Two Volatile Organic Compounds Trigger Plant Self-Defense against a Bacterial Pathogen and a Sucking Insect in Cucumber under Open Field Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
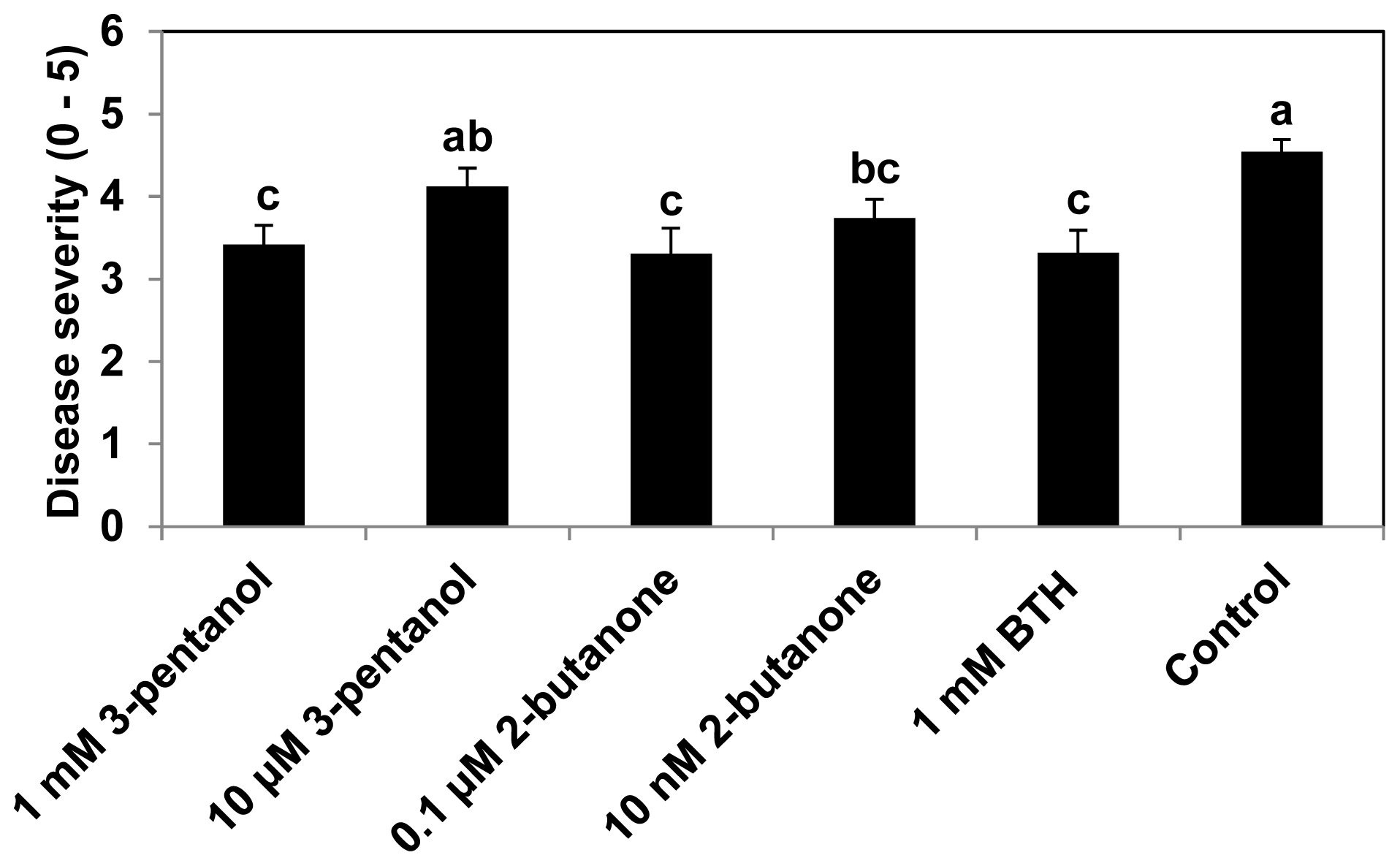
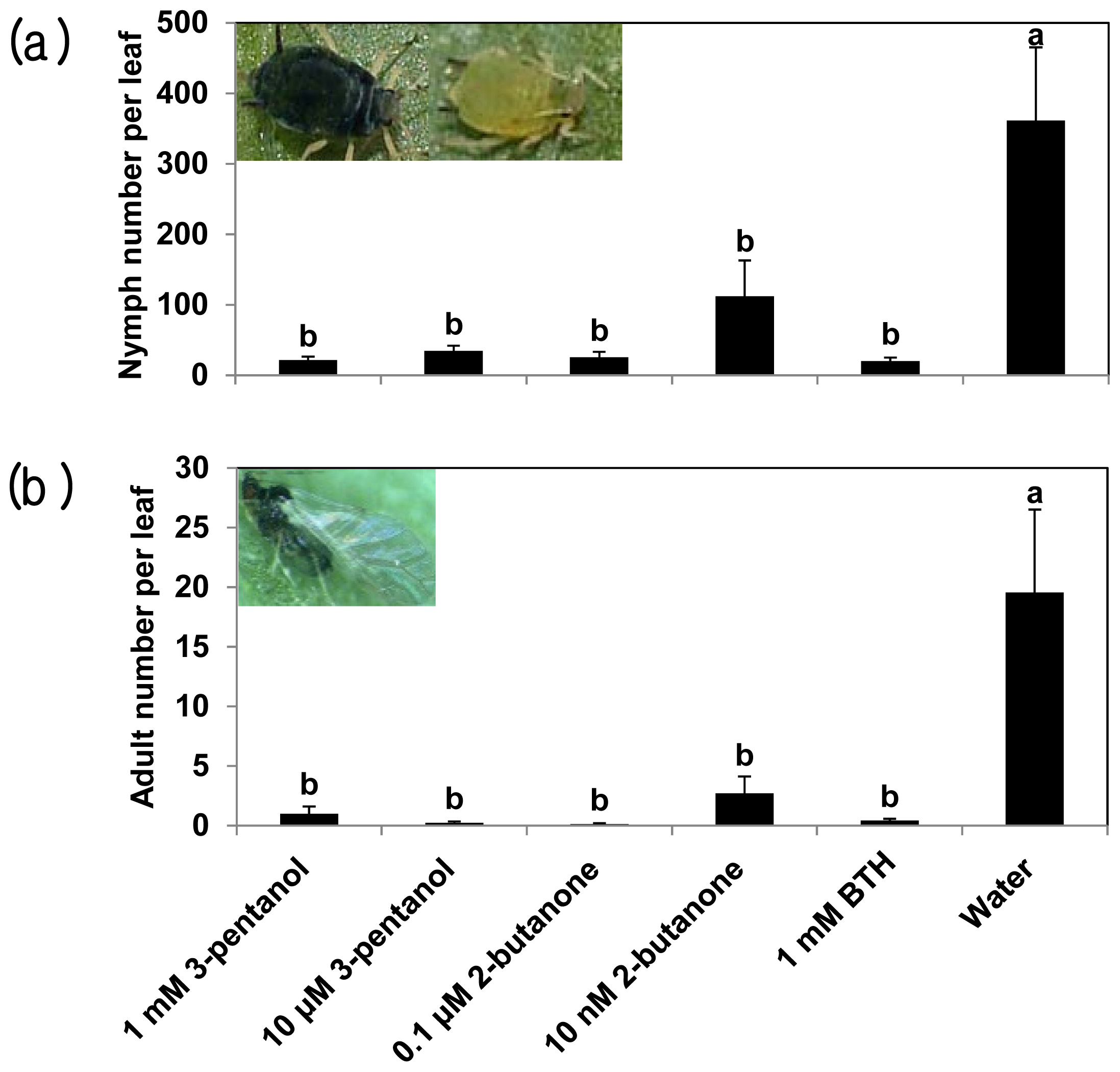
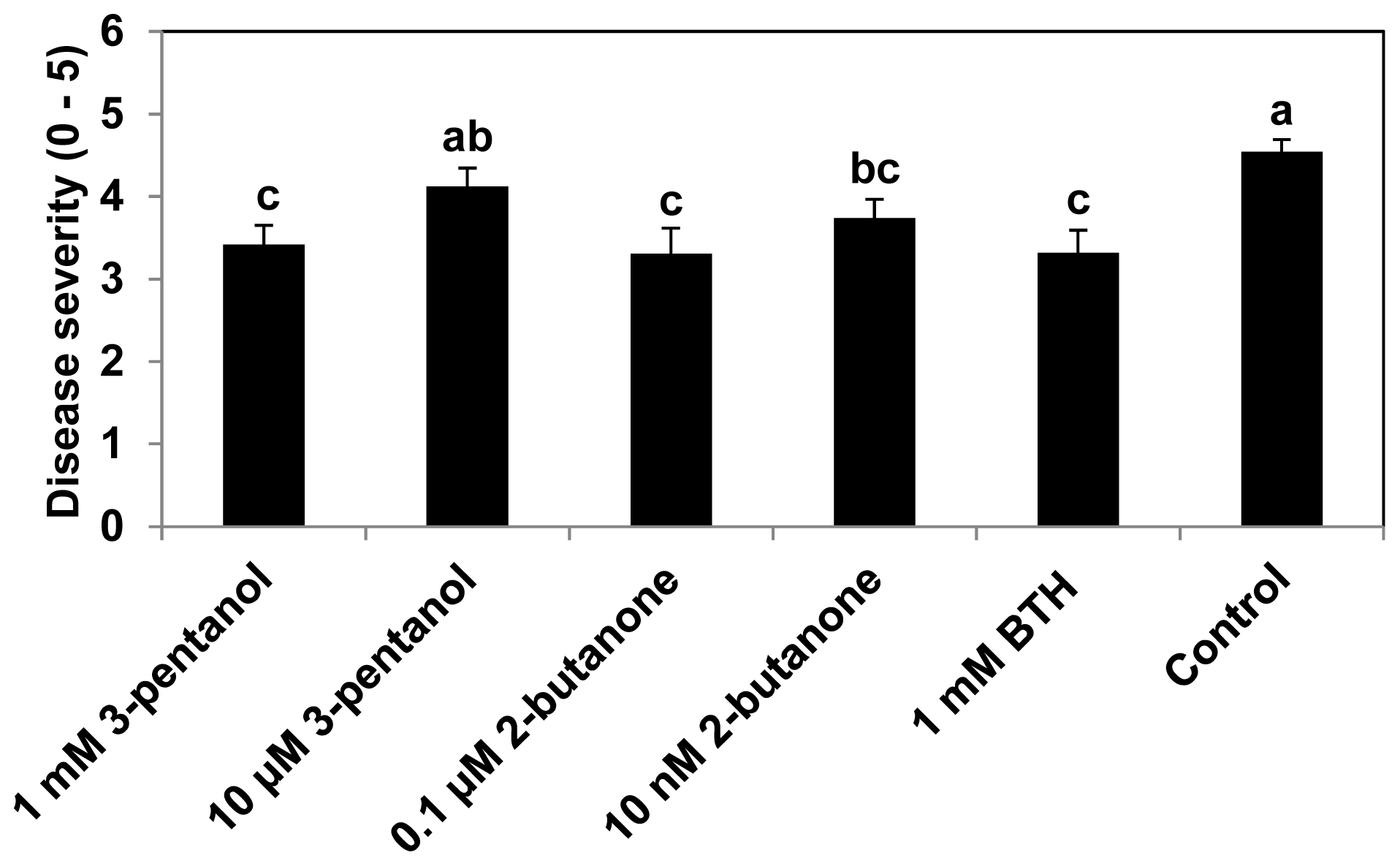
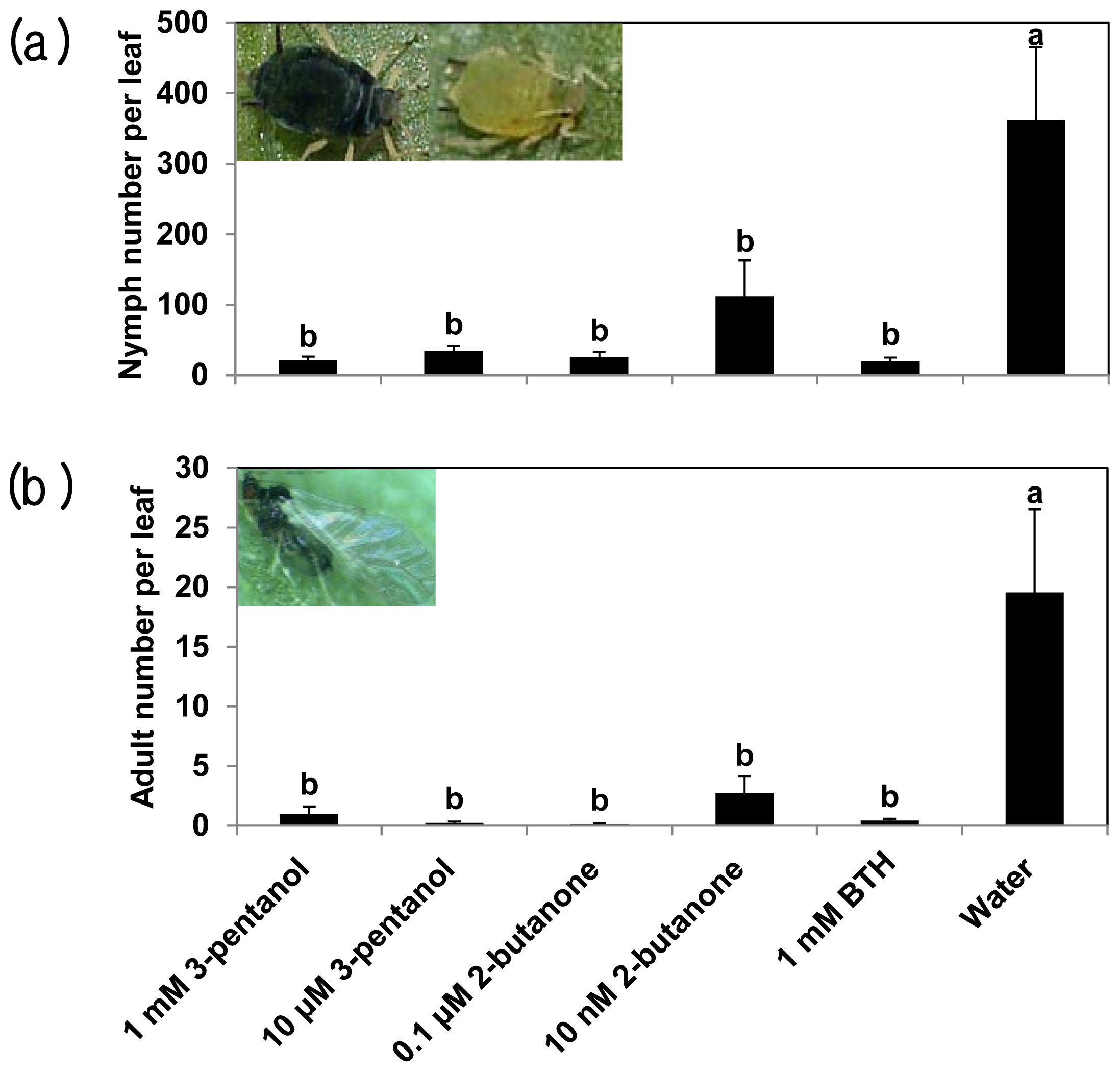
2.1. 3-Pentanol and 2-Butanone-Elicit Induced Resistance against Pseudomonas syringae and Aphids
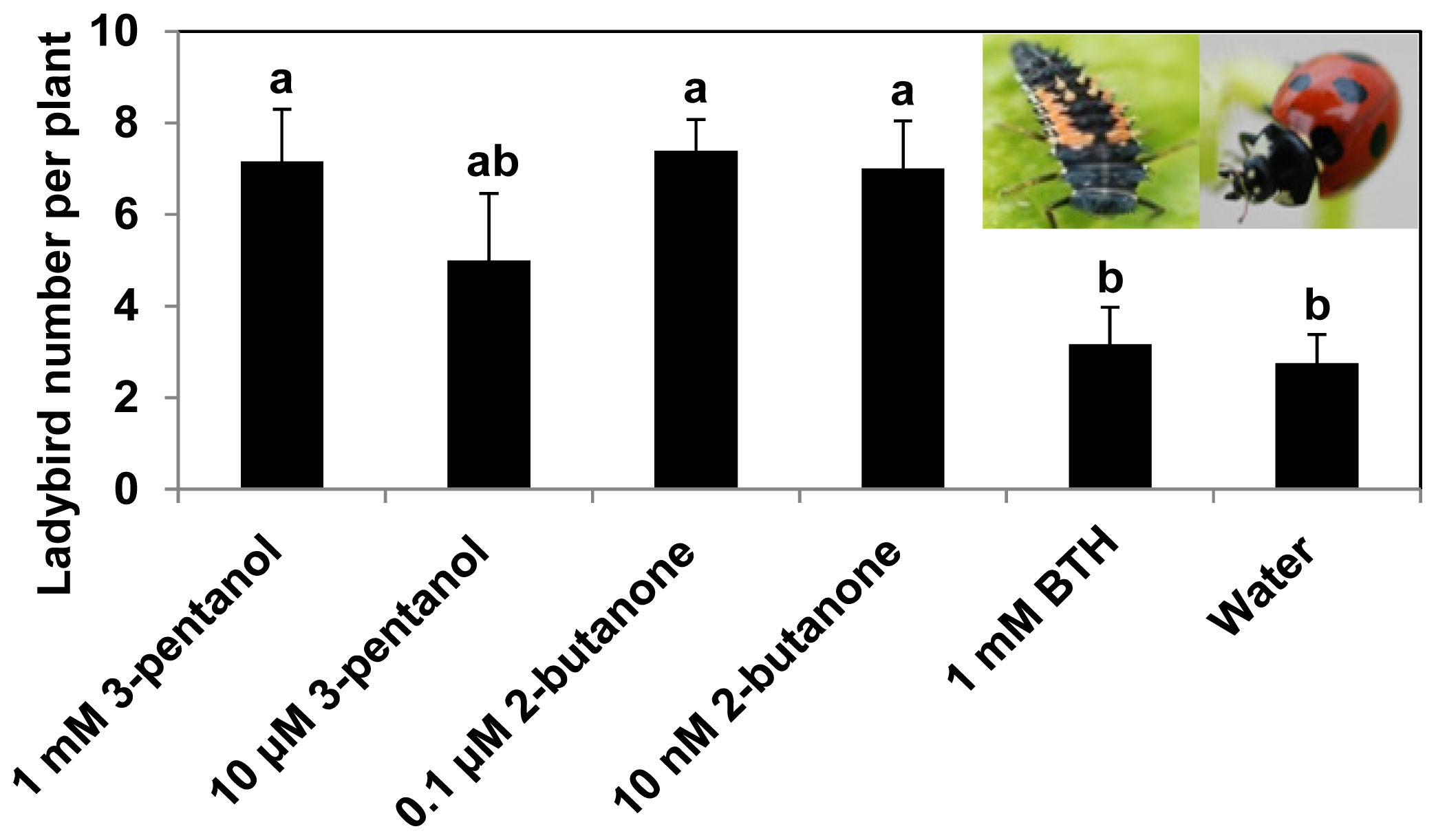
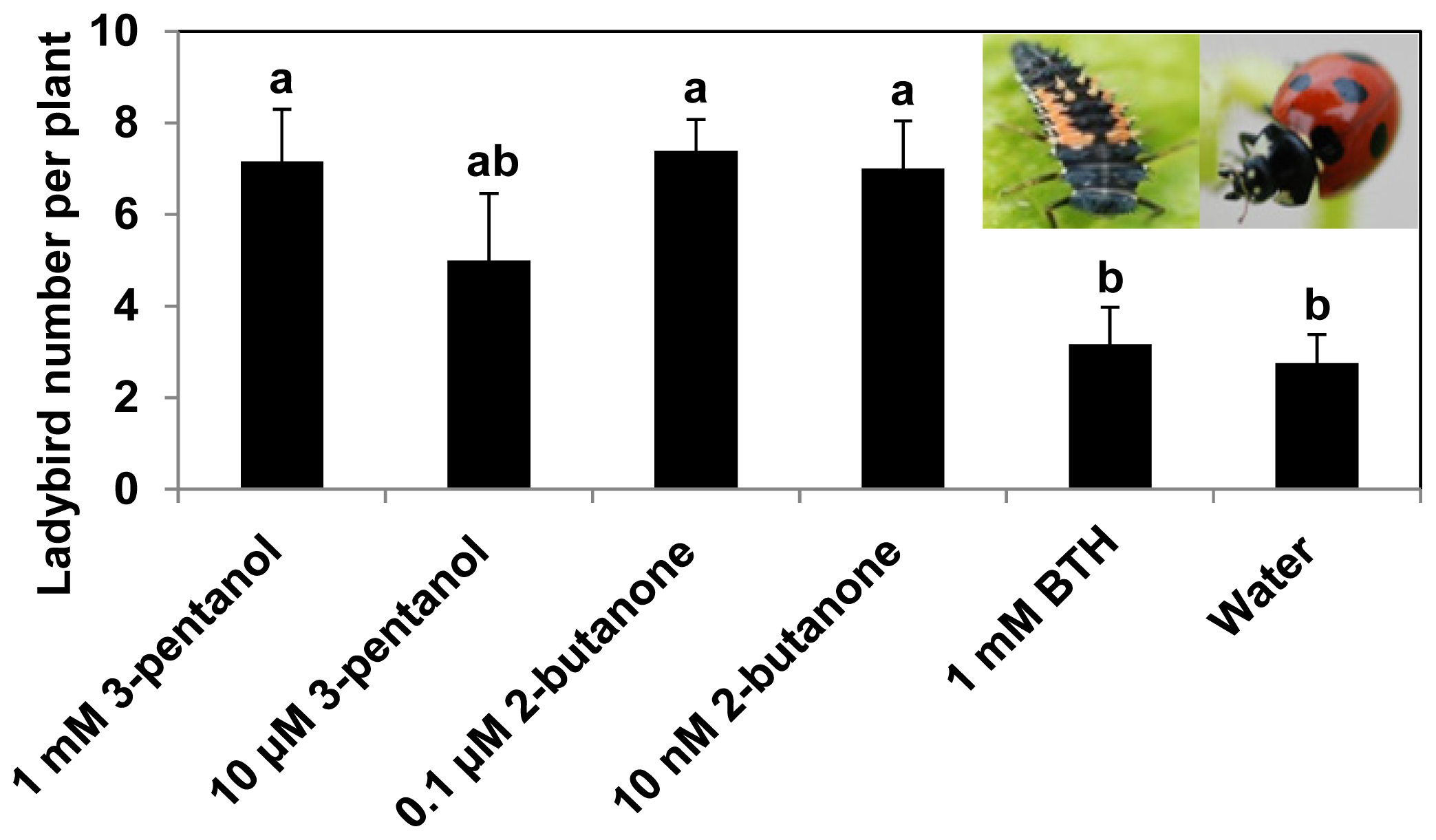
2.2. 3-Pentanol and 2-Butanone Treatment Increase the Numbers of Ladybird Beetles
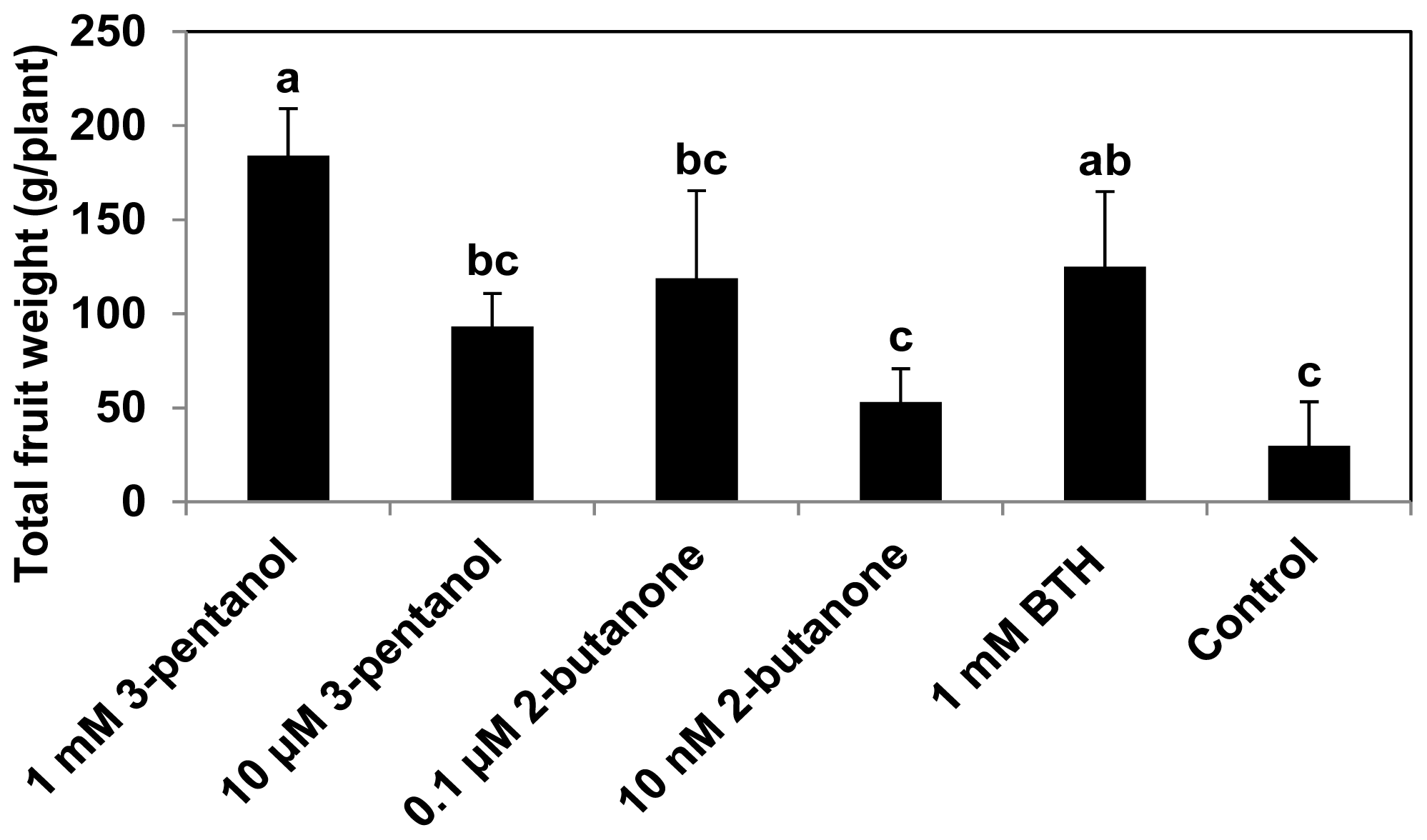
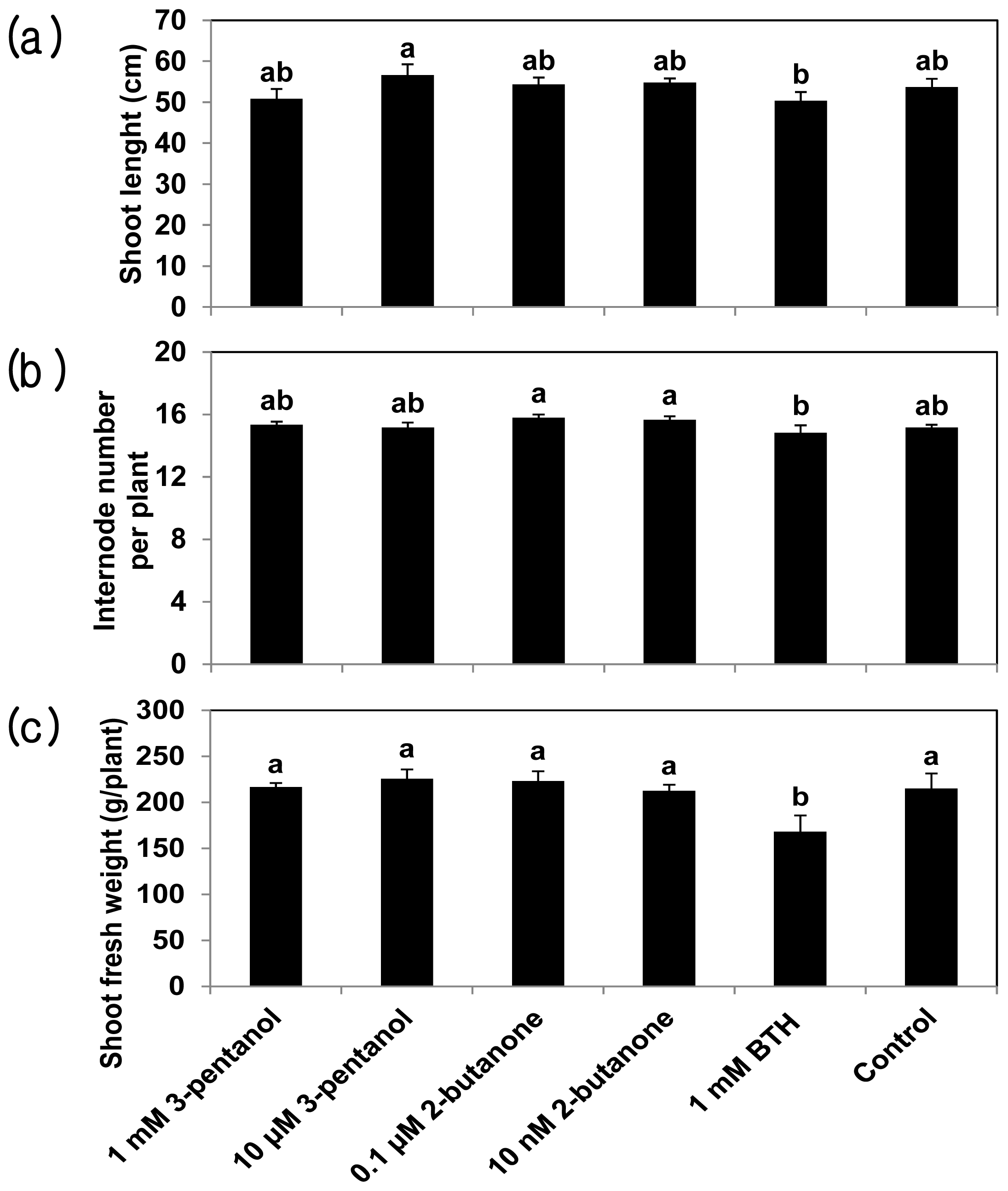
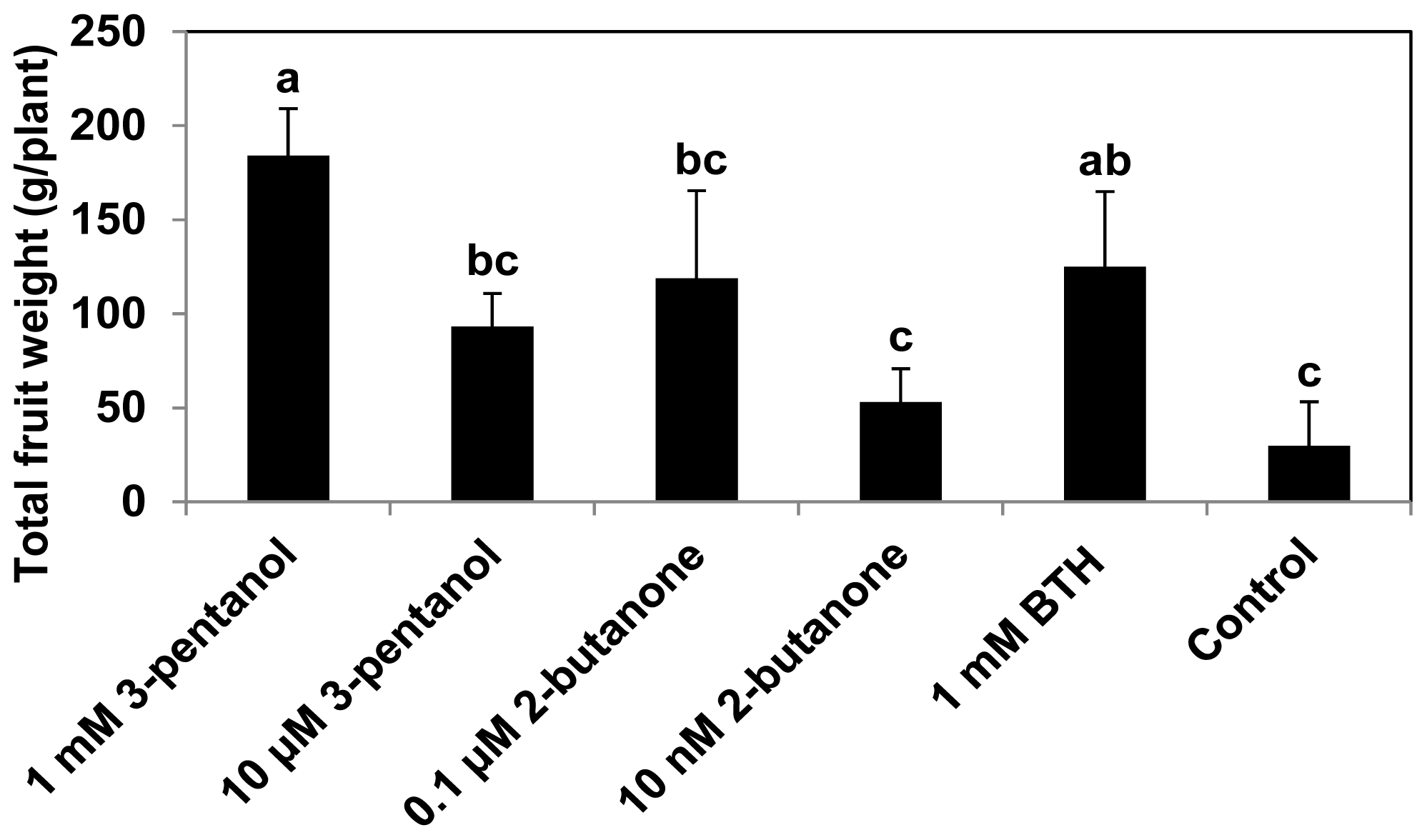
2.3. 3-Pentanol and 2-Butanone Do Not Affect Plant Growth but Increase Fruit Yield
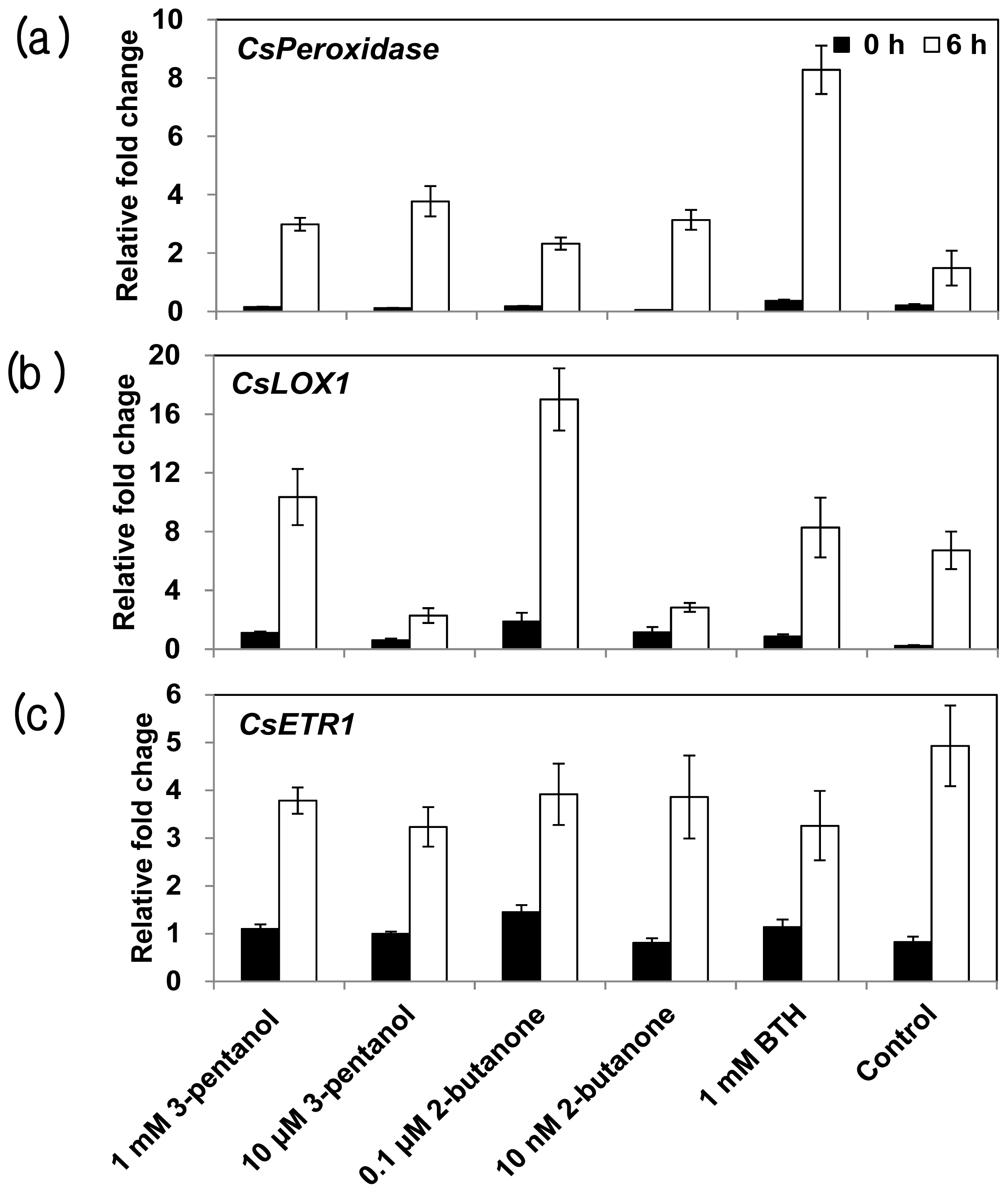
2.4. Expression of Defense-Related Genes in Response to 3-Pentanol and 2-Butanone
3. Experimental Section
3.1. Plant Growth and Chemical Treatment
3.2. Assessment of Angular Leaf Spot Disease and Aphid Infestation
3.3. Measurement of Plant Growth Parameters
3.4. RT-PCR and Quantitative RT-PCR
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Schneider, M.; Schweizer, P.; Meuwly, P.; Métraux, J. Systemic acquired resistance in plants. Int. Rev. Cytol 1996, 168, 303–340. [Google Scholar]
- Ross, A.F. Systemic acquired resistance induced by localized virus infections in plants. Virology 1961, 14, 340–358. [Google Scholar]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.Y.; Hunt, M.D. Systemic acquired resistance. Plant Cell 1996, 8, 1809–1819. [Google Scholar]
