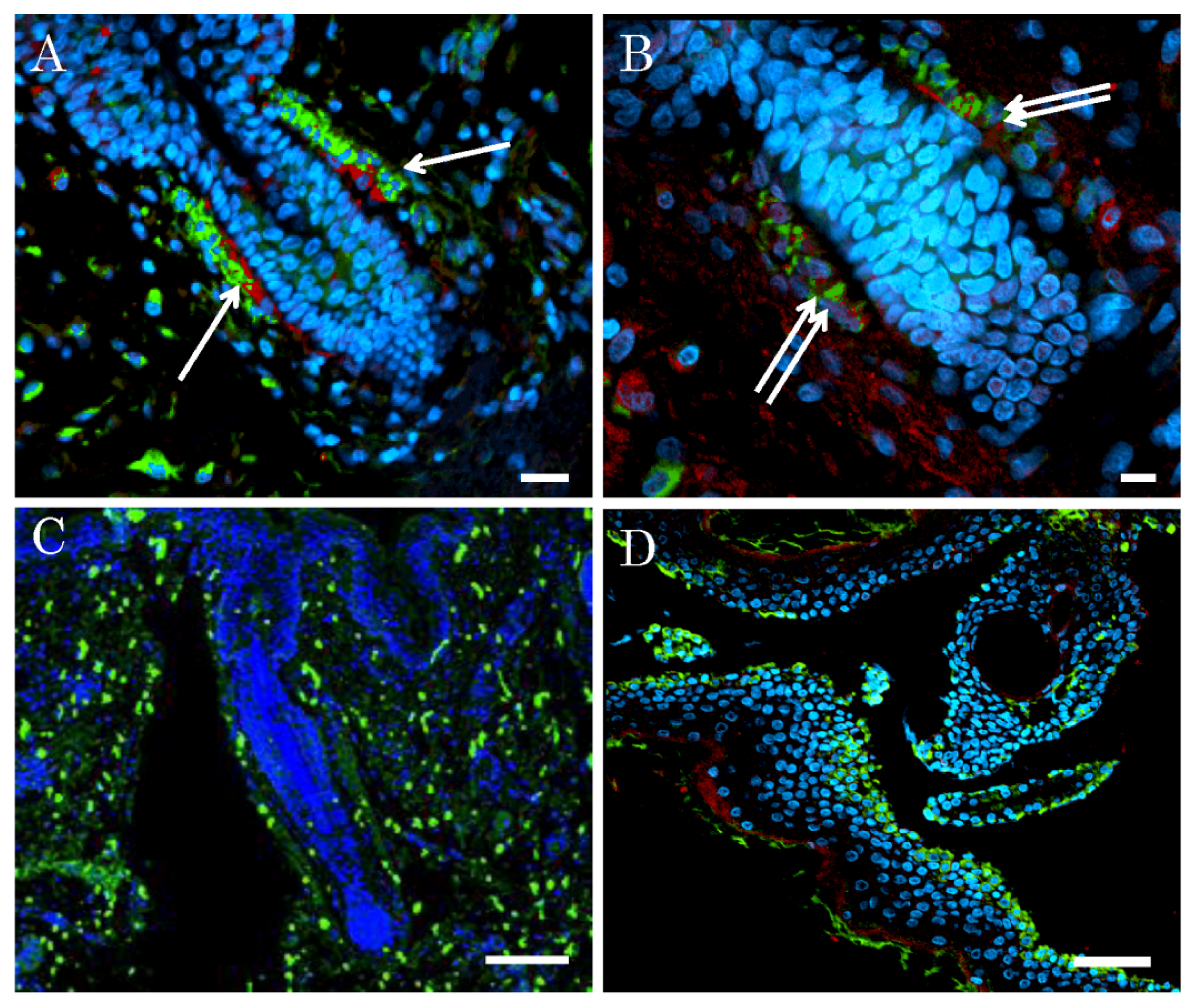
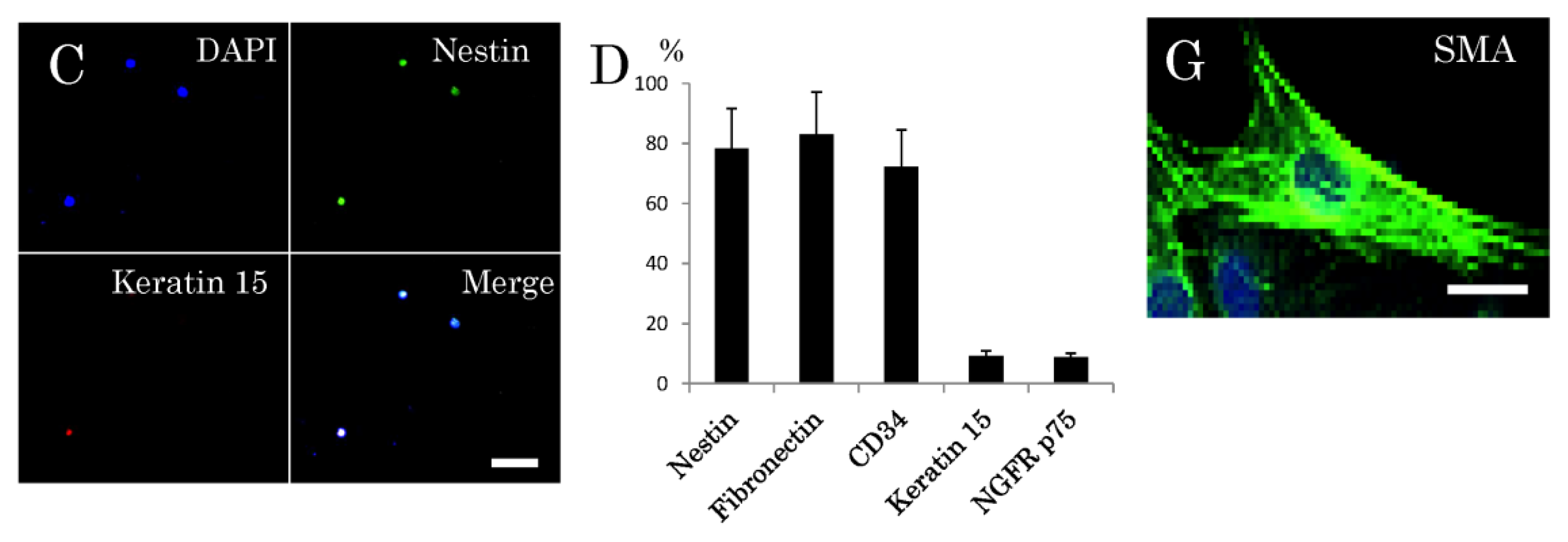
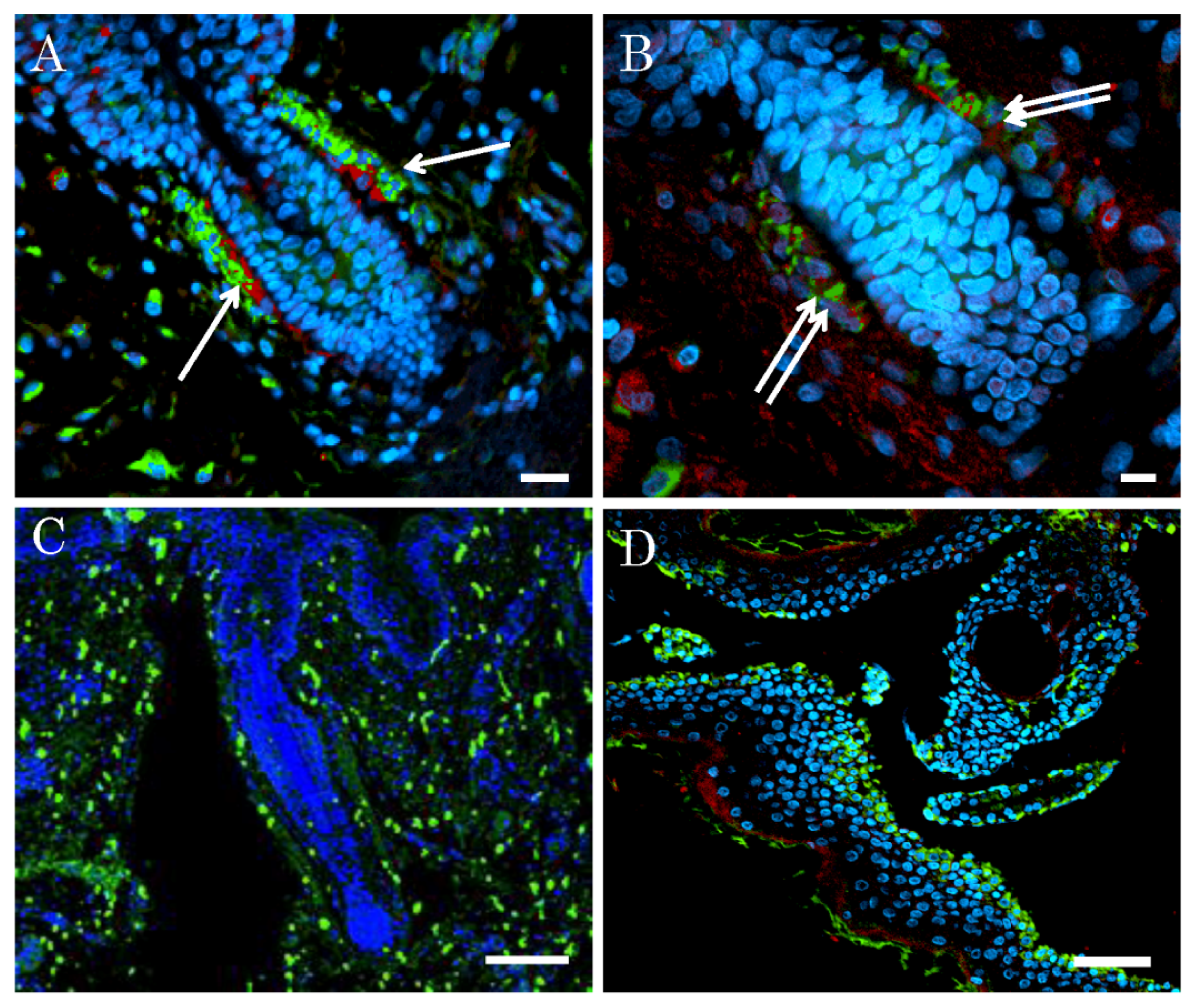
2.2. Niche of Multipotent Nestin-Expressing Stem Cells Derived from Epidermis of Elderly Humans
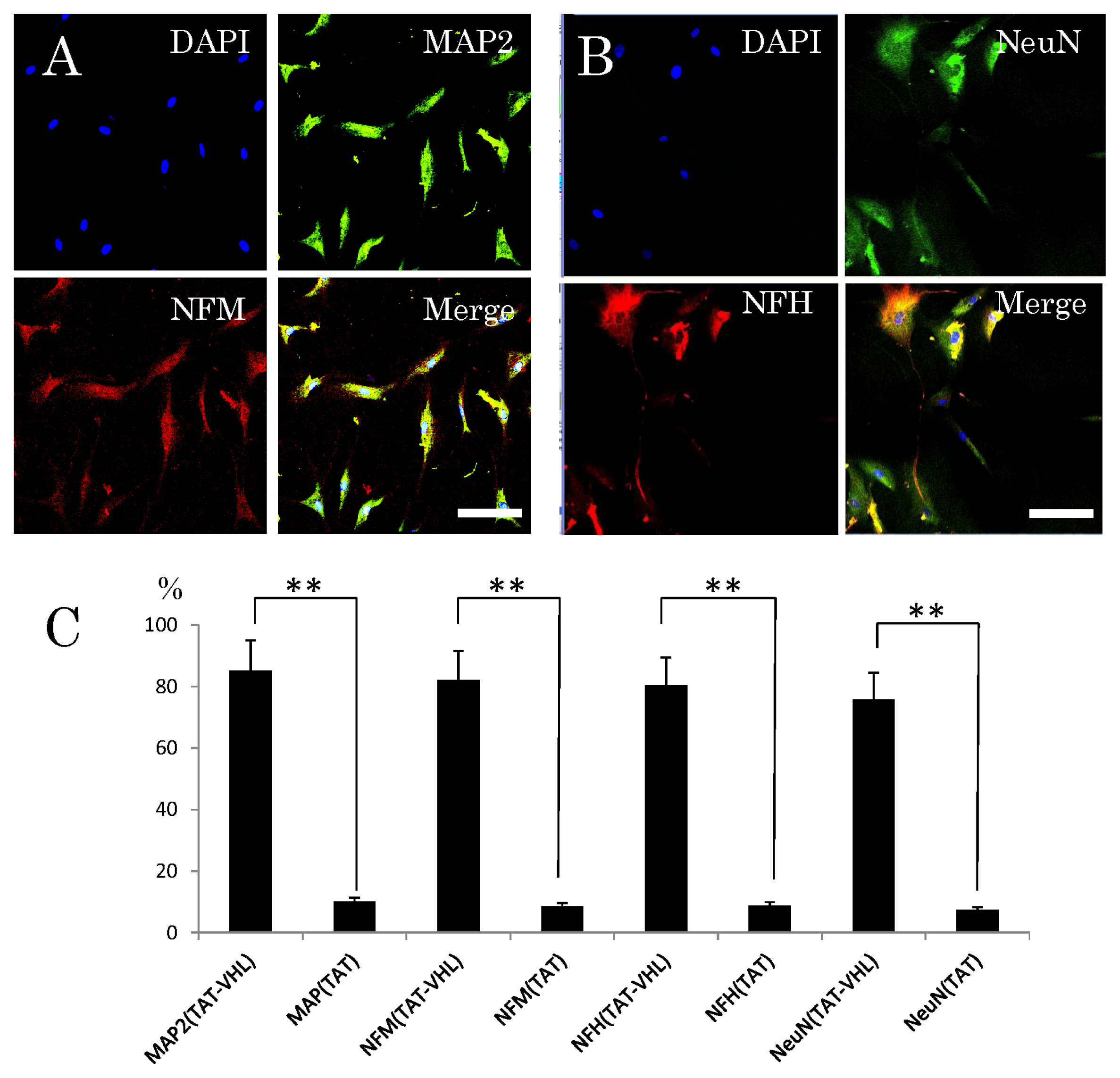
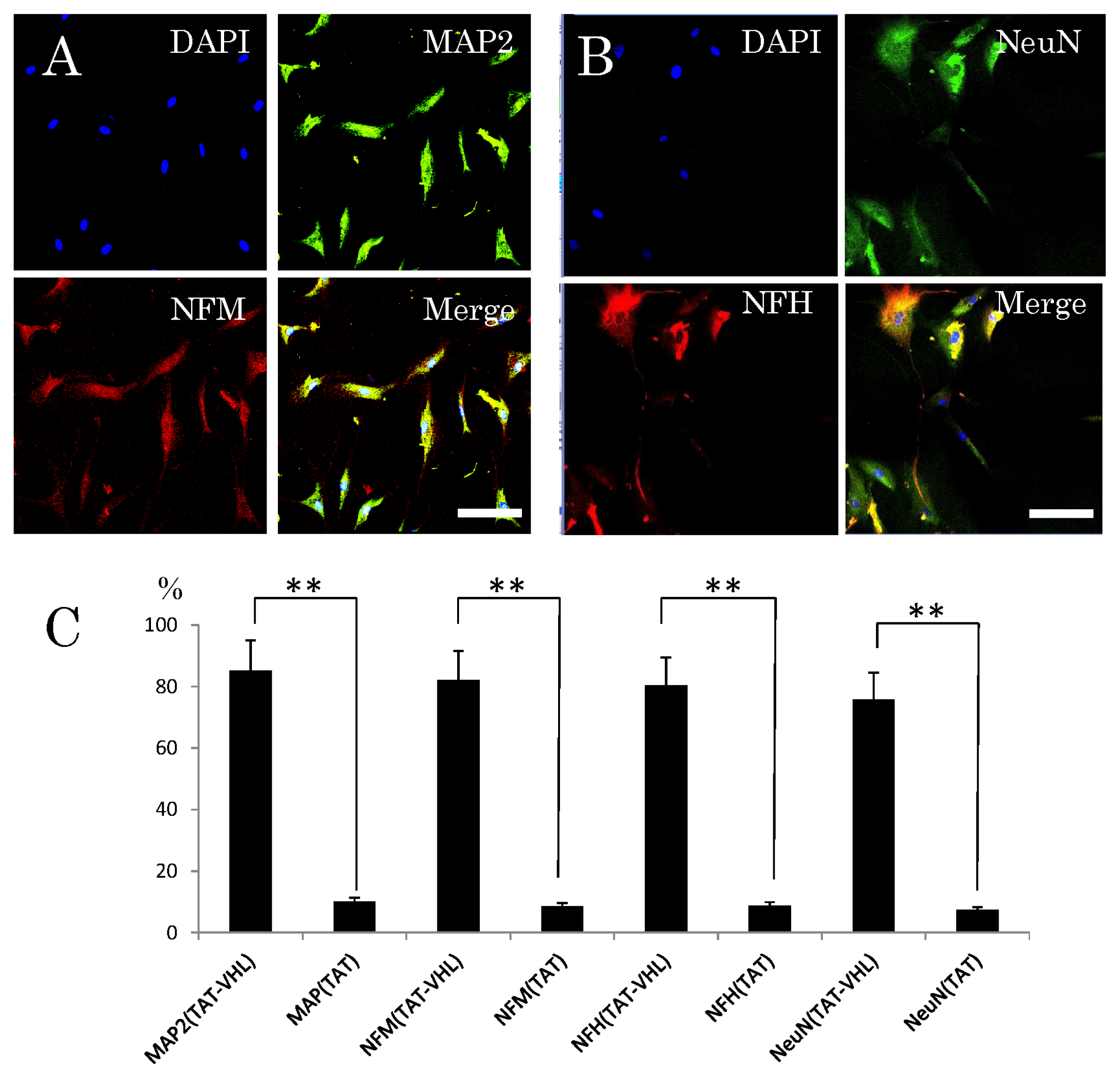
2.3. Neuronal Differentiation of Stem Cells after Intracellular Delivery of TAT-VHL Peptide
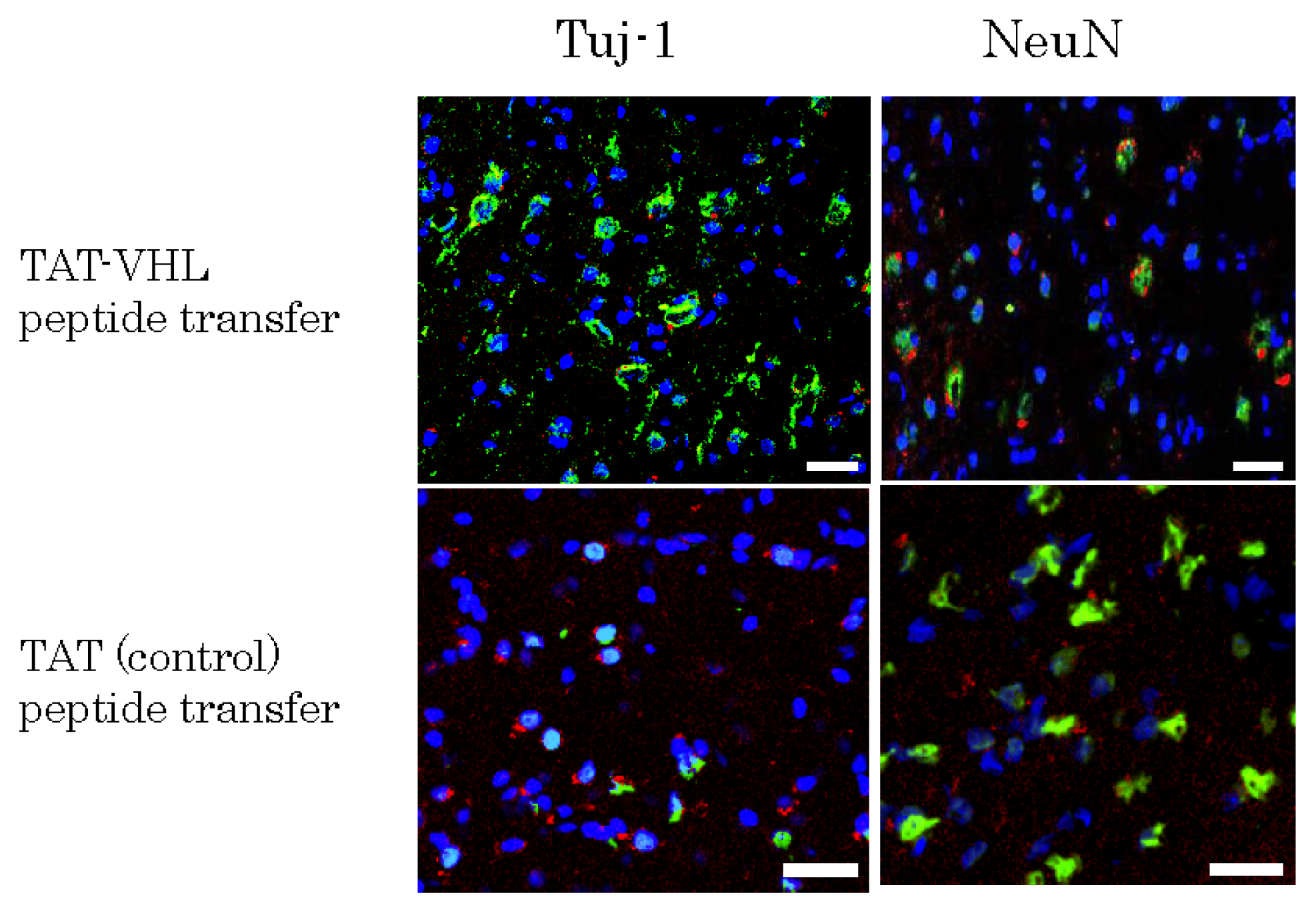
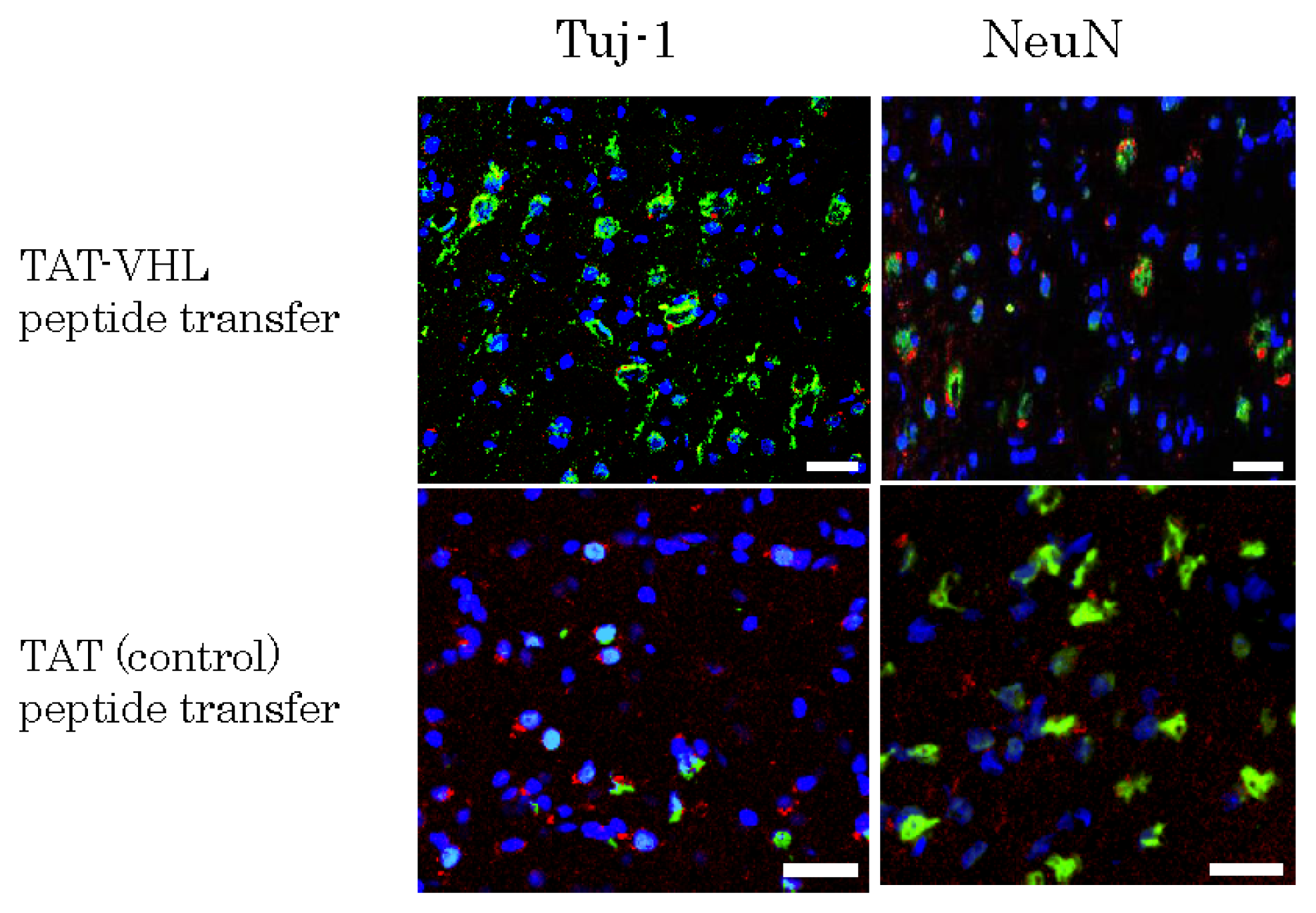
2.4. Implantation of Mutipotent Nestin-Expressing Stem Cells into the Rodent Brain
3. Discussion
4. Experimental Section
4.1. Cell Culture
4.2. Identification of Niche of Multipotent Nestin-Expressing Stem Cells Derived from Human Epidermis
4.3. Immunocytochemical Characterization of Multipotent Stem Cells
4.4. Sphere-Forming Assay
4.5. Induction of Neuronal Differentiation with TAT-VHL Peptide
4.6. Implantation of Multipotent Stem Cells Derived from Epidermis
4.7. Statistics
4.8. Ethical Approval
5. Conclusions
Acknowledgements
Conflict of Interest
References
- Abbas, O.; Mahalingam, M. Epidermal stem cells: Practical perspectives and potential uses. Br. J. Dermatol 2009, 161, 228–236. [Google Scholar]
- Yu, H.; Fang, D. Isolation of a novel population of multipotent adult stem cells from human follicles. Am. J. Pathol 2006, 168, 1879–1888. [Google Scholar]
