Increased Susceptibility of Radiation-Induced Intestinal Apoptosis in SMP30 KO Mice

 ,
, 
Abstract
:1. Introduction
2. Results
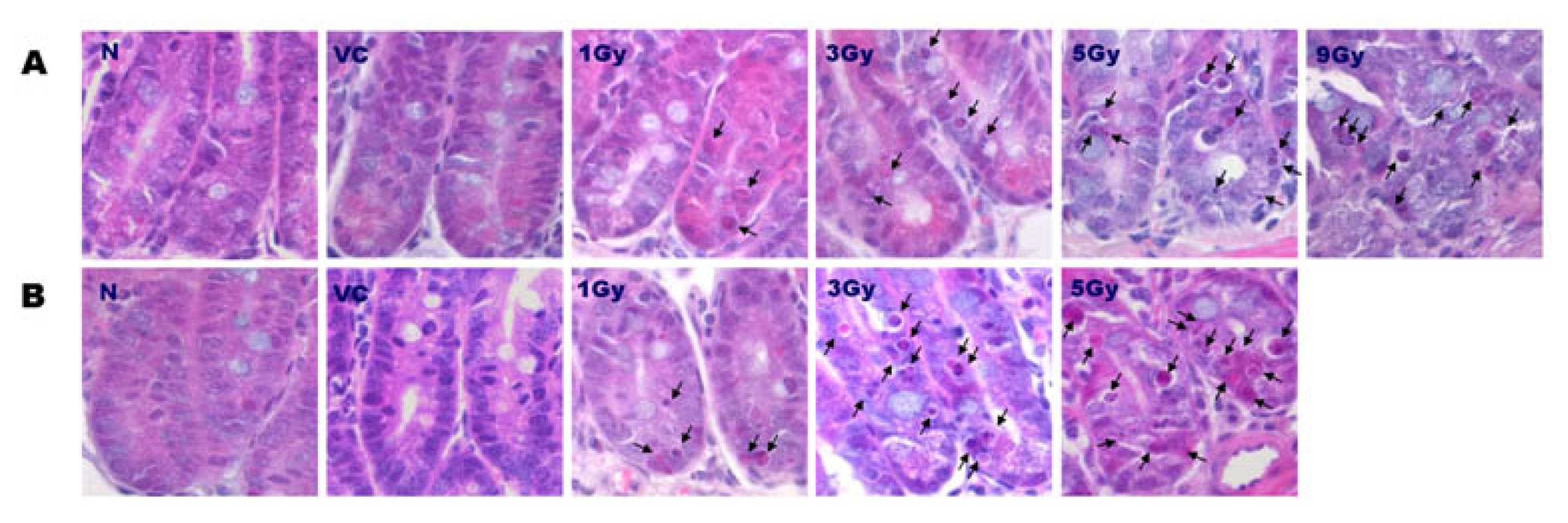
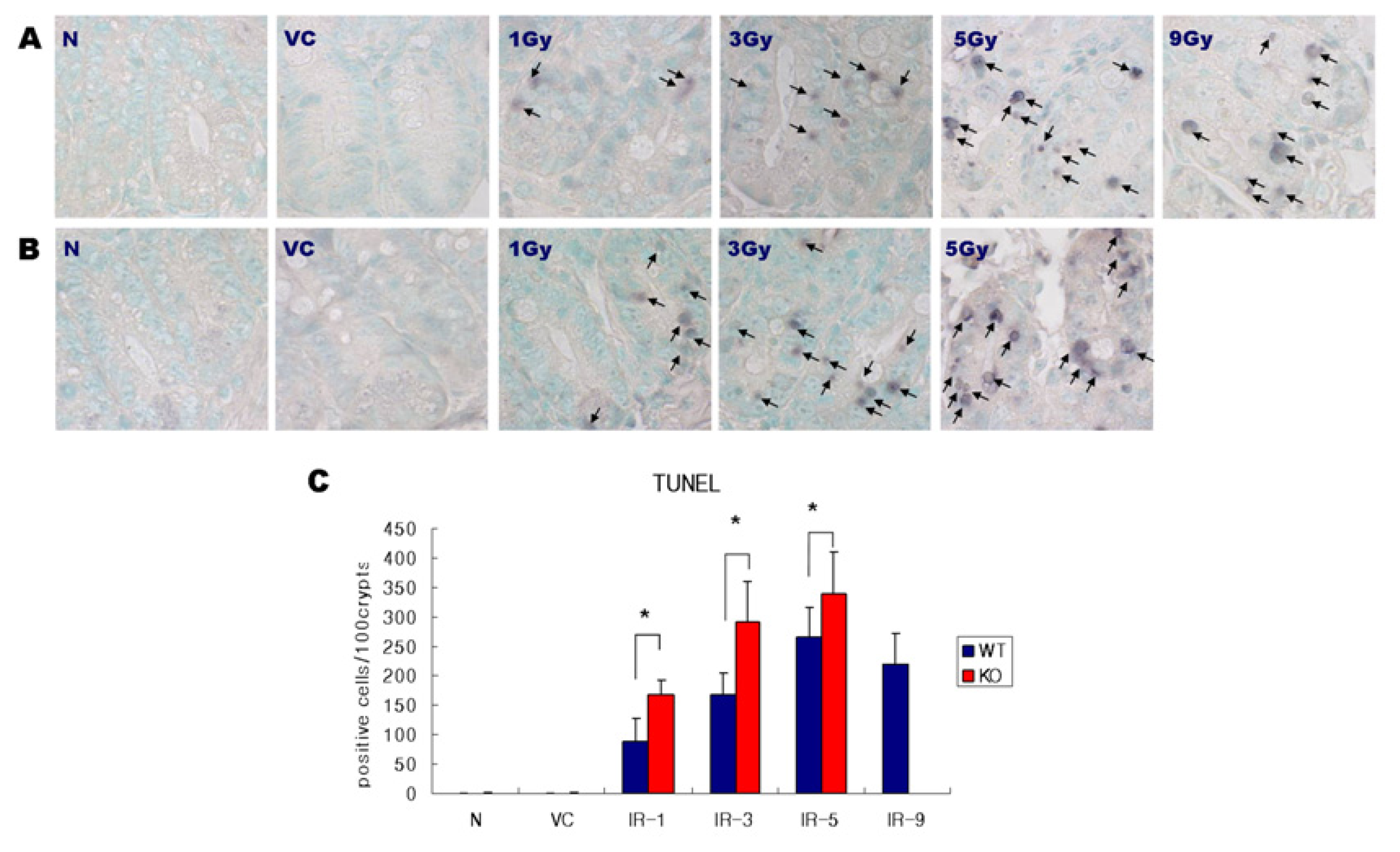
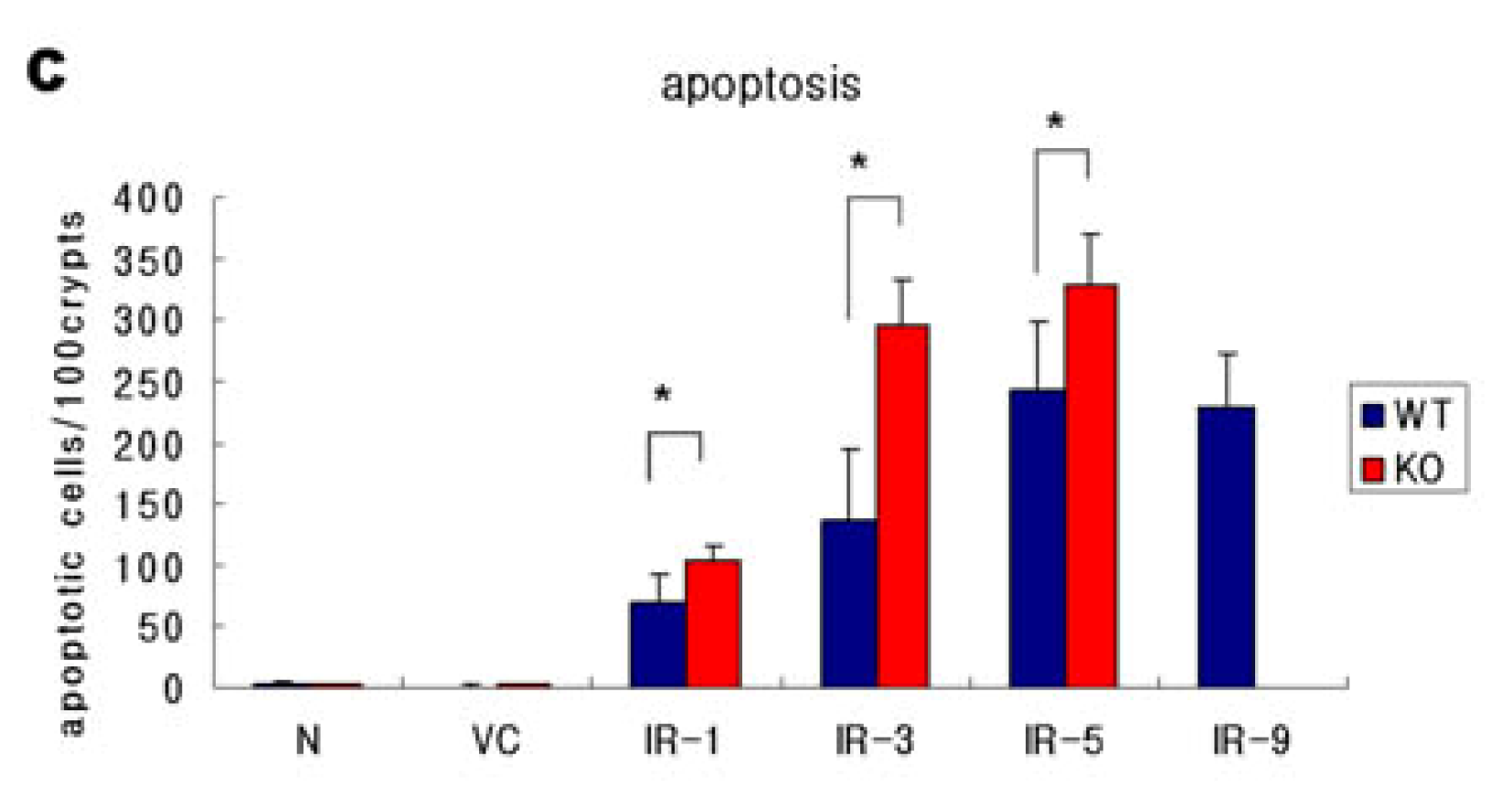
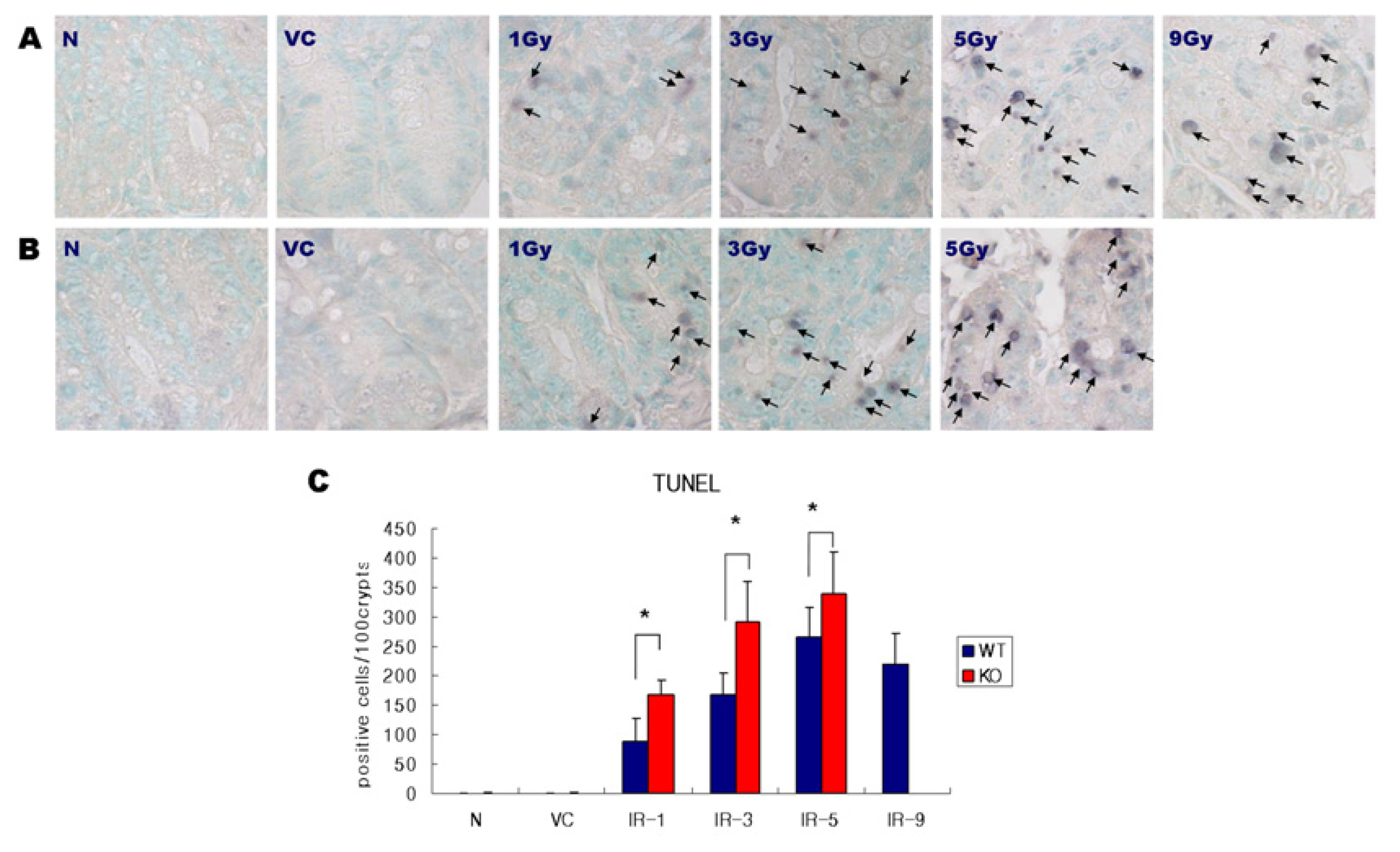
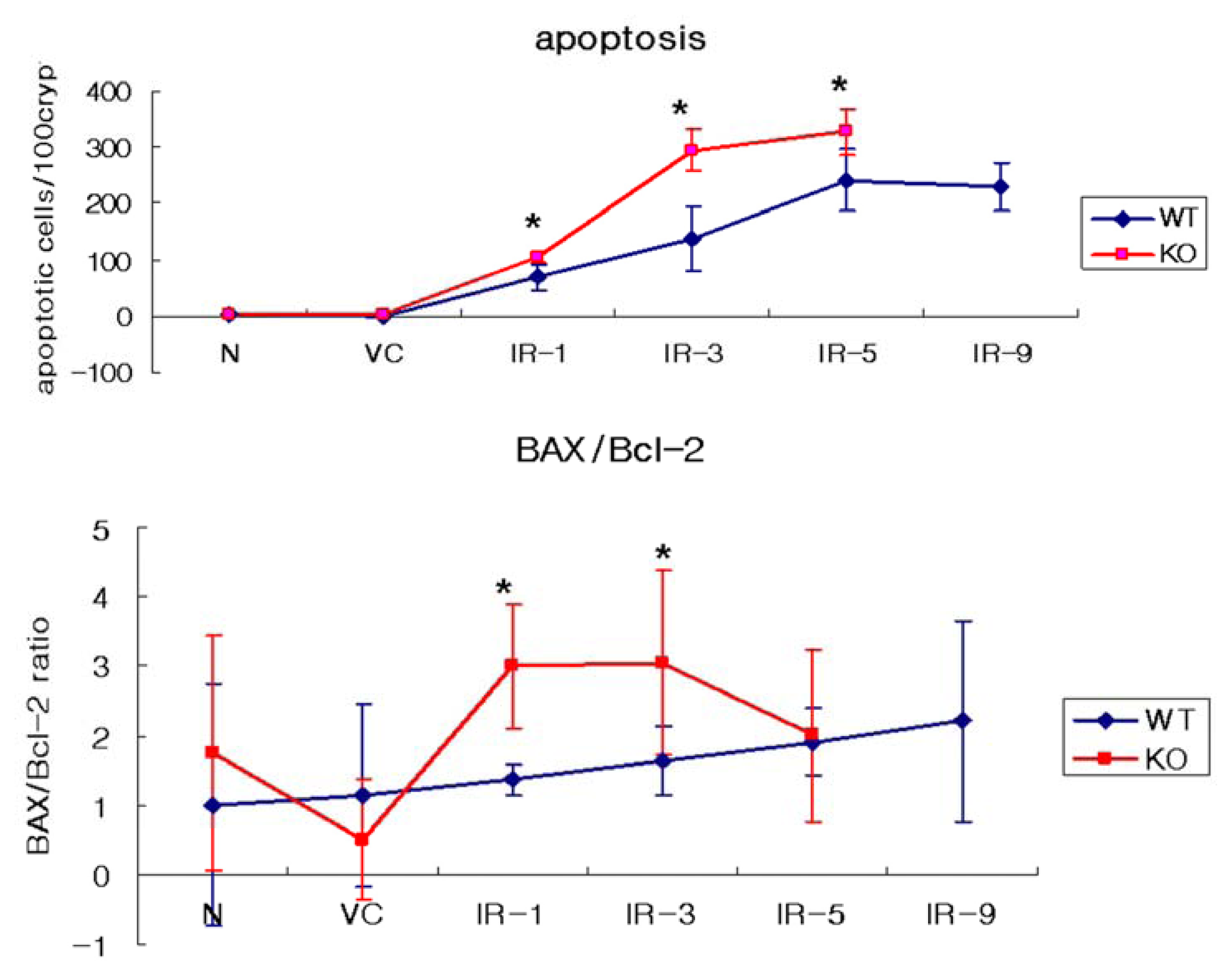
2.1. Distribution of Apoptosis
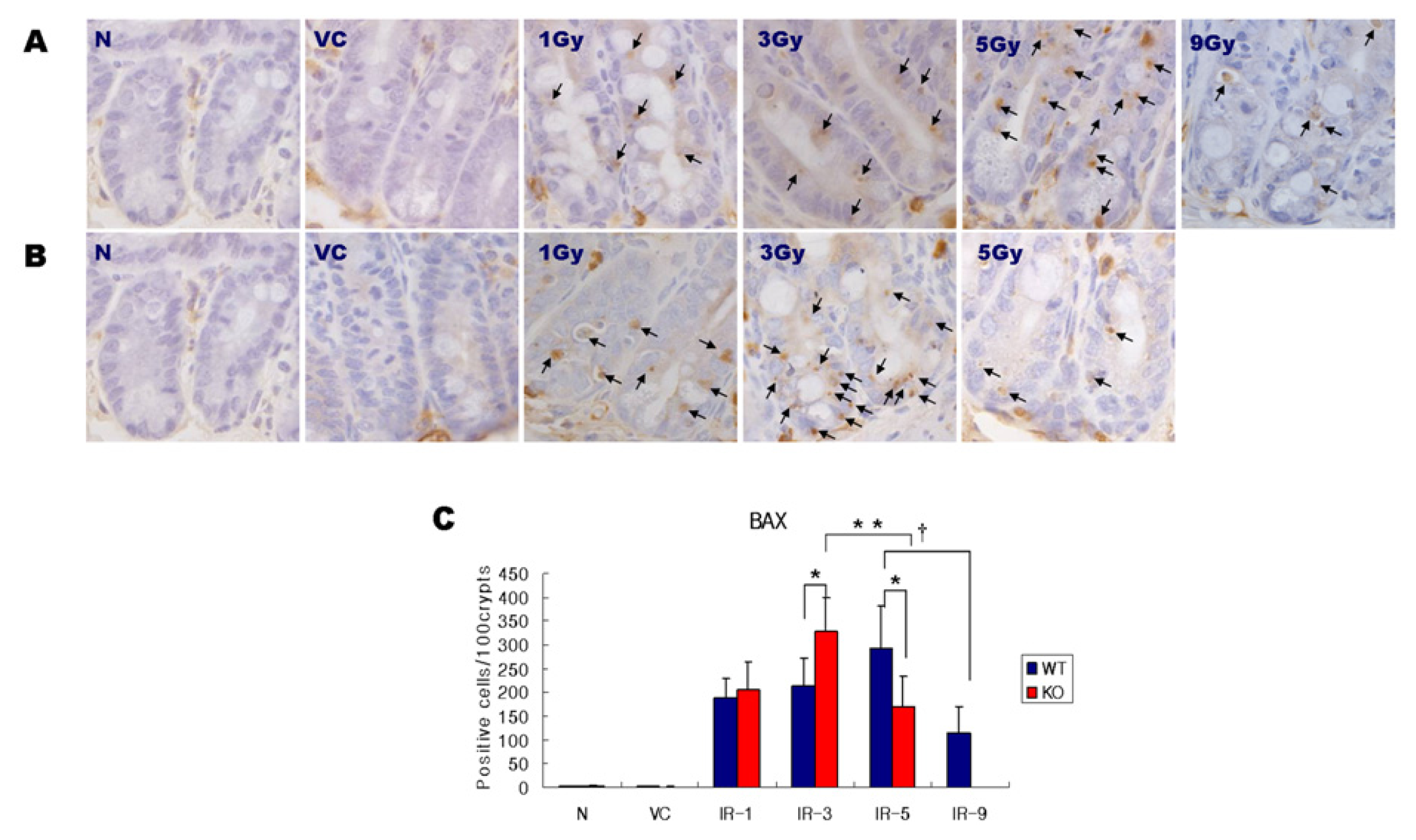
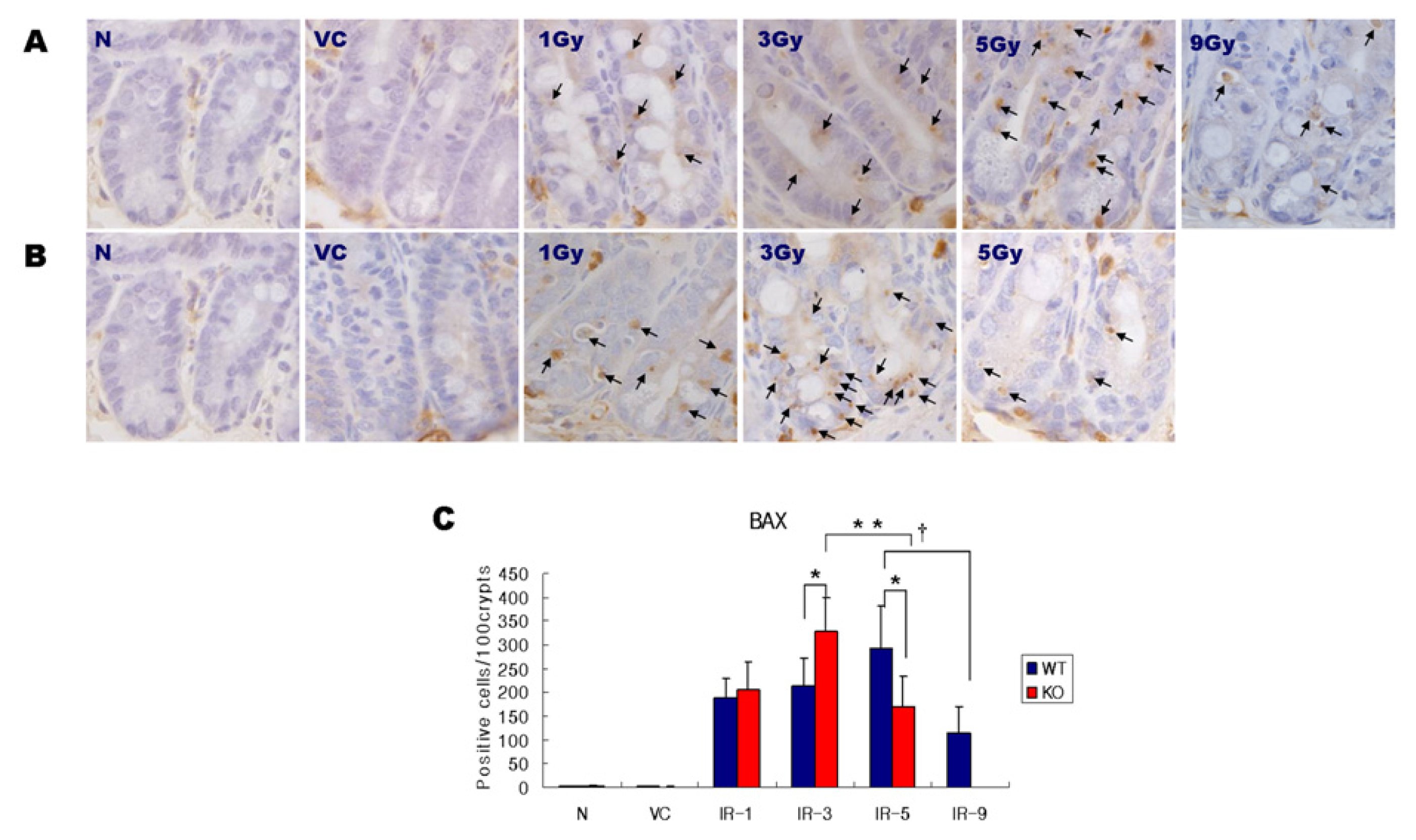
2.2. Expression of BAX
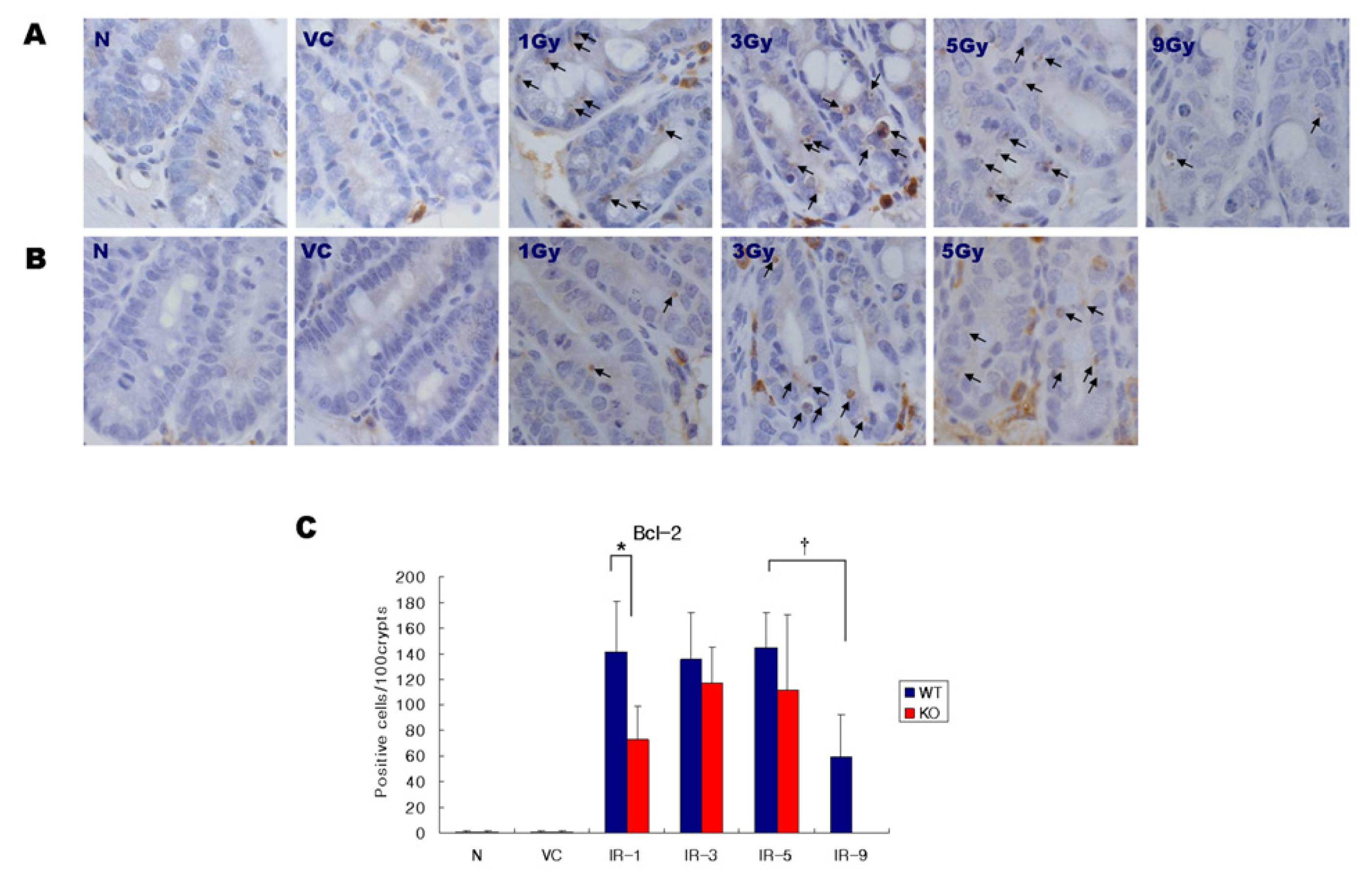
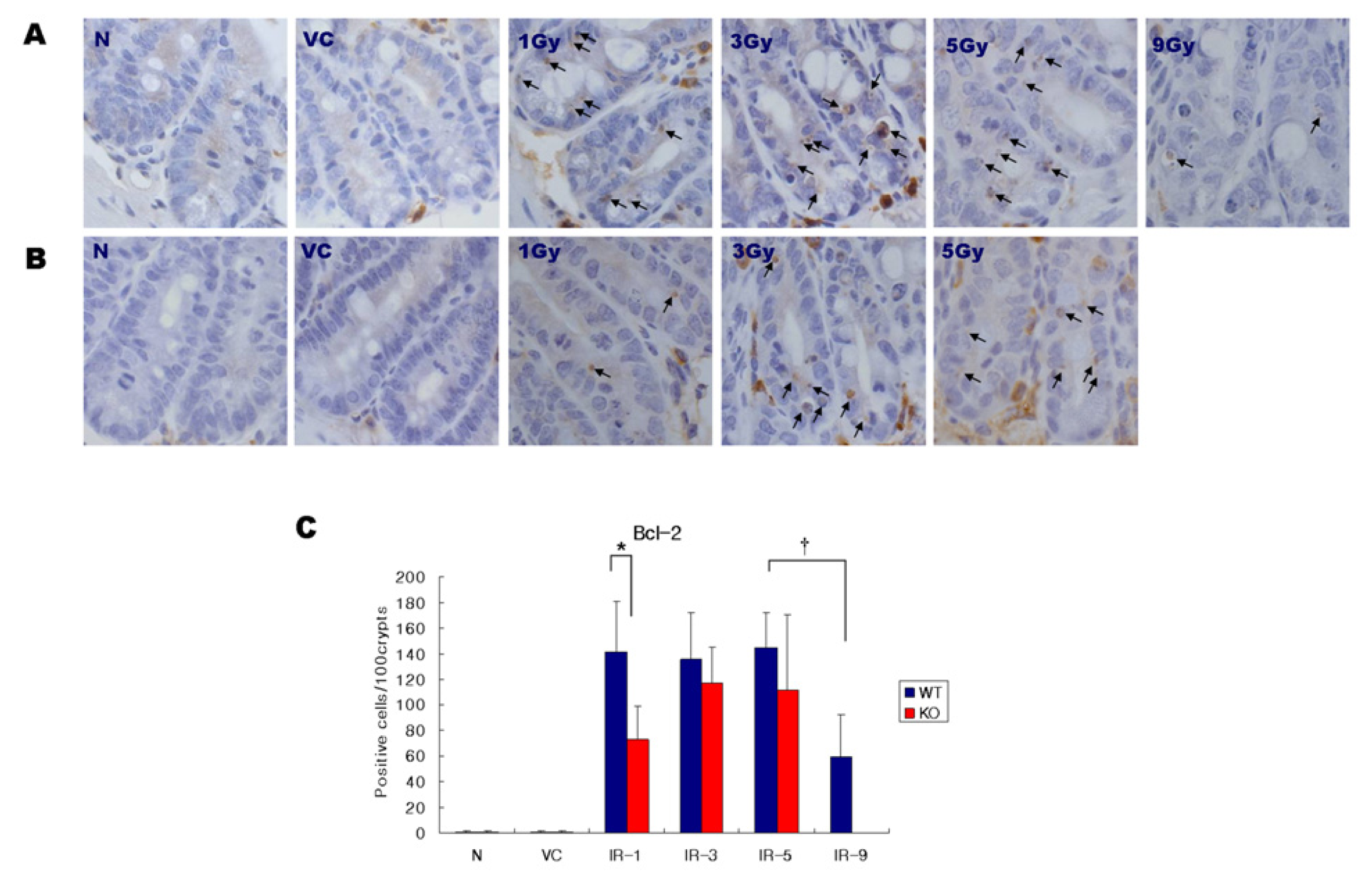
2.3. Expression of Bcl-2
2.4. The Ratio of BAX/Bcl-2
3. Discussion
4. Experimental Section
4.1. Animal Models and Experimental Design
4.2. Irradiation
4.3. Histology and Assessing Apoptosis
4.4. Immunohistochemistry
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Salvesen, G.S.; Dixit, V.M. Caspases: Intracellular signaling by proteolysis. Cell 1997, 91, 443–446. [Google Scholar]
- Merritt, A.J.; Potten, C.S.; Watson, A.J.; Loh, D.Y.; Nakayama, K.; Nakayama, K.; Hickman, J.A. Differential expression of bcl-2 in intestinal epithelia. Correlation with attenuation of apoptosis in colonic crypts and the incidence of colonic neoplasia. J. Cell Sci 1995, 108, 2261–2271. [Google Scholar]
