Characterization of the Pinus massoniana Transcriptional Response to Bursaphelenchus xylophilus Infection Using Suppression Subtractive Hybridization

Abstract
:
1. Introduction
2. Results
2.1. Analysis of cDNA Libraries Constructed by SSH
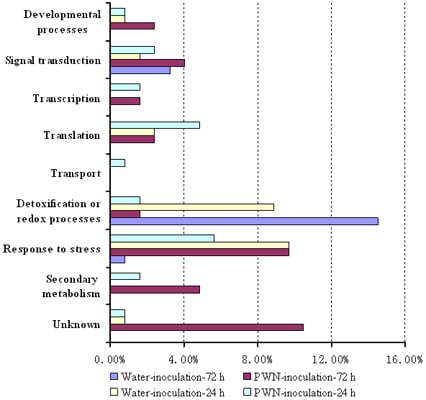
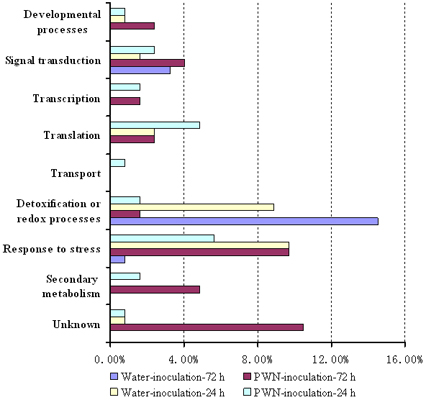
2.2. Functional Classification and Identification of differentially Expressed Genes in PWN-Inoculated P. massoniana Trees
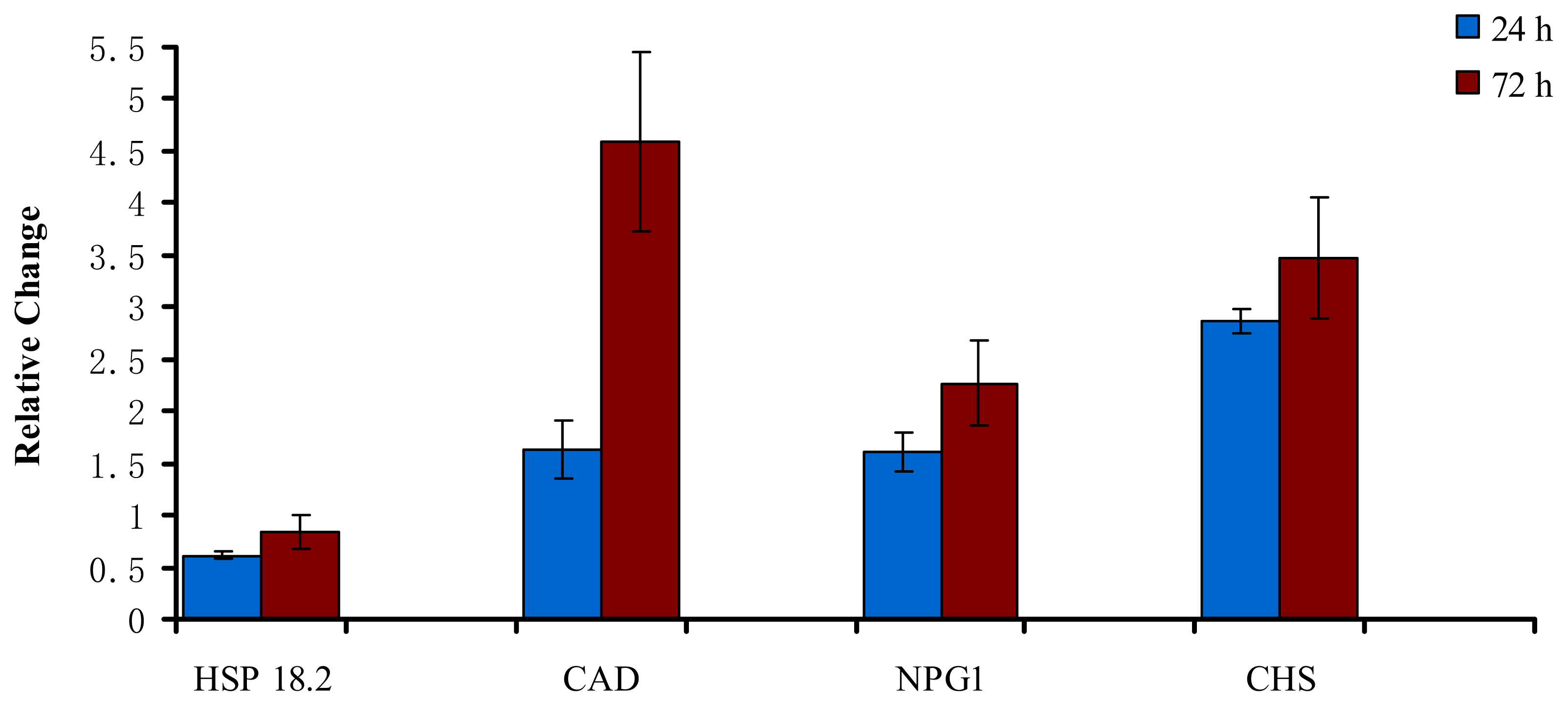
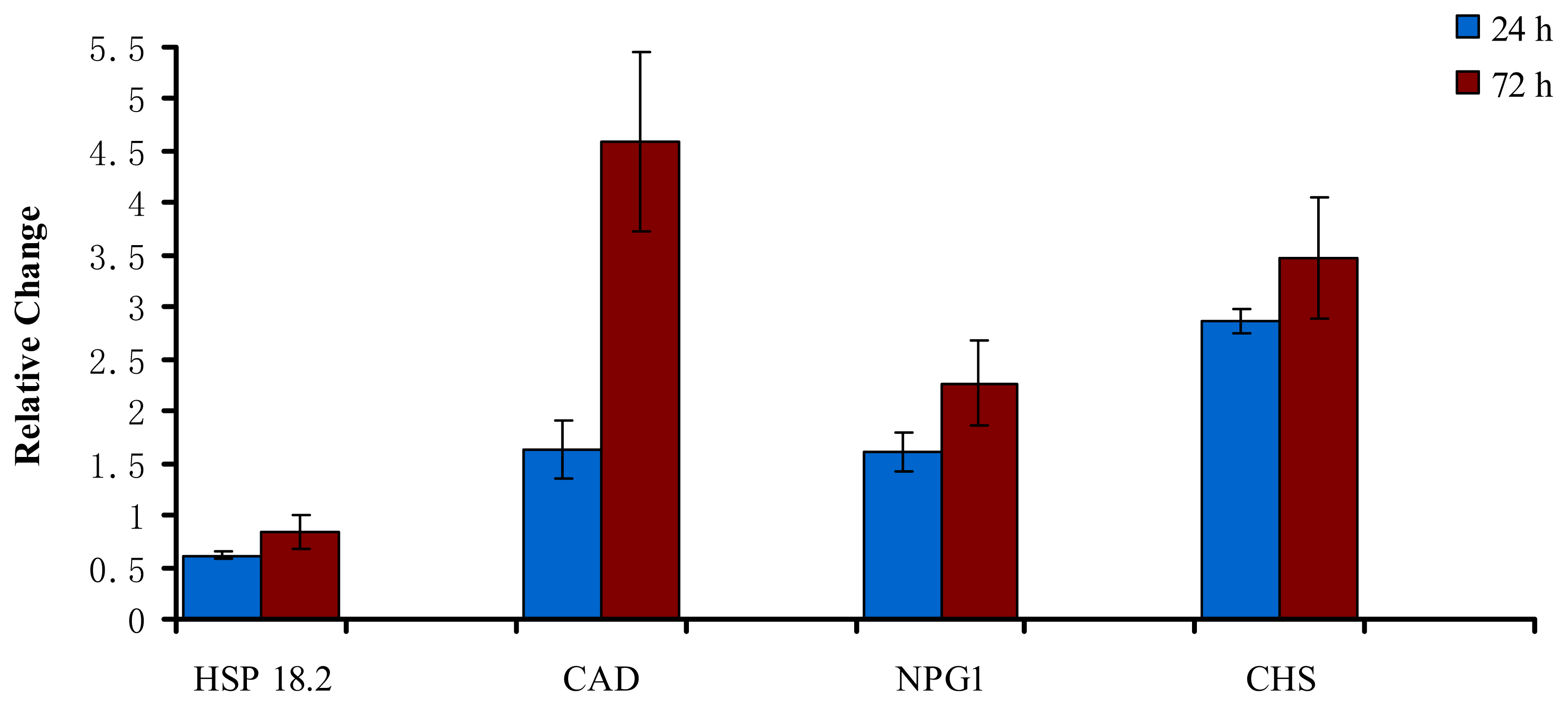
2.3. Validation of Differential Expression using Selected SSH Clones and qRT-PCR
3. Discussion
4. Experimental Section
4.1. Plant Material and Nematode Culture
4.2. PWN Inoculation and Sampling Time
4.3. RNA Extraction and cDNA Synthesis
4.4. Construction of SSH Libraries and Dot-Blot Hybridization
4.5. DNA Sequencing and Data Analysis
4.6. Quantitative Real-Time PCR (qRT-PCR)
5. Conclusions
Supplementary Information
ijms-14-11356-s001.pdfAcknowledgments
Conflict of Interest
References
- Tokushige, Y.; Kiyohara, T. Bursaphelenchus sp. in the wood of dead pine trees. J. Jpn. For. Soc 1969, 51, 193–195. [Google Scholar]
- Huang, L.; Ye, J.; Wu, X.; Xu, X.; Sheng, J.; Zhou, Q. Detection of pine wood nematode using a real-time PCR assay to target the DNA topoisomerase I gene. Eur. J. Plant Pathol 2010, 127, 89–98. [Google Scholar]
- Vicente, C.; Espada, M.; Vieira, P.; Mota, M. Pine Wilt Disease: A threat to European forestry. Eur. J. Plant Pathol 2012, 133, 89–99. [Google Scholar]
- Forestry Agency, Annual Report on Trends of Forest and Forestry-Fiscal Year 2003; The Ministry of Agriculture, Forestry and Fisheries of Japan: Tokyo, Japan, 2004.
- Shin, H.; Lee, H.; Woo, K.S.; Noh, E.W.; Koo, Y.B.; Lee, K.J. Identification of genes upregulated by pinewood nematode inoculation in Japanese red pine. Tree Physiol 2009, 29, 411–421. [Google Scholar]
- Yang, B.J.; Pan, H.Y.; Tang, J.; Wang, Y.Y.; Wang, L.F.; Wang, Q. Pine Wood Nematode Disease; Chinese Forestry Press: Beijing, China, 2003; pp. 6–143. [Google Scholar]
- Jones, J.T.; Moens, M.; Mota, M.; Li, H.; Kikuchi, T. Bursaphelenchus xylophilus: Opportunities in comparative genomics and molecular host-parasite interactions. Mol. Plant Pathol 2008, 9, 357–368. [Google Scholar]