- Amoh, Y.; Mii, S.; Aki, R.; Hamada, Y.; Kawahara, K.; Hoffman, R.M.; Katsuoka, K. Multipotent nestin-expressing stem cells capable of forming neurons are located in the upper, middle and lower part of the vibrissa hair follicle. Cell Cycle 2012, 11, 3513–3517. [Google Scholar]
- Amoh, Y.; Li, L.; Katsuoka, K.; Hoffman, R.M. Multipotent nestin-expressing hair follicle stem cells. J. Dermatol 2009, 36, 1–9. [Google Scholar]
- Sieber-Blum, M.; Grim, M.; Hu, Y.F.; Szeder, V. Pluripotent neural crest stem cells in the adult hair follicle. Dev. Dyn 2004, 231, 258–269. [Google Scholar]
- Hill, R.P.; Gledhill, K.; Gardner, A.; Higgins, C.A.; Crawford, H.; Lawrence, C.; Hutchison, C.J.; Owens, W.A.; Kara, B.; James, S.E.; et al. Generation and characterization of multipotent stem cells from established dermal cultures. PLoS One 2012, 7, e50742. [Google Scholar]
- Toma, J.G.; McKenzie, I.A.; Bagli, D.; Miller, F.D. Isolation and characterization of multipotent skin-derived precursors from human skin. Stem Cells 2005, 23, 727–737. [Google Scholar]
- Liu, F.; Uchugonova, A.; Kimura, H.; Zhang, C.; Zhao, M.; Zhang, L.; Koenig, K.; Duong, J.; Aki, R.; Saito, N.; et al. The bulge area is the major hair follicle source of nestin-expressing pluripotent stem cells which can repair the spinal cord compared to the dermal papilla. Cell Cycle 2011, 10, 830–839. [Google Scholar]