- Kloepper, J.W.; Ryu, C.M.; Zhang, S. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar]
- Van Loon, L.; Bakker, P.; Pieterse, C. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol 1998, 36, 453–483. [Google Scholar]
- Ryu, C.M.; Farag, M.A.; Hu, C.H.; Reddy, M.S.; Kloepper, J.W.; Paré, P.W. Bacterial volatiles induce systemic resistance in Arabidopsis. Plant Physiol 2004, 134, 1017–1026. [Google Scholar]
- Kwon, Y.S.; Ryu, C.M.; Lee, S.; Park, H.B.; Han, K.S.; Lee, J.H.; Lee, K.; Chung, W.S.; Jeong, M.J.; Kim, H.K. Proteome analysis of Arabidopsis seedlings exposed to bacterial volatiles. Planta 2010, 232, 1355–1370. [Google Scholar]
- Farag, M.A.; Ryu, C.M.; Sumner, L.W.; Paré, P.W. GC–MS SPME profiling of rhizobacterial volatiles reveals prospective inducers of growth promotion and induced systemic resistance in plants. Phytochemistry 2006, 67, 2262–2268. [Google Scholar]
- Han, S.H.; Lee, S.J.; Moon, J.H.; Park, K.H.; Yang, K.Y.; Cho, B.H.; Kim, K.Y.; Kim, Y.W.; Lee, M.C.; Anderson, A.J. GacS-dependent production of 2R, 3R-butanediol by Pseudomonas chlororaphis O6 is a major determinant for eliciting systemic resistance against Erwinia carotovora but not against Pseudomonas syringae pv. tabaci in tobacco. Mol. Plant Microbe Interact 2006, 19, 924–930. [Google Scholar]
- Cortes-Barco, A.; Goodwin, P.; Hsiang, T. Comparison of induced resistance activated by benzothiadiazole,(2R,3R)-butanediol and an isoparaffin mixture against anthracnose of Nicotiana benthamiana. Plant Pathol 2010, 59, 643–653. [Google Scholar]