- Maharwal, J.; Samarth, R.M.; Saini, R. Antioxidative effect of rajgira leaf extract in liver of swiss albino mice after exposure to different doses of gamma radiation. Phytother. Res 2005, 19, 717–720. [Google Scholar]
- Mittal, A.; Pathania, V.; Agrawala, P.K.; Prasad, J.; Singh, S.; Goel, H.C. Influence of podophyllum hexandrum on endogenous antioxidant defense system in mice: Possible role in radioprotection. J. Ethnopharmacol 2001, 76, 253–262. [Google Scholar]
- Kondo, Y.; Ishigami, A.; Kubo, S.; Handa, S.; Gomi, K.; Hirokawa, K.; Kajiyama, N.; Chiba, T.; Shimokado, K.; Maruyama, N. Senescence marker protein-30 is a unique enzyme that hydrolyzes diisopropyl phosphorofluoridate in the liver. FEBS Lett 2004, 570, 57–62. [Google Scholar]
- Linster, C.L.; van Schaftingen, E. Vitamin C. Biosynthesis, recycling and degradation in mammals. FEBS J 2007, 274, 1–22. [Google Scholar]
- Fujita, T.; Uchida, K.; Maruyama, N. Purification of senescence marker protein-30 (SMP30) and its androgen independent decrease with age in the rat liver. Biochim. Biophys. Acta 1998, 1116, 122–128. [Google Scholar]