- Uchugonova, A.; Duong, J.; Zhang, N.; König, K.; Hoffman, R.M. The bulge area is the origin of nestin-expressing pluripotent stem cells of the hair follicle. J. Cell. Biochem 2011, 112, 2046–2050. [Google Scholar]
- Amoh, Y.; Li, L.; Yang, M.; Moossa, A.R.; Katsuoka, K.; Penman, S.; Hoffman, R.M. Nascent blood vessels in the skin arise from nestin-expressing hair-follicle cells. Proc. Natl. Acad. Sci. USA 2004, 101, 13291–13295. [Google Scholar]
- Uchugonova, A.; Hoffman, R.M.; Weinige, M.; Koenig, K. Watching stem cells in the skin of living mice noninvasively. Cell Cycle 2011, 10, 2017–2020. [Google Scholar]
- Duong, J.; Mii, S.; Uchugonova, A.; Liu, F.; Moossa, A.R.; Hoffman, R.M. Real-time confocal imaging of trafficking of nestin-expressing multipotent stem cells in mouse whiskers in long-term 3-D histoculture. In Vitro Cell. Dev. Biol. Anim 2012, 48, 301–305. [Google Scholar]
- Amoh, Y.; Kanoh, M.; Niiyama, S.; Kawahara, K.; Sato, Y.; Katsuoka, K.; Hoffman, R.M. Human and mouse hair follicles contain both multipotent and monopotent stem cells. Cell Cycle 2009, 8, 176–177. [Google Scholar]
- Brownell, I.; Guevara, E.; Bai, C.B.; Loomis, C.A.; Joyner, A.L. Nerve-derived sonic hedgehog defines a niche for hair follicle stem cells capable of becoming epidermal stem cells. Cell Stem Cell 2011, 8, 552–565. [Google Scholar]