- Cortes-Barco, A.; Hsiang, T.; Goodwin, P. Induced systemic resistance against three foliar diseases of Agrostis stolonifera by (2R,3R)-butanediol or an isoparaffin mixture. Ann. Appl. Biol 2010, 157, 179–189. [Google Scholar]
- Tally, A.; Oostendorp, M.; Lawton, K.; Staub, T.; Bassi, B.; Agrawal, A.; Tuzun, S.; Bent, E. Commercial Development of Elicitors of Induced Resistance to Pathogens; APS press: St. Paul, MN, USA, 1999; pp. 357–369. [Google Scholar]
- Molina, A.; Hunt, M.D.; Ryals, J.A. Impaired fungicide activity in plants blocked in disease resistance signal transduction. Plant Cell 1998, 10, 1903–1914. [Google Scholar]
- Heil, M.; Hilpert, A.; Kaiser, W.; Linsenmair, K.E. Reduced growth and seed set following chemical induction of pathogen defence: Does systemic acquired resistance (SAR) incur allocation costs? J. Ecol 2001, 88, 645–654. [Google Scholar]
- Van Hulten, M.; Pelser, M.; van Loon, L.; Pieterse, C.M.J.; Ton, J. Costs and benefits of priming for defense in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 5602–5607. [Google Scholar]
- Heil, M. The ecological concept of costs of induced systemic resistance (ISR). Eur. J. Plant Pathol 2001, 107, 137–146. [Google Scholar]
- Yang, J.W.; Yu, S.H.; Ryu, C.M. Priming of defense-related genes confers root-colonizing bacilli-elicited induced systemic resistance in pepper. Plant Pathol. J 2009, 25, 389–399. [Google Scholar]
- Funes, H.; Zerba, E.; González Audino, P. Comparison of three types of traps baited with sexual pheromones for ambrosia beetle Megaplatypus mutatus (Coleoptera: Platypodinae) in poplar plantations. J. Econ. Entomol 2009, 102, 1546–1550. [Google Scholar]
