Cisplatin Protects against Acute Liver Failure by Inhibiting Nuclear HMGB1 Release

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
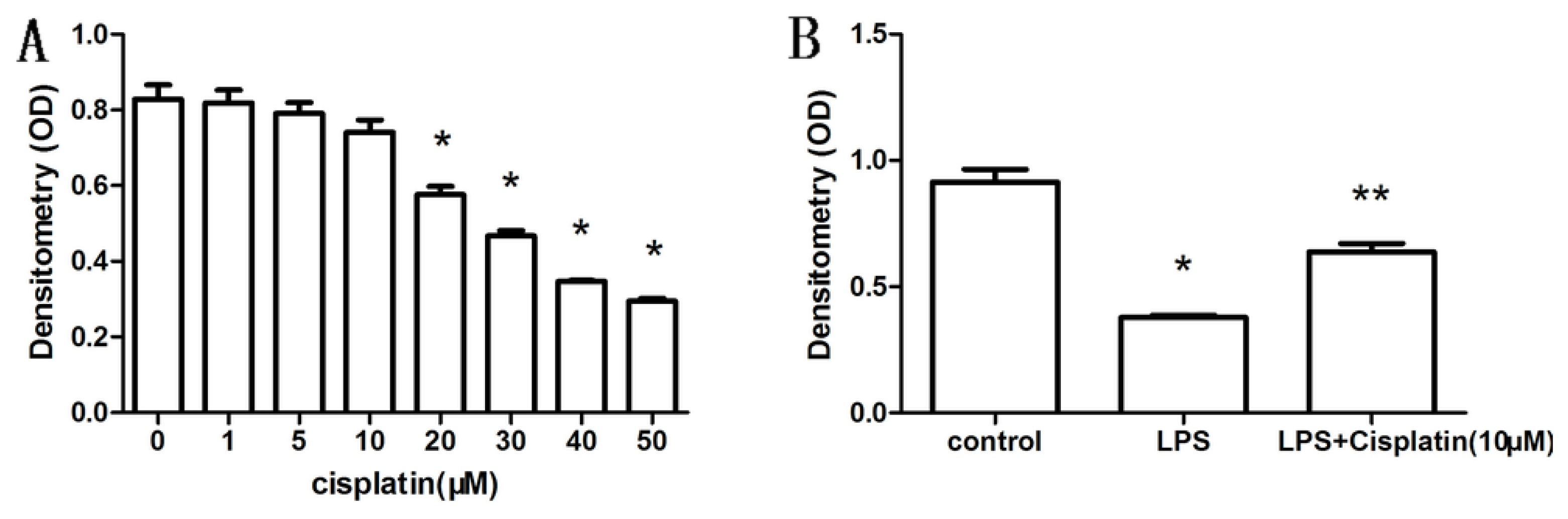
2.1. Effect of Cisplatin on the Viability of Macrophage-Like Cells
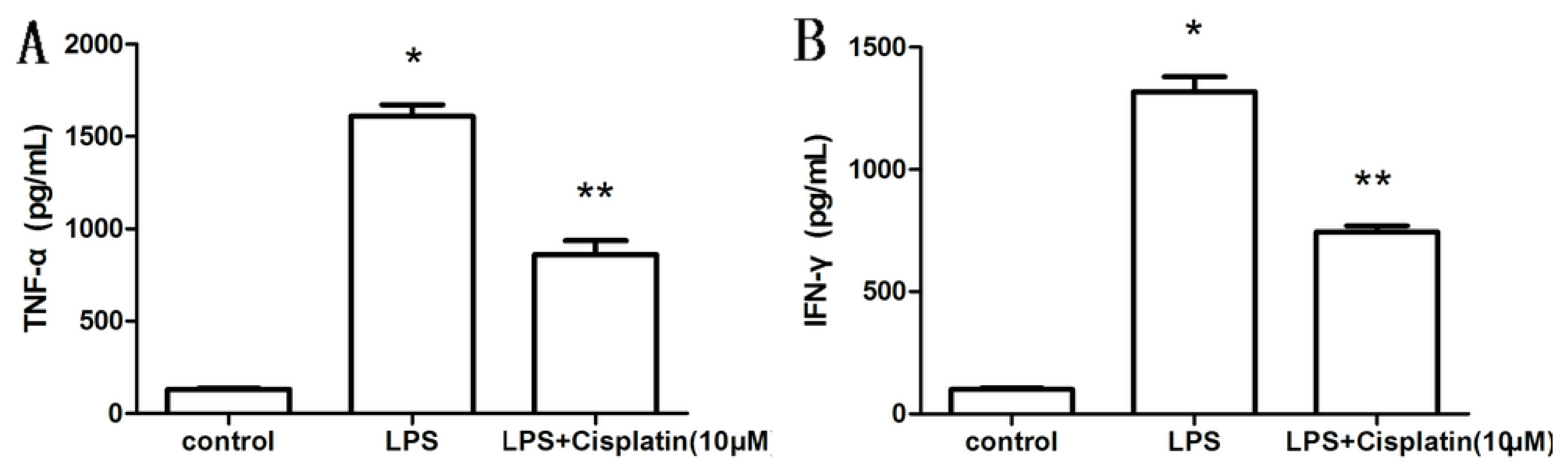
2.2. Effect of Cisplatin on Cytokine Production in Macrophage-Like Cells
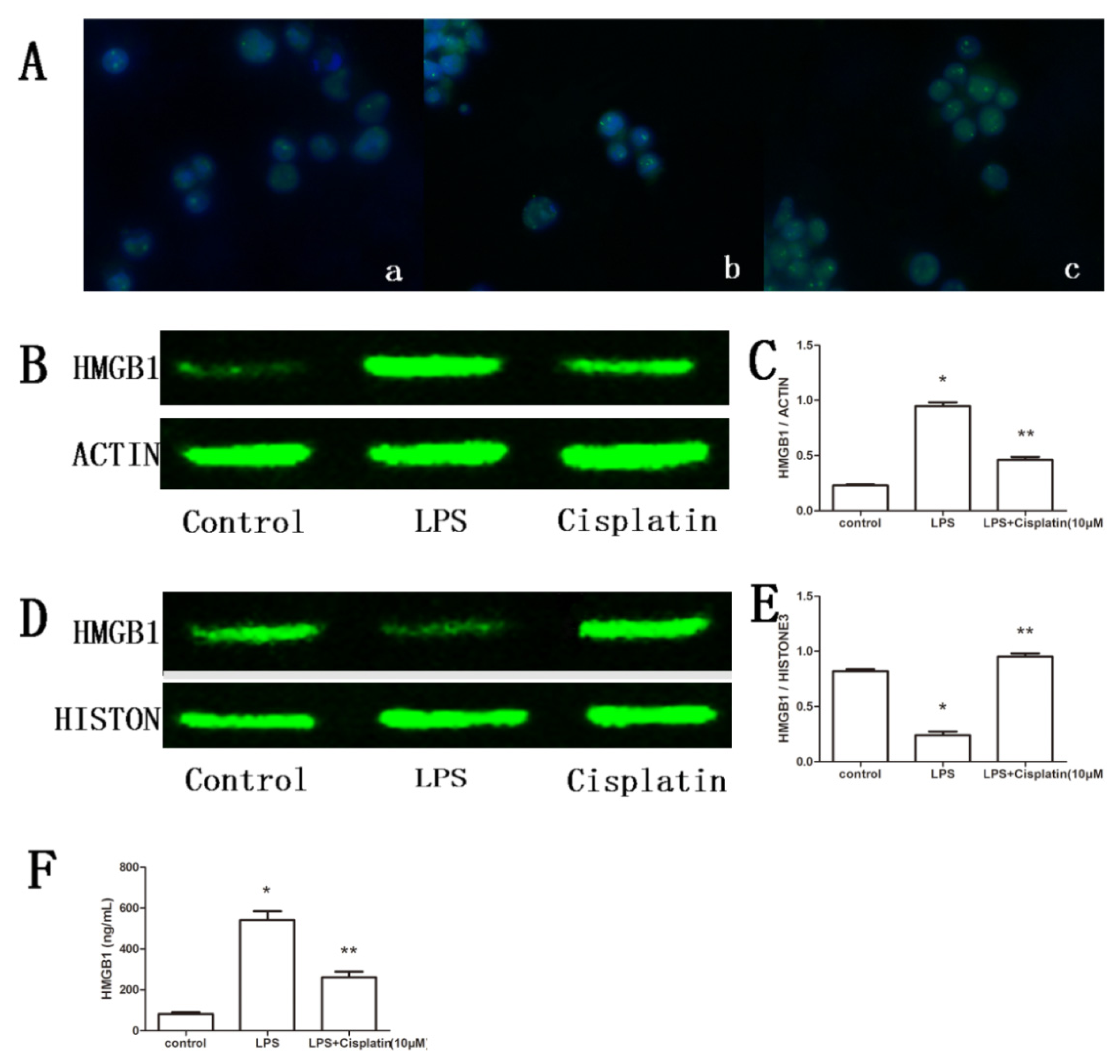
2.3. Effect of Cisplatin on HMGB1 in Macrophage-Like Cells
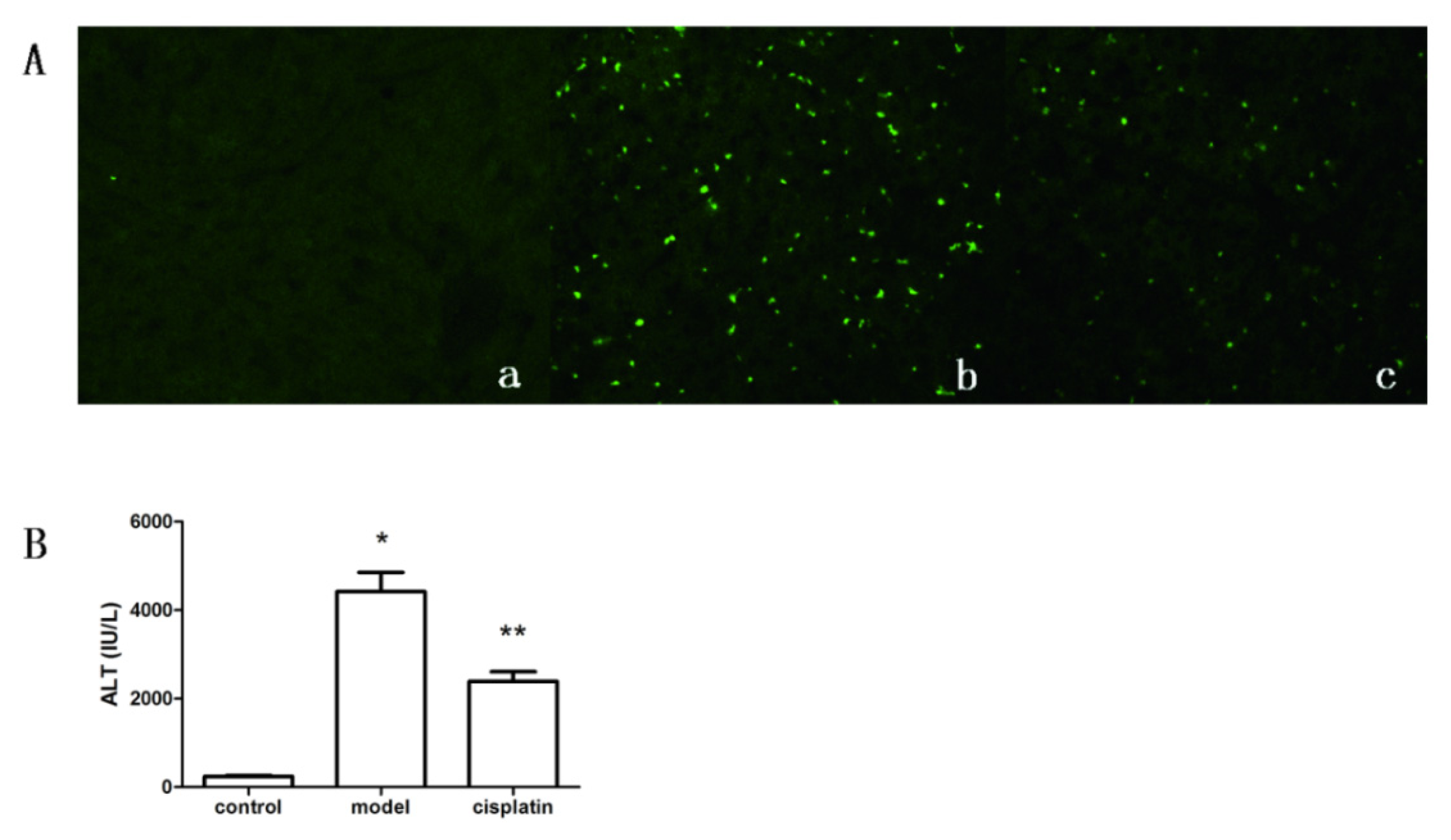
2.4. Effect of Cisplatin on ALT Level and Apoptosis of Hepatic Tissue
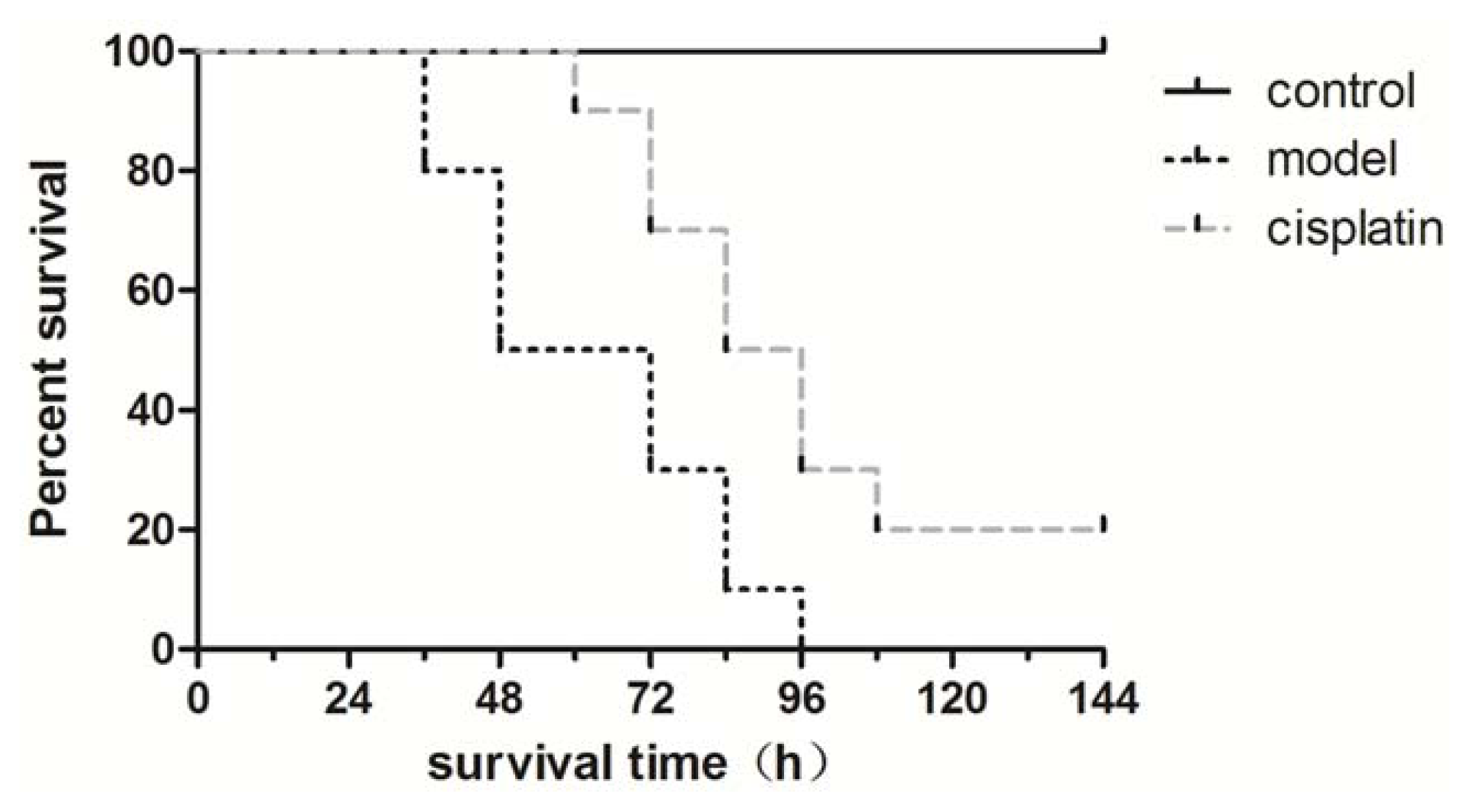
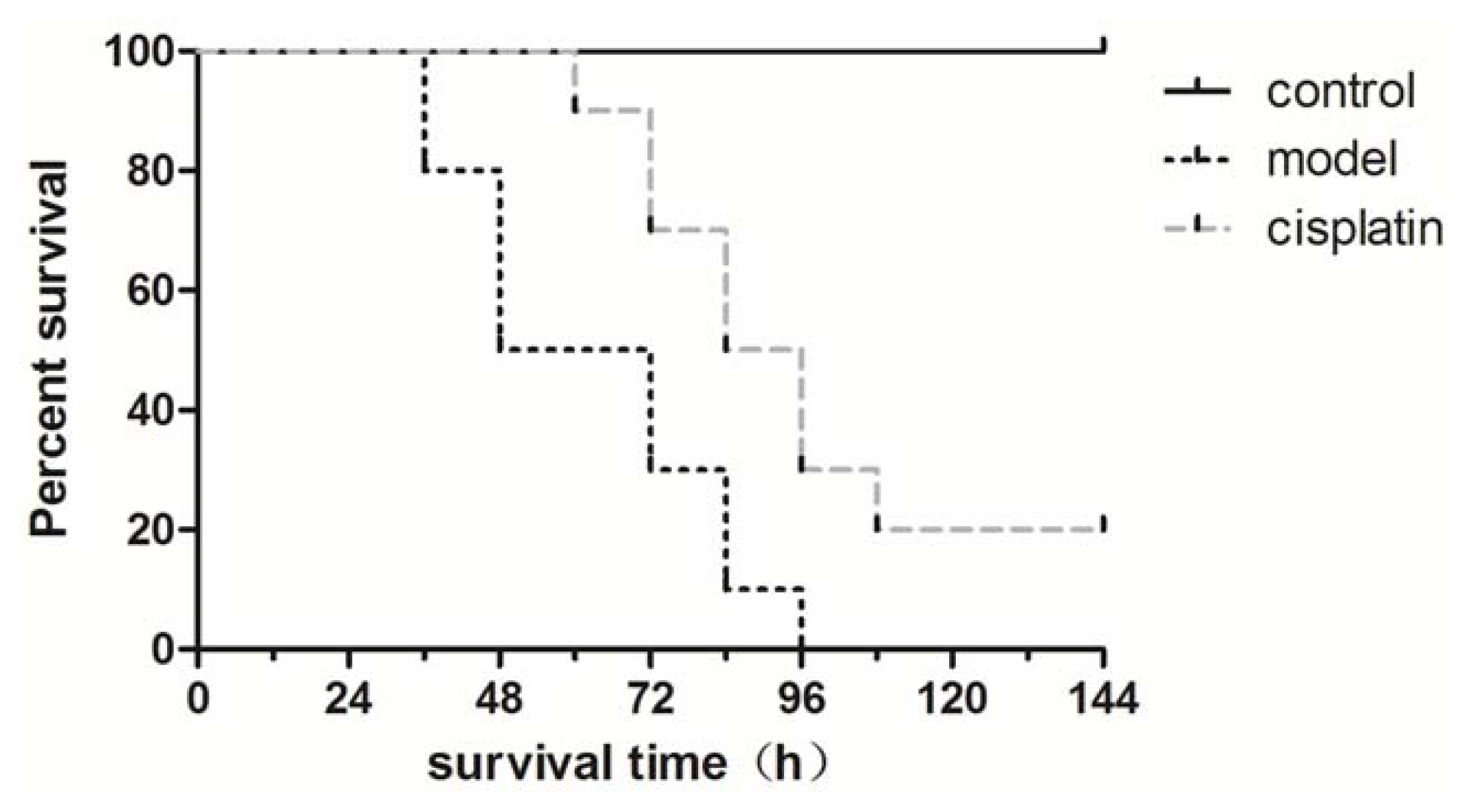
2.5. Effect of Cisplatin on Survival Time
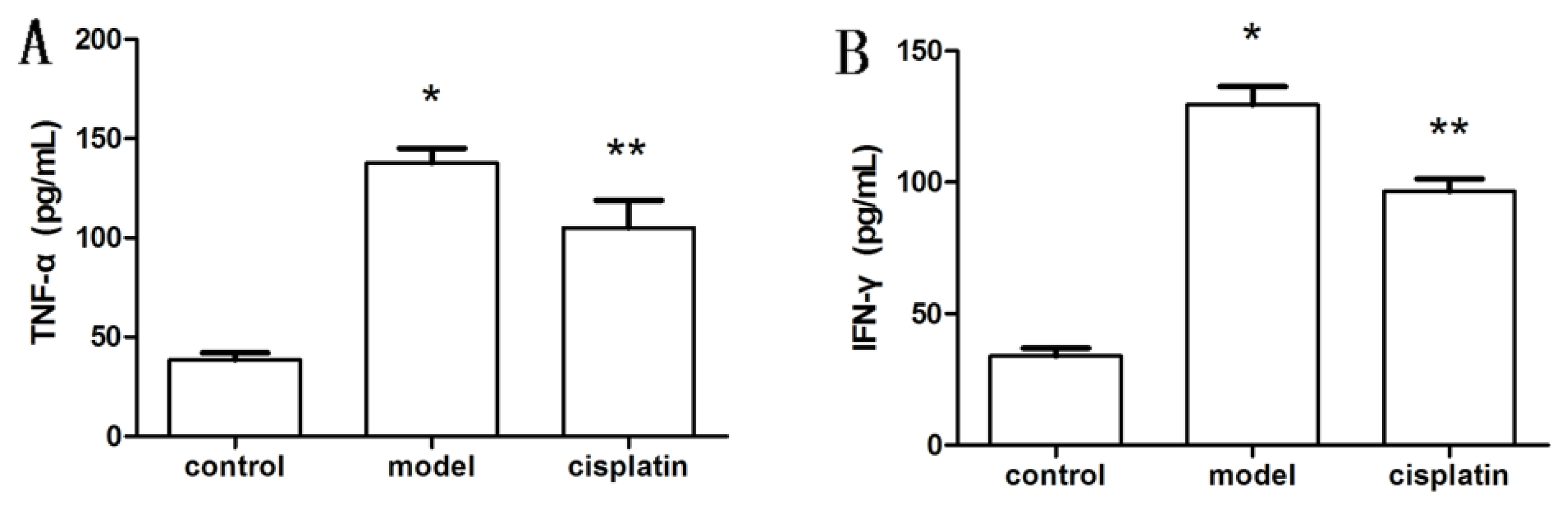
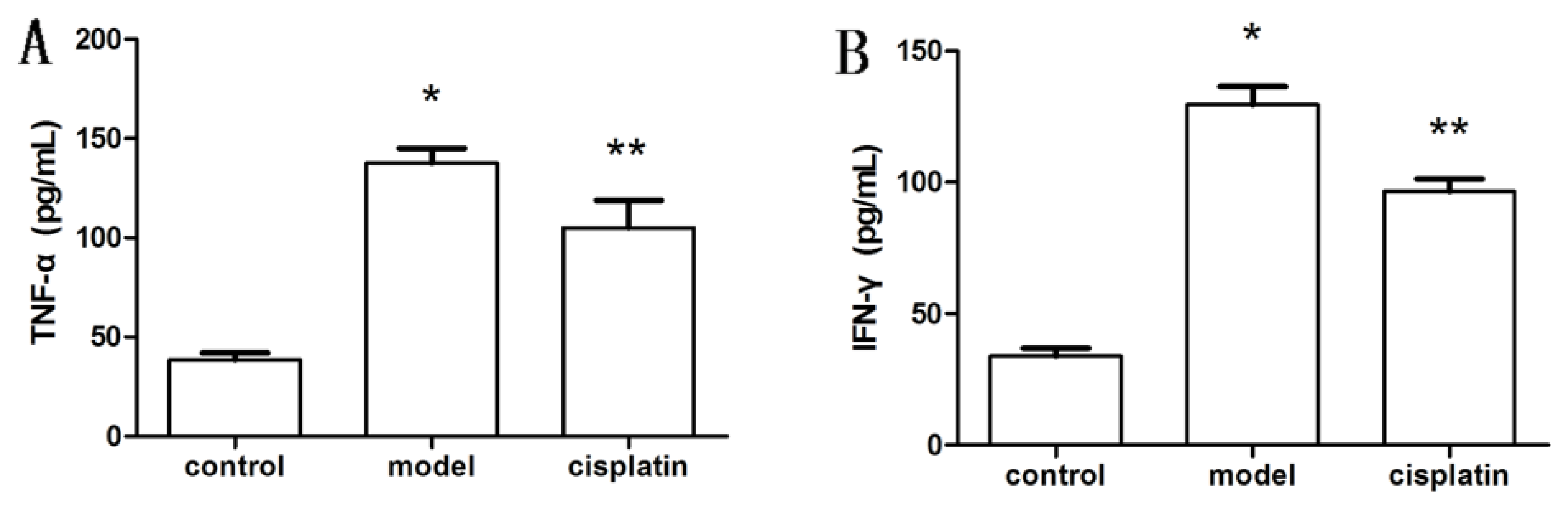
2.6. Effect of Cisplatin on Cytokine Production in ALF
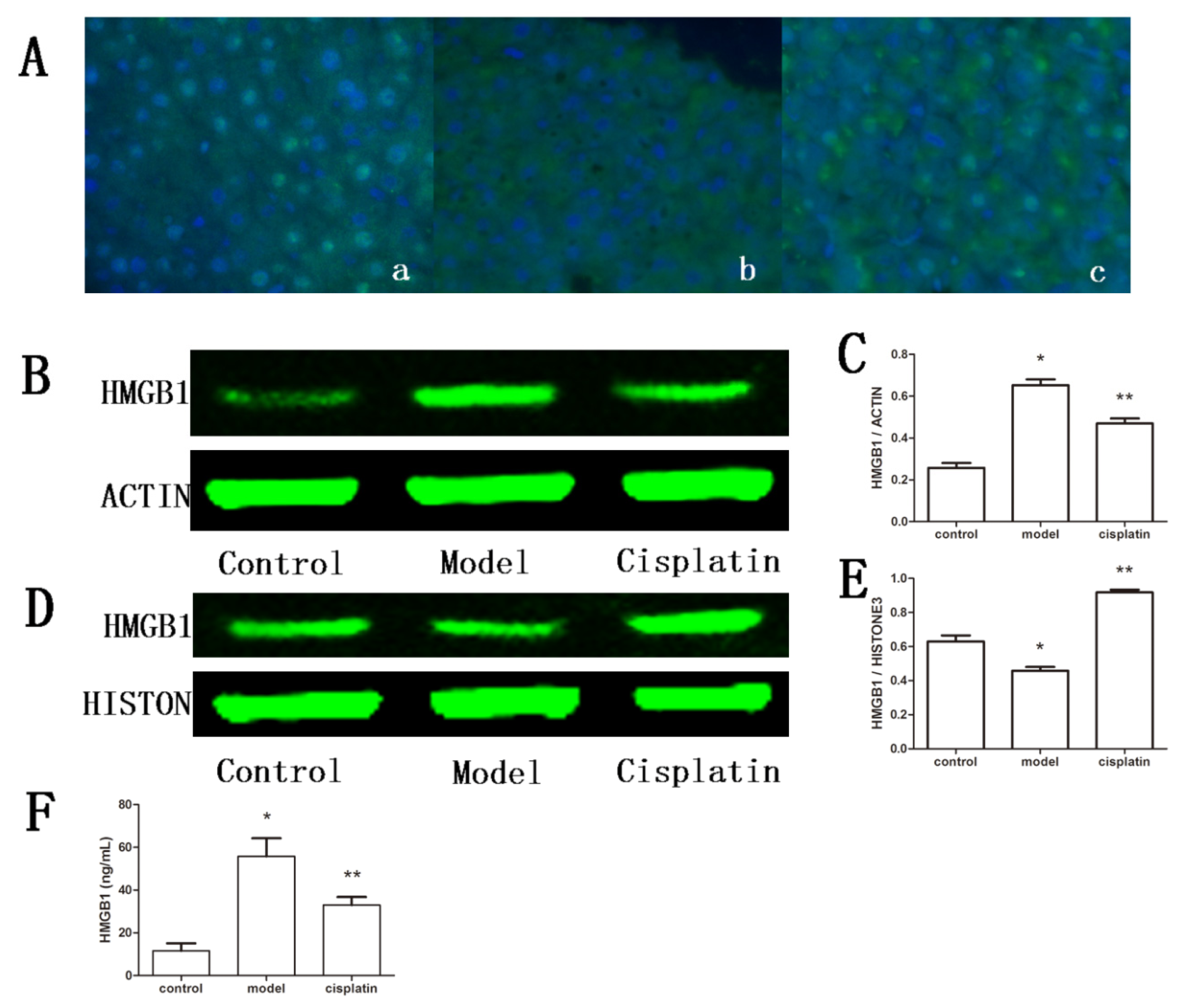
2.7. Effect of Cisplatin on HMGB1 in ALF
3. Experimental Section
3.1. Cell Culture and Treatment
3.2. Animals
3.3. Model Production and Sample Collection
3.4. Cell Viability Test
3.5. TUNEL Assay
3.6. Analysis of Cell Supernatants and Blood Samples