- Fukuda, K. Physiological process of the symptom development and resistance mechanism in pine wilt disease. J. For. Res 1997, 2, 171–181. [Google Scholar]
- Myers, R.F. Pathogenesis in pine wilt caused by pinewood nematode, Bursaphelenchus xylophilus. J. Nematol 1988, 20, 236–244. [Google Scholar]
- Fukuda, K.; Utsuzawa, S.; Sakaue, D. Correlation between acoustic emission, water status and xylem embolism in pine wilt disease. Tree Physiol 2007, 27, 969–976. [Google Scholar]
- Schiffer, R.; Görg, R.; Jarosch, B.; Beckhove, U.; Bahrenberg, G.; Kogel, K.H.; Schulze-Lefert, P. Tissue dependence and differential cordycepin sensitivity of race-specific resistance responses in the barley-powdery mildew interaction. Mol. Plant Microbe Interact 1997, 10, 830–839. [Google Scholar]
- Santos, C.S.S.; Vasconcelos, M.W. Identification of genes differentially expressed in Pinus pinaster and Pinus pinea after infection with the pine wood nematode. Eur. J. Plant Pathol 2012, 132, 407–418. [Google Scholar]
- Hirao, T.; Fukatsu, E.; Watanabe, A. Characterization of resistance to pine wood nematode infection in Pinus thunbergii using suppression subtractive hybridization. BMC Plant Biol 2012, 12, 13. [Google Scholar]
- Matsye, P.D.; Kumar, R.; Hosseini, P.; Jones, C.M.; Tremblay, A.; Alkharouf, N.W.; Matthews, B.F.; Klink, V.P. Mapping cell fate decisions that occur during soybean defense responses. Plant Mol. Biol 2011, 77, 513–528. [Google Scholar]
- Matsye, P.D.; Lawrence, G.W.; Youssef, R.M.; Kim, K.H.; Lawrence, K.S.; Matthews, B.F.; Klink, V.P. The expression of a naturally occurring, truncated allele of an α-SNAP gene suppresses plant parasitic nematode infection. Plant Mol. Biol 2012, 80, 131–155. [Google Scholar]
- Scherer, N.M.; Thompson, C.E.; Freitas, L.B.; Bonatto, S.L.; Salzano, F.M. Patterns of molecular evolution in pathogenesis-related proteins. Genet. Mol. Biol 2005, 28, 645–653. [Google Scholar]
- Nose, M.; Shiraishi, S. Comparison of the gene expression profiles of resistant and non-resistant Japanese black pine inoculated with pine wood nematode using a modified LongSAGE technique. For. Path 2011, 41, 143–155. [Google Scholar]