- Manrique, G.; Vitta, A.C.R.; Ferreira, R.A.; Zani, C.L.; Unelius, C.R.; Lazzari, C.R.; Diotaiuti, L.; Lorenzo, M.G. Chemical communication in Chagas disease vectors. Source, identity, and potential function of volatiles released by the metasternal and Brindley’s glands of Triatoma infestans adults. J. Chem Ecol 2006, 32, 2035–2052. [Google Scholar]
- Vitta, A.C.R.; Lorenzo, M.G. Copulation and mate guarding behavior in Triatoma brasiliensis (Hemiptera: Reduviidae). J. Med. Entomol 2009, 46, 789–795. [Google Scholar]
- Funes, H.; Griffo, R.; Zerba, E.; Gonzalez-Audino, P. Mating disruption of the ambrosia beetle Megaplatypus mutatus in poplar and hazelnut plantations using reservoir systems for pheromones. Entomol. Exp. Appl 2011, 139, 226–234. [Google Scholar]
- Bukovinszky, T.; Gols, R.; Posthumus, M.; Vet, L.; Van Lenteren, J. Variation in plant volatiles and attraction of the parasitoiddiadegma semiclausum (Hellen). J. Chem. Ecol 2005, 31, 461–480. [Google Scholar]
- Gols, R.; Bullock, J.M.; Dicke, M.; Bukovinszky, T.; Harvey, J.A. Smelling the wood from the trees: Non-linear parasitoid responses to volatile attractants produced by wild and cultivated cabbage. J. Chem. Ecol 2011, 37, 795–807. [Google Scholar]
- Zhuge, P.P.; Luo, S.L.; Wang, M.Q.; Zhang, G. Electrophysiological responses of Batocera horsfieldi (Hope) adults to plant volatiles. J. Appl. Entomol 2010, 134, 600–607. [Google Scholar]