- Biernaskie, J.; Paris, M.; Morozova, O.; Fagan, B.M.; Marra, M.; Pevny, L.; Miller, F.D. SKPs derive from hair follicle precursors and exhibit properties of adult dermal stem cells. Cell Stem Cell 2009, 5, 610–623. [Google Scholar]
- Amoh, Y.; Li, L.; Katsuoka, K.; Penman, S.P.; Hoffman, R.M. Multipotent nestin-positive, keratin-negative hair-follicle bulge stem cells can form neurons. Proc. Natl. Acad. Sci. USA 2005, 102, 5530–5534. [Google Scholar]
- Kloepper, J.E.; Tiede, S.; Brinckmann, J.; Reinhardt, D.P.; Meyer, W.; Faessler, R.; Paus, R. Immunophenotyping of the human bulge region: The quest to define useful in situ markers for human epithelial hair follicle stem cells and their niche. Exp. Dermatol 2008, 17, 592–609. [Google Scholar]
- Tiede, S.; Koop, N.; Kloepper, J.E.; Fässler, R.; Paus, R. Nonviral in situ green fluorescent protein labeling and culture of primary, adult human hair follicle epithelial progenitor cells. Stem Cells 2009, 27, 2793–2803. [Google Scholar]
- Garza, L.A.; Yang, C.C.; Zhao, T.; Blatt, H.B.; Lee, M.; He, H.; Stanton, D.C.; Carrasco, L.; Spiegel, J.H.; Tobias, J.W.; et al. Bald scalp in men with androgenetic alopecia retains hair follicle stem cells but lacks CD200-rich and CD34-positive hair follicle progenitor cells. J. Clin. Invest 2011, 121, 613–622. [Google Scholar]