3.7. Extraction of Nuclear and Cytoplasmic Protein from Cells and Hepatic Tissue
3.8. Western Blot Analysis of HMGB1 Levels from U937 Cells and Hepatic Tissue
3.9. Immunohistological Analysis of HMGB1
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Lotze, M.T.; Tracey, K.J. High-mobility group box 1 protein (HMGB1): Nuclear weapon in the immune arsenal. Nat. Rev. Immunol 2005, 5, 331–342. [Google Scholar]
- Zhang, C.C.; Krieg, S.; Shapiro, D.J. HMG-1 stimulates estrogen response element binding by estrogen receptor from stably transfected HeLa cells. Mol. Endocrinol 1999, 13, 632–643. [Google Scholar]
- Bustin, M. Regulation of DNA-dependent activities by the functional motifs of the high-mobility-group chromosomal proteins. Mol. Cell. Biol 1999, 19, 5237–5246. [Google Scholar]
- Yasuda, T.; Ueda, T.; Shinzeki, M.; Sawa, H.; Nakajima, T.; Takeyama, Y.; Kuroda, Y. Increase of high-mobility group box chromosomal protein 1 in blood and injured organs in experimental severe acute pancreatitis. Pancreas 2007, 34, 487–488. [Google Scholar]
- Ueno, H.; Matsuda, T.; Hashimoto, S.; Amaya, F.; Kitamura, Y.; Tanaka, M.; Kobayashi, A.; Maruyama, I.; Yamada, S.; Hasegawa, N.; et al. Contributions of high mobility group box protein in experimental and clinical acute lung injury. Am. J. Respir. Crit. Care Med 2004, 170, 1310–1316. [Google Scholar]
- Hamada, T.; Torikai, M.; Kuwazuru, A.; Tanaka, M.; Horai, N.; Fukuda, T.; Yamada, S.; Nagayama, S.; Hashiguchi, K.; Sunahara, N.; et al. Extracellular high mobility group box chromosomal protein 1 is a coupling factor for hypoxia and inflammation in arthritis. Arthritis Rheum 2008, 58, 2675–2685. [Google Scholar]
- Fan, J.; Li, Y.; Levy, R.M.; Hackam, D.J.; Vodovotz, Y.; Yang, H.; Tracey, K.J.; Billiar, T.R.; Wilson, M.A. Hemorrhagic shock induces NAD(P)H oxidase activation in neutrophils: Role of HMGB1-TLR4 signaling. J. Immunol 2007, 178, 6573–6580. [Google Scholar]
- Tsung, A.; Sahai, R.; Tanaka, H.; Nakao, A.; Fink, M.P.; Lotze, M.T.; Yang, H.; Li, J.; Tracey, K.J.; Geller, D.A.; et al. The nuclear factor HMGB1 mediates hepatic injury after murine liver ischemia-reperfusion. J. Exp. Med 2005, 201, 1135–1143. [Google Scholar]