- Jollivet, N.; Bézenger, M.C.; Vayssier, Y.; Belin, J.M. Production of volatile compounds in liquid cultures by six strains of coryneform bacteria. Appl. Microbiol. Biotechnol 1992, 36, 790–794. [Google Scholar]
- Wheatley, R. The consequences of volatile organic compound mediated bacterial and fungal interactions. Antonie van Leeuwenhoek 2002, 81, 357–364. [Google Scholar]
- Wilkins, K. Volatile metabolites from actinomycetes. Chemosphere 1996, 32, 1427–1434. [Google Scholar]
- Wheatley, R.; Hackett, C.; Bruce, A.; Kundzewicz, A. Effect of substrate composition on production of volatile organic compounds from Trichoderma spp. Inhibitory to wood decay fungi. Int. Biodeterior. Biodegrad 1997, 39, 199–205. [Google Scholar]
- Dicke, M.; Baldwin, I.T. The evolutionary context for herbivore-induced plant volatiles: Beyond the “cry for help”. Trends Plant Sci 2010, 15, 167–175. [Google Scholar]
- Rudrappa, T.; Biedrzycki, M.L.; Kunjeti, S.G.; Donofrio, N.M.; Czymmek, K.J.; Bais, H.P. The rhizobacterial elicitor acetoin induces systemic resistance in Arabidopsis thaliana. Commun. Integr. Biol 2010, 3, 130–138. [Google Scholar]
- Lee, B.; Lee, S.; Ryu, C.M. Foliar aphid feeding recruits rhizosphere bacteria and primes plant immunity against pathogenic and non-pathogenic bacteria in pepper. Ann. Bot 2012, 110, 281–290. [Google Scholar]
- Görlach, J.; Volrath, S.; Knauf-Beiter, G.; Hengy, G.; Beckhove, U.; Kogel, K.H.; Oostendorp, M.; Staub, T.; Ward, E.; Kessmann, H. Benzothiadiazole, a novel class of inducers of systemic acquired resistance, activates gene expression and disease resistance in wheat. Plant Cell 1996, 8, 629–643. [Google Scholar]