- Amoh, Y.; Li, L.; Campillo, R.; Kawahara, K.; Katsuoka, K.; Penman, S.; Hoffman, R.M. Implanted hair follicle stem cells form Schwann cells that support repair of severed peripheral nerves. Proc. Natl. Acad. Sci. USA 2005, 102, 17734–17738. [Google Scholar]
- Amoh, Y.; Li, L.; Katsuoka, K.; Hoffman, R.M. Multipotent hair follicle stem cells promote repair of spinal cord injury and recovery of walking function. Cell Cycle 2008, 7, 1865–1869. [Google Scholar]
- Amoh, Y.; Kanoh, M.; Niiyama, S.; Hamada, Y.; Kawahara, K.; Sato, Y.; Hoffman, R.M.; Katsuoka, K. Human hair follicle pluripotent stem (hfPS) cells promote regeneration of peripheral-nerve injury: An advantageous alternative to ES and iPS cells. J. Cell. Biochem 2009, 107, 1016–1020. [Google Scholar]
- Kanno, H.; Saljooque, F.; Yamamoto, I.; Hattori, S.; Yao, M.; Shuin, T.; UH-S. Role of the von Hippel-Lindau tumor suppressor protein during neuronal differentiation. Cancer Res 2000, 60, 2820–2824. [Google Scholar]
- Kanno, H.; Nakano, S.; Kubo, A.; Mimura, T.; Tajima, N.; Sugimoto, N. Neuronal differentiation of neural progenitor cells by intracellular delivery of synthetic oligopeptide derived from von Hippel-Lindau protein. Protein Pept. Lett 2009, 16, 1291–1296. [Google Scholar]
- Kubo, A.; Yoshida, T.; Kobayashi, N.; Yokoyama, T.; Mimura, T.; Nishiguchi, T.; Higashida, T.; Yamamoto, I.; Kanno, H. Efficient generation of dopamine neuron-like cells from skin-derived precursors with a synthetic peptide derived from von Hippel-Lindau protein. Stem Cells Dev. 2009, 1–8, 1523–1532. [Google Scholar]