- Rabik, C.A.; Dolan, M.E. Molecular mechanisms of resistance and toxicity associated with platinating agents. Cancer Treat Rev 2007, 33, 9–23. [Google Scholar]
- Jamieson, E.R.; Lippard, S.J. Structure, recognition, and processing of cisplatin-DNA adducts. Chem. Rev 1999, 99, 2467–2498. [Google Scholar]
- WoŸniak, K.; Błasiak, J. Recognition and repair of DNA-cisplatin adducts. Acta Biochim. Pol 2002, 49, 583–596. [Google Scholar]
- Chauhan, P.; Sodhi, A.; Shrivastava, A. Cisplatin primes murine peritoneal macrophages for enhanced expression of nitric oxide, proinflammatory cytokines, TLRs, transcription factors and activation of MAP kinases upon co-incubation with L929 cells. Immunobiology 2009, 214, 197–209. [Google Scholar]
- Pan, P.; Cardinal, J.; Dhupar, R.; Rosengart, M.R.; Lotze, M.T.; Geller, D.A.; Billiar, T.R.; Tsung, A. Low-dose cisplatin administration in murine cecal ligation and puncture prevents the systemic release of HMGB1 and attenuates lethality. J. Leukoc. Biol 2009, 86, 625–632. [Google Scholar]
- Park, S.; Lippard, S.J. Redox state-dependent interaction of HMGB1 and cisplatin-modified DNA. Biochemistry 2011, 50, 2567–2574. [Google Scholar]