- Ahmad, S.; Veyrat, N.; Gordon-Weeks, R.; Zhang, Y.; Martin, J.; Smart, L.; Glauser, G.; Erb, M.; Flors, V.; Frey, M. Benzoxazinoid metabolites regulate innate immunity against aphids and fungi in maize. Plant Physiol 2011, 157, 317–327. [Google Scholar]
- Van Lenteren, J.C. A greenhouse without pesticides: Fact or fantasy? Crop Prot 2000, 19, 375–384. [Google Scholar]
- Snyder, W.E.; Clevenger, G.M.; Eigenbrode, S.D. Intraguild predation and successful invasion by introduced ladybird beetles. Oecologia 2004, 140, 559–565. [Google Scholar]
- Harmel, N.; Almohamad, R.; Fauconnier, M.L.; Du Jardin, P.; Verheggen, F.; Marlier, M.; Haubruge, E.; Francis, F. Role of terpenes from aphid-infested potato on searching and oviposition behavior of Episyrphus balteatus. Insect Sci 2007, 14, 57–63. [Google Scholar]
- Heil, M.; Ton, J. Long-distance signalling in plant defence. Trends Plant Sci 2008, 13, 264–272. [Google Scholar]
- Yang, J.W.; Yi, H.S.; Kim, H.; Lee, B.; Lee, S.; Ghim, S.Y.; Ryu, C.M. Whitefly infestation of pepper plants elicits defence responses against bacterial pathogens in leaves and roots and changes the below-ground microflora. J. Ecol 2011, 99, 46–56. [Google Scholar]
- Cipollini, D.; Heil, M. Costs and benefits of induced resistance to herbivores and pathogens in plants. Plant Sci. Rev 2010, 5, 1–25. [Google Scholar]
- Cools, H.J.; Ishii, H. Pre-treatment of cucumber plants with acibenzolar-S-methyl systemically primes a phenylalanine ammonia lyase gene (PAL1) for enhanced expression upon attack with a pathogenic fungus. Physiol. Mol. Plant Pathol 2002, 61, 273–280. [Google Scholar]