- Ishigami, A.; Fujita, T.; Handa, S.; Shirasawa, T.; Koseki, H.; Kitamura, T.; Enomoto, N.; Sato, N.; Shimosawa, T.; Maruyama, N. Senescence marker protein-30 knockout mouse liver is highly susceptible to tumor necrosis factor-alpha- and Fas-mediated apoptosis. Am. J. Pathol 2002, 161, 1273–1281. [Google Scholar]
- Matsuyama, S.; Kitamura, T.; Enomoto, N.; Fujita, T.; Ishigami, A.; Handa, S.; Maruyama, N.; Zheng, D.; Ikejima, K.; Takei, Y.; et al. Senescence marker protein-30 regulates Akt activity and contributes to cell survival in Hep G2 cells. Biochem. Biophys. Res. Commun 2004, 321, 386–390. [Google Scholar]
- Koike, K.; Kondo, Y.; Sekiya, M.; Sato, Y.; Tobino, K.; Iwakami, S.I.; Goto, S.; Takahashi, K.; Maruyama, N.; Seyama, K.; et al. Complete lack of vitamin C intake generates pulmonary emphysema in senescence marker protein-30 knockout mice. Am. J. Physiol. Lung. Cell. Mol. Physiol 2010, 298, L784–L792. [Google Scholar]
- Park, J.K.; Ki, M.R.; Lee, H.R.; Hong, I.H.; Ji, A.R.; Ishigami, A.; Park, S.I.; Kim, J.M.; Chung, H.Y.; Yoo, S.E.; et al. Vitamin C deficiency attenuates liver fibrosis by way of up-regulated peroxisome proliferator-activated receptor-gamma expression in senescence marker protein 30 knockout mice. Hepatology 2010, 51, 1766–1777. [Google Scholar]