- Maeda, K.; Kanno, H.; Yamazaki, Y.; Kubo, A.; Sato, F.; Yamaguchi, Y.; Saito, T. Transplantation of von Hippel-Lindau peptide delivered neural stem cells promotes recovery in the injured rat spinal cord. Neuroreport 2009, 20, 1559–1563. [Google Scholar]
- Yamazaki, Y.; Kanno, H.; Maeda, K.; Yoshida, T.; Kobayashi, N.; Kubo, A.; Yamaguchi, Y.; Saito, T. Engrafted VHL peptide-delivered bone marrow stromal cells promote spinal cord repair in rats. Neuroreport 2010, 21, 287–292. [Google Scholar]
- Kanno, H.; Sato, H.; Yokoyama, T.; Yoshizumi, T.; Yamada, S. The VHL tumor suppressor protein regulates tumorigenicity of U87-derived glioma stem-like cells by inhibiting the JAK/STAT signaling pathway. Int. J. Oncol 2013, 42, 881–886. [Google Scholar]
- Wong, C.E.; Paratore, C.; Dours-Zimmermann, M.T.; Rochat, A.; Pietri, T.; Suter, U.; Zimmermann, D.R.; Dufour, S.; Thiery, J.P.; Meijer, D.; et al. Neural crest-derived cells with stem cell features can be traced back to multiple lineages in the adult skin. J. Cell. Biol 2006, 175, 1005–1015. [Google Scholar]
- Hoffman, R.M. The potential of nestin-expressing hair follicle stem cells in regenerative medicine. Expert Opin. Biol. Ther 2007, 7, 289–291. [Google Scholar]