- Shoresh, M.; Yedidia, I.; Chet, I. Involvement of jasmonic Acid/ethylene signaling pathway in the systemic resistance induced in cucumber by Trichoderma asperellumT203. Phytopathology 2004, 95, 76–84. [Google Scholar]
- Conrath, U.; Beckers, G.J.M.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Newman, M.A.; Pieterse, C.M.J.; Poinssot, B.; Pozo, M.J. Priming: Getting ready for battle. Mol. Plant Microbe Interact 2006, 19, 1062–1071. [Google Scholar]
- Chen, S.; Dai, X.; Qiang, S.; Tang, Y. Effect of a nonhost-selective toxin from Alternaria alternata on chloroplast-electron transfer activity in Eupatorium adenophorum. Plant Pathol 2005, 54, 671–677. [Google Scholar]
- Weber, H. Fatty acid-derived signals in plants. Trends Plant Sci 2002, 7, 217–224. [Google Scholar]
- Turner, J.G.; Ellis, C.; Devoto, A. The jasmonate signal pathway. Plant Cell 2002, 14, S153–S164. [Google Scholar]
- Kappers, I.F.; Verstappen, F.W.A.; Luckerhoff, L.L.P.; Bouwmeester, H.J.; Dicke, M. Genetic variation in jasmonic acid-and spider mite-induced plant volatile emission of cucumber accessions and attraction of the predator Phytoseiulus persimilis. J. Chem. Ecol 2010, 36, 500–512. [Google Scholar]
- Mercke, P.; Kappers, I.F.; Verstappen, F.W.A.; Vorst, O.; Dicke, M.; Bouwmeester, H.J. Combined transcript and metabolite analysis reveals genes involved in spider mite induced volatile formation in cucumber plants. Plant Physiol 2004, 135, 2012–2024. [Google Scholar]