- Park, J.K.; Lee, E.M.; Kim, A.Y.; Lee, E.J.; Min, C.W.; Kang, K.K.; Lee, M.M.; Jeong, K.S. Vitamin C deficiency accelerates bone loss inducing an increase in PPAR-γ expression in SMP30 knockout mice. Int. J. Exp. Pathol 2012, 93, 332–340. [Google Scholar]
- Kondo, Y.; Inai, Y.; Sato, Y.; Handa, S.; Kubo, S.; Shimokado, K.; Goto, S.; Nishikimi, M.; Maruyama, N.; Ishigami, A. Senescence marker protein 30 functions as gluconolactonase in l-ascorbic acid biosynthesis, and its knockout mice are prone to scurvy. Proc. Natl. Acad. Sci. USA 2006, 103, 5723–5728. [Google Scholar]
- Son, T.G.; Zou, Y.; Jung, K.J.; Yu, B.P.; Ishigami, A.; Maruyama, N.; Lee, J. SMP30 deficiency causes increased oxidative stress in brain. Mech. Ageing. Dev 2006, 127, 451–457. [Google Scholar]
- Arai, K.Y.; Sato, Y.; Kondo, Y.; Kudo, C.; Tsuchiya, H.; Nomura, Y.; Ishigami, A.; Nishiyama, T. Effects of vitamin C deficiency on the skin of the senescence marker protein-30 (SMP30) knockout mouse. Biochem. Biophys. Res. Commun 2009, 385, 478–483. [Google Scholar]