- Nishio, Y.; Koda, M.; Kamada, T.; Someya, Y.; Yoshinaga, K.; Okada, S.; Harada, H.; Okawa, A.; Moriya, H.; Yamazaki, M. The use of hemopoietic stem cells derived from human umbilical cord blood to promote restoration of spinal cord tissue and recovery of hindlimb function in adult rats. J. Neurosurg. Spine 2006, 5, 424–433. [Google Scholar]
- Barker, R.A. Winder H: Immune problems in central nervous system cell therapy. J. Am. Soc. Exp. Neuro. Ther 2004, 1, 472–481. [Google Scholar]
- Yamada, H.; Dezawa, M.; Shimazu, S.; Baba, M.; Sawada, H.; Kuroiwa, Y.; Yamamoto, I.; Kanno, H. Transfer of the von Hippel-Lindau tumor sup-pressor gene to neuronal progenitor cells in treatment for Parkinson’s disease. Ann. Neurol 2003, 54, 352–359. [Google Scholar]
- Higashida, T.; Jitsuki, S.; Kubo, A.; Mitsushima, D.; Kamiya, Y.; Kanno, H. Skin-derived precursors differentiate into dopaminergic neuronal cells in the brains of Parkinson’s disease model rats. J. Neurosurg 2010, 113, 648–655. [Google Scholar]
- Mahrour, N.; Redwine, W.B.; Florens, L.; Swanson, S.K.; Martin-Brown, S.; Bradford, W.D.; Staehling-Hampton, K.; Washburn, M.P.; Conaway, R.C.; Conaway, J.W. Characterization of Cullin-box sequences that direct recruitment of Cul2-Rbx1 and Cul5-Rbx2 modules to Elongin BC-based ubiquitin ligases. J. Biol. Chem 2008, 283, 8005–8013. [Google Scholar]