- Cardinal, J.; Pan, P.; Dhupar, R.; Ross, M.; Nakao, A.; Lotze, M.; Billiar, T.; Geller, D.; Tsung, A. Cisplatin prevents high mobility group box 1 release and is protective in a murine model of hepatic ischemia/reperfusion injury. Hepatology 2009, 50, 565–574. [Google Scholar]
- Sozinov, A.S. Systemic endotoxemia during chronic viral hepatitis. Bull. Exp. Biol. Med 2002, 133, 153–155. [Google Scholar]
- Beutler, B.; Rietschel, E.T. Innate immune sensing and its roots: The story of endotoxin. Nat. Rev. Immunol 2003, 3, 169–176. [Google Scholar]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar]
- Del Bufalo, A.; Bernad, J.; Dardenne, C.; Verda, D.; Meunier, J.R.; Rousset, F.; Martinozzi-Teissier, S.; Pipy, B. Contact sensitizers modulate the arachidonic acid metabolism of PMA-differentiated U-937 monocytic cells activated by LPS. Toxicol. Appl. Pharmacol 2011, 256, 35–43. [Google Scholar]
- Siddik, Z.H. Cisplatin: Mode of cytotoxic action and molecular basis of resistance. Oncogene 2003, 22, 7265–7279. [Google Scholar]