- Matsuu-Matsuyama, M.; Shichijo, K.; Okaichi, K.; Ishii, K.; Wen, C.Y.; Fukuda, E.; Nakayama, T.; Nakashima, M.; Okumura, Y.; Sekine, I. Sucralfate protects intestinal epithelial cells from radiation-induced apoptosis in rats. J. Radiat. Res. (Tokyo) 2006, 47, 1–8. [Google Scholar]
- Bowen, J.M.; Gibson, R.J.; Cummins, A.G.; Keefe, D.M. Intestinal mucositis: The role of the Bcl-2 family, p53 and caspases in chemotherapy-induced damage. Support. Care. Cancer 2006, 14, 713–731. [Google Scholar]
- Potten, C.S. Radiation and Gut; Potten, C.S., Hendry, J.H., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1995; pp. 1–31. [Google Scholar]
- Hua, G.; Thin, T.H.; Feldman, R.; Haimovitz-Friedman, A.; Clevers, H.; Fuks, Z.; Kolesnick, R. Crypt base columnar stem cells in small intestines of mice are radioresistant. Gastroenterology 2012, 143, 1266–1276. [Google Scholar]
- Potten, C.S. The significance of spontaneous and induced apoptosis in the gastrointestinal tract of mice. Cancer. Metastasis. Rev 1992, 11, 179–195. [Google Scholar]