- Wingfield, M.J. Fungi associated with the pine wood nematode, Bursaphelenchus xylophilus, and cerambycid beetles in Wisconsin. Mycologia 1987, 79, 325–328. [Google Scholar]
- Osmond, R.I.W.; Hrmova, M.; Fontaine, F.; Imberty, A.; Fincher, G.B. Binding interactions between barley thaumatin-like proteins and (1,3)-β-d-glucans. Eur. J. Biochem 2001, 268, 4190–4199. [Google Scholar]
- Zamani, A.; Sturrock, R.; Ekramoddoullah, A.K.M.; Wiseman, S.B.; Griffith, M. Endochitinase activity in the apoplastic fluid of Phellinus weirii-infected Douglas-fir and its association with over wintering and antifreeze activity. For. Pathol 2003, 33, 299–316. [Google Scholar]
- Regente, M.C.; Giudici, A.M.; Villalaín, J.; de la Canal, L. The cytotoxic properties of a plant lipid transfer protein involve membrane permeabilization of target cells. Lett. Appl. Microbiol 2005, 40, 183–189. [Google Scholar]
- Santos, C.S.S.; Pinheiro, M.; Silva, A.I.; Egas, C.; Vasconcelos, M.W. Searching for resistance genes to Bursaphelenchus xylophilus using high throughput screening. BMC Genomics 2012, 13, 599. [Google Scholar]
- Vandenborre, G.; Smagghe, G.; van Damme, E.J.M. Plant lectins as defense proteins against phytophagous insects. Phytochemistry 2011, 72, 1538–1550. [Google Scholar]
- Kanzaki, H.; Saitoh, H.; Ito, A.; Fujisawa, S.; Kamoun, S.; Katou, S.; Yoshioka, H.; Terauchi, R. Cytosolic HSP90 and HSP70 are essential components of INF1-mediated hypersensitive response and non-host resistance to Pseudomonas cichorii in Nicotiana benthamiana. Mol. Plant Pathol 2003, 4, 383–391. [Google Scholar]
- Klink, V.P.; Overall, C.C.; Alkharouf, N.W.; MacDonald, M.H.; Matthews, B.F. Laser capture microdissection (LCM) and comparative microarray expression analysis of syncytial cells isolated from incompatible and compatible soybean (Glycine max) roots infected by the soybean cyst nematode (Heterodera glycines). Planta 2007, 226, 1389–1409. [Google Scholar]
- Zhang, Q.; Bai, G.; Yang, W.; Li, H.; Xiong, H. Pathogenic cellulase assay of pine wilt disease and immunological localization. Biosci. Biotechnol. Biochem 2006, 70, 2727–2732. [Google Scholar]
- Ralph, S.; Park, J.Y.; Bohlmann, J.; Mansfield, S.D. Dirigent proteins in conifer defense: Gene discovery, phylogeny, and differential wound- and insect-induced expression of a family of DIR and DIR-like genes in spruce (Picea spp.). Plant Mol. Biol 2006, 60, 21–40. [Google Scholar]