- Yan, S.; Ding, Y.; Sun, F.; Lu, Z.; Xue, L.; Liu, X.; Shuai, M.; Fang, C.; Wang, Y.; Cheng, H.; et al. Pretreatment of cisplatin in recipients attenuates post-transplantation pancreatitis in murine model. Int. J. Biol. Sci 2012, 8, 298–309. [Google Scholar]
- Yang, H.; Ochani, M.; Li, J.; Qiang, X.; Tanovic, M.; Harris, H.E.; Susarla, S.M.; Ulloa, L.; Wang, H.; DiRaimo, R.; et al. Reversing established sepsis with antagonists of endogenous high-mobility group box 1. Proc. Natl. Acad. Sci. USA 2004, 101, 296–301. [Google Scholar]
- Wang, H.; Bloom, O.; Zhang, M.; Vishnubhakat, J.M.; Ombrellino, M.; Che, J.; Frazier, A.; Yang, H.; Ivanova, S.; Borovikova, L.; et al. HMG-1 as a late mediator of endotoxin lethality in mice. Science 1999, 285, 248–251. [Google Scholar]
- Rendon-Mitchell, B.; Ochani, M.; Li, J.; Han, J.; Wang, H.; Yang, H.; Susarla, S.; Czura, C.; Mitchell, R.A.; Chen, G.; et al. IFN-gamma induces high mobility group box 1 protein release partly through a TNF-dependent mechanism. J. Immunol 2003, 170, 3890–3897. [Google Scholar]
- Gardella, S.; Andrei, C.; Ferrera, D.; Lotti, L.V.; Torrisi, M.R.; Bianchi, M.E.; Rubartelli, A. The nuclear protein HMGB1 is secreted by monocytes via a non-classical, vesicle-mediated secretory pathway. EMBO Rep 2002, 3, 995–1001. [Google Scholar]