- Eble, M.J.; Lehnert, T.; Treiber, M.; Latz, D.; Herfarth, C.; Wannenmacher, M. Moderate dose intraoperative and external beam radiotherapy for locally recurrent rectal carcinoma. Radiother. Oncol 1998, 49, 169–174. [Google Scholar]
- Coopersmith, C.M.; Gordon, J.I. Gamma-Ray-induced apoptosis in transgenic mice with proliferative abnormalities in their intestinal epithelium: Re-entry of villus enterocytes into the cell cycle does not affect their radioresistance but enhances the radiosensitivity of the crypt by inducing p53. Oncogene 1997, 15, 131–141. [Google Scholar]
- Reed, J.C. Apoptosis and Cancer Chemotherapy; Hickman, J.A., Dive, C., Eds.; Humana Press: Totowa, MJ, USA, 1999; pp. 99–116. [Google Scholar]
- Oltvai, Z.N.; Milliman, C.L.; Korsmeyer, S.J. Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that accelerates programmed cell death. Cell 1993, 74, 609–619. [Google Scholar]
- Brown, M. What causes the radiation gastrointestinal syndrome? Overview. Int. J. Radiat. Oncol. Biol. Phys 2008, 70, 799–800. [Google Scholar]
- Jee, Y.H.; Jeong, W.I.; Kim, T.H.; Hwang, I.S.; Ahn, M.J.; Joo, H.G.; Hong, S.H.; Jeong, K.S. p53 and cell-cycle-regulated protein expression in small intestinal cells after fast-neutron irradiation in mice. Mol. Cell. Biochem 2005, 270, 21–28. [Google Scholar]