- Pieterse, C.M.J.; Leon-Reyes, A.; van der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol 2009, 5, 308–316. [Google Scholar]
- Song, G.C.; Chio, H.K.; Ryu, C.M. The folate precursor para-aminobenzoic acid elicits induced resistance against Cucumber mosaic virus and Xanthomonas axonopodis. Ann. Bot 2013, 111, 925–934. [Google Scholar]
- Mathooko, F.M.; Mwaniki, M.W.; Nakatsuka, A.; Shiomi, S.; Kubo, Y.; Inaba, A.; Nakamura, R. Expression characteristics of CS-ACS1, CS-ACS2 and CS-ACS3, three members of the 1-aminocyclopropane-1-carboxylate synthase gene family in cucumber (Cucumis sativus L.) fruit under carbon dioxide stress. Plant Cell Physiol 1999, 40, 164–172. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Song, G.C.; Ryu, C.-M. Two Volatile Organic Compounds Trigger Plant Self-Defense against a Bacterial Pathogen and a Sucking Insect in Cucumber under Open Field Conditions. Int. J. Mol. Sci. 2013, 14, 9803-9819. https://doi.org/10.3390/ijms14059803
Song GC, Ryu C-M. Two Volatile Organic Compounds Trigger Plant Self-Defense against a Bacterial Pathogen and a Sucking Insect in Cucumber under Open Field Conditions. International Journal of Molecular Sciences. 2013; 14(5):9803-9819. https://doi.org/10.3390/ijms14059803
Chicago/Turabian StyleSong, Geun Cheol, and Choong-Min Ryu. 2013. "Two Volatile Organic Compounds Trigger Plant Self-Defense against a Bacterial Pathogen and a Sucking Insect in Cucumber under Open Field Conditions" International Journal of Molecular Sciences 14, no. 5: 9803-9819. https://doi.org/10.3390/ijms14059803