- Bonaldi, T.; Talamo, F.; Scaffidi, P.; Ferrera, D.; Porto, A.; Bachi, A.; Rubartelli, A.; Agresti, A.; Bianchi, M.E. Monocytic cells hyperacetylate chromatin protein HMGB1 to redirect it towards secretion. EMBO J 2003, 22, 5551–5560. [Google Scholar]
- Liu, S.; Stolz, D.B.; Sappington, P.L.; Macias, C.A.; Killeen, M.E.; Tenhunen, J.J.; Delude, R.L.; Fink, M.P. HMGB1 is secreted by immunostimulated enterocytes and contributes to cytomix-induced hyperpermeability of Caco-2 monolayers. Am. J. Physiol. Cell Physiol 2006, 290, C990–C999. [Google Scholar]
- Scaffidi, P.; Misteli, T.; Bianchi, M.E. Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 2002, 418, 191–195. [Google Scholar]
- Yang, H.; Wang, H.; Czura, C.J.; Tracey, K.J. The cytokine activity of HMGB1. J. Leukoc. Biol 2005, 78, 1–8. [Google Scholar]
- Wang, H.; Yang, H.; Czura, C.J.; Sama, A.E.; Tracey, K.J. HMGB1 as a late mediator of lethal systemic inflammation. Am. J. Respir. Crit. Care Med 2001, 164, 1768–1773. [Google Scholar]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar]