- Bidlack, W.R.; Tappel, A.L. Fluorescent products of phospholipids during lipid peroxidation. Lipids 1973, 8, 203–207. [Google Scholar]
- Kato, S.; Misawa, T. Lipid peroxidation during the appearance of hypersensitive reaction in cowpea leaves infected with cucumber mosaic virus. Ann. Phytopathol. Soc. Jpn 1976, 42, 472–480. [Google Scholar]
- Dhindsa, R.S.; Plumb-dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot 1981, 32, 93–101. [Google Scholar]
- Futai, K. Pine Wilt Is an Epidemic Disease in Forests: Notes on the Interrelationship of Forest Microbes; Bun-ichi Sogo Shyuppan: Tokyo, Japan, 2003. [Google Scholar]
- Mano, J.; Belles-Boix, E.; Babiychuk, E.; Inzé, D.; Torii, Y.; Hiraoka, E.; Takimoto, K.; Slooten, L.; Asada, K.; Kushnir, S. Protection against photooxidative injury of tobacco leaves by 2-Alkenal reductase. Detoxication of lipid peroxide-derived reactive carbonyls. Plant Physiol 2005, 139, 1773–1783. [Google Scholar]
- Yamauchi, Y.; Hasegawa, A.; Taninaka, A.; Mizutani, M.; Sugimoto, Y. NADPH-dependent reductases involved in the detoxification of reactive carbonyls in plants. J. Biol. Chem 2011, 286, 6999–7009. [Google Scholar]
- Van Breusegem, F.; Vranová, E.; Dat, J.F.; Inzé, D. The role of active oxygen species in plant signal transduction. Plant Sci 2001, 161, 405–414. [Google Scholar]
- Verelst, W.; Asard, H. A phylogenetic study of cytochrome b561 proteins. Genome Biol 2003, 4, R38. [Google Scholar]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Ann. Rev. Plant Physiol. Plant Mol. Biol 1998, 49, 249–279. [Google Scholar]
- Mir, G.; Demenech, J.; Huguet, G.; Guo, W.J.; Goldsbrough, P.; Atrian, S.; Molinas, M. A plant type 2 metallothionein (MT) from cork tissue responds to oxidative stress. J. Exp. Bot 2004, 55, 2483–2493. [Google Scholar]
- Etscheid, M.; Klumper, S.; Riesner, D. Accumulation of a metallothionein-like mRNA in Norway spruce under environmental stress. J. Phytopathol 1999, 147, 207–213. [Google Scholar]
- Ishida, K.; Hogetsu, T.; Fukuda, K.; Suzuki, K. Cortical responses in Japanese black pine to attack by the pine wood nematode. Can. J. Bot 1993, 71, 1399–1405. [Google Scholar]