- Wijsman, J.H.; Jonker, R.R.; Keijzer, R.; van de Velde, C.J.; Cornelisse, C.J.; van Dierendonck, J.H. A new method to detect apoptosis in paraffin sections: In situ end-labeling of fragmented DNA. J. Histochem. Cytochem 1993, 41, 7–12. [Google Scholar]
- Kerr, J.F.; Winterford, C.M.; Harmon, B.V. Apoptosis. Its significance in cancer and cancer therapy. Cancer 1994, 73, 2013–2026. [Google Scholar]
- Walker, N.I.; Harmon, B.V.; Gobé, G.C.; Kerr, J.F. Patterns of cell death. Methods Achiev. Exp. Pathol 1998, 13, 18–54. [Google Scholar]
- Ijiri, K.; Potten, C.S. Response of intestinal cells of differing topographical and hierarchical status to ten cytotoxic drugs and five sources of radiation. Br. J. Cancer 1983, 47, 175–185. [Google Scholar]
- Hall, P.A.; Coates, P.J.; Ansari, B.; Hopwood, D. Regulation of cell number in the mammalian gastrointestinal tract: The importance of apoptosis. J. Cell Sci 1994, 107, 3569–3577. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Treatment | SMP30-WT | SMP30-KO |
|---|---|---|---|
| N | Vit.C-free diet + tap water | 6 | 5 |
| VC | Vit.C-free diet + Vit.C 2.5mg/ml in drinking water | 6 | 5 |