- Wakao, S.; Kitada, M.; Kuroda, Y.; Shigemoto, T.; Matsuse, D.; Akashi, H.; Tanimura, Y.; Tsuchiyama, K.; Kikuchi, T.; Goda, M.; et al. Multilineage-differentiating stress-enduring (Muse) cells are a primary source of induced pluripotent stem cells in human fibroblasts. Proc. Natl. Acad. Sci. USA 2011, 108, 9875–9880. [Google Scholar]

© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kanno, H.; Kubo, A.; Yoshizumi, T.; Mikami, T.; Maegawa, J. Isolation of Multipotent Nestin-Expressing Stem Cells Derived from the Epidermis of Elderly Humans and TAT-VHL Peptide-Mediated Neuronal Differentiation of These Cells. Int. J. Mol. Sci. 2013, 14, 9604-9617. https://doi.org/10.3390/ijms14059604
Kanno H, Kubo A, Yoshizumi T, Mikami T, Maegawa J. Isolation of Multipotent Nestin-Expressing Stem Cells Derived from the Epidermis of Elderly Humans and TAT-VHL Peptide-Mediated Neuronal Differentiation of These Cells. International Journal of Molecular Sciences. 2013; 14(5):9604-9617. https://doi.org/10.3390/ijms14059604
Chicago/Turabian StyleKanno, Hiroshi, Atsuhiko Kubo, Tetsuya Yoshizumi, Taro Mikami, and Jiro Maegawa. 2013. "Isolation of Multipotent Nestin-Expressing Stem Cells Derived from the Epidermis of Elderly Humans and TAT-VHL Peptide-Mediated Neuronal Differentiation of These Cells" International Journal of Molecular Sciences 14, no. 5: 9604-9617. https://doi.org/10.3390/ijms14059604