- Hückelhoven, R. Cell wall-associated mechanisms of disease resistance and susceptibility. Annu. Rev. Phytopathol 2007, 45, 101–127. [Google Scholar]
- Afzal, A.J.; Natarajan, A.; Saini, N.; Iqbal, M.J.; Geisler, M.; El Shemy, H.A.; Mungur, R.; Willmitzer, L.; Lightfoot, D.A. The nematode resistance allele at the rhg1 locus alters the proteome and primary metabolism of soybean roots. Plant Physiol 2009, 151, 1264–1280. [Google Scholar]
- Burlat, V.; Kwon, M.; Davin, L.B.; Lewis, N.G. Dirigent proteins and dirigent sites in lignifying tissues. Phytochmistry 2001, 57, 883–897. [Google Scholar]
- Keen, N.T. The molecular biology of disease resistance. Plant Mol. Biol 1992, 19, 109–122. [Google Scholar]
- Ithal, N.; Recknor, J.; Nettleton, D.; Hearne, L.; Maier, T.; Baum, T.J.; Mitchum, M.G. Parallel genome-wide expression profiling of host and pathogen during soybean cyst nematode infection of soybean. Mol. Plant Microbe Interact 2007, 20, 293–305. [Google Scholar]
- Chen, W.; Provart, N.J.; Glazebrook, J.; Katagiri, F.; Chang, H.S.; Eulgem, T.; Mauch, F.; Luan, S.; Zou, G.; Whitham, S.A.; et al. Expression profile matrix of Arabidopsis transcription factor genes suggests their putative functions in response to environmental stresses. Plant Cell 2002, 14, 559–574. [Google Scholar]
- Ulker, B.; Somssich, I.E. WRKY transcription factors: From DNA binding towards biological function. Curr. Opin. Plant Biol 2004, 7, 491–498. [Google Scholar]
- Ayoub, S.M. Plant Nematology: An Agricultural Training Aid; NemaAid Publication: Sacramento, CA, USA, 1980; p. 195. [Google Scholar]
- Futai, K.; Furuno, T. The variety of resistances among pine species to pine wood nematode, Bursaphelenchus lignicolus. Bull. Kyoto Univ. For 1979, 51, 23–36. [Google Scholar]
- Online VecScreen Tool. Available online: http://www.ncbi.nlm.nih.gov/VecScreen/VecScreen.html (accessed on 28 March 2013).
- Huang, X.; Madan, A. CAP3: A DNA sequence assembly program. Genome Res 1999, 9, 868–877. [Google Scholar]
- Gene Ontology (GO) Annotation Analysis. Available online: http://www.arabidopsis.org (accessed on 1 April 2013).
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 2001, 29, 2002–2007. [Google Scholar]