| IR-1 Gy | Vit.C-free diet + tap water + IR-1 Gy | 7 | 6 |
| IR-3 Gy | Vit.C-free diet + tap water + IR-3 Gy | 7 | 6 |
| IR-5 Gy | Vit.C-free diet + tap water + IR-5 Gy | 7 | 6 |
| IR-9 Gy | Vit.C-free diet + tap water + IR-9 Gy | 7 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Goo, M.-J.; Park, J.-K.; Hong, I.-H.; Kim, A.-Y.; Lee, E.-M.; Lee, E.-J.; Hwang, M.; Jeong, K.-S. Increased Susceptibility of Radiation-Induced Intestinal Apoptosis in SMP30 KO Mice. Int. J. Mol. Sci. 2013, 14, 11084-11095. https://doi.org/10.3390/ijms140611084
Goo M-J, Park J-K, Hong I-H, Kim A-Y, Lee E-M, Lee E-J, Hwang M, Jeong K-S. Increased Susceptibility of Radiation-Induced Intestinal Apoptosis in SMP30 KO Mice. International Journal of Molecular Sciences. 2013; 14(6):11084-11095. https://doi.org/10.3390/ijms140611084
Chicago/Turabian StyleGoo, Moon-Jung, Jin-Kyu Park, Il-Hwa Hong, Ah-Young Kim, Eun-Mi Lee, Eun-Joo Lee, Meeyul Hwang, and Kyu-Shik Jeong. 2013. "Increased Susceptibility of Radiation-Induced Intestinal Apoptosis in SMP30 KO Mice" International Journal of Molecular Sciences 14, no. 6: 11084-11095. https://doi.org/10.3390/ijms140611084