- Yang, H.; Tracey, K.J. Targeting HMGB1 in inflammation. Biochim. Biophys. Acta 2010, 1799, 149–156. [Google Scholar]
- Zhang, C.X.; Chang, P.V.; Lippard, S.J. Identification of nuclear proteins that interact with platinum-modified DNA by photoaffinity labeling. J. Am. Chem. Soc 2004, 126, 6536–6537. [Google Scholar]

© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, X.; Wang, L.-K.; Wang, L.-W.; Han, X.-Q.; Yang, F.; Gong, Z.-J. Cisplatin Protects against Acute Liver Failure by Inhibiting Nuclear HMGB1 Release. Int. J. Mol. Sci. 2013, 14, 11224-11237. https://doi.org/10.3390/ijms140611224
Li X, Wang L-K, Wang L-W, Han X-Q, Yang F, Gong Z-J. Cisplatin Protects against Acute Liver Failure by Inhibiting Nuclear HMGB1 Release. International Journal of Molecular Sciences. 2013; 14(6):11224-11237. https://doi.org/10.3390/ijms140611224
Chicago/Turabian StyleLi, Xun, Li-Kun Wang, Lu-Wen Wang, Xiao-Qun Han, Fan Yang, and Zuo-Jiong Gong. 2013. "Cisplatin Protects against Acute Liver Failure by Inhibiting Nuclear HMGB1 Release" International Journal of Molecular Sciences 14, no. 6: 11224-11237. https://doi.org/10.3390/ijms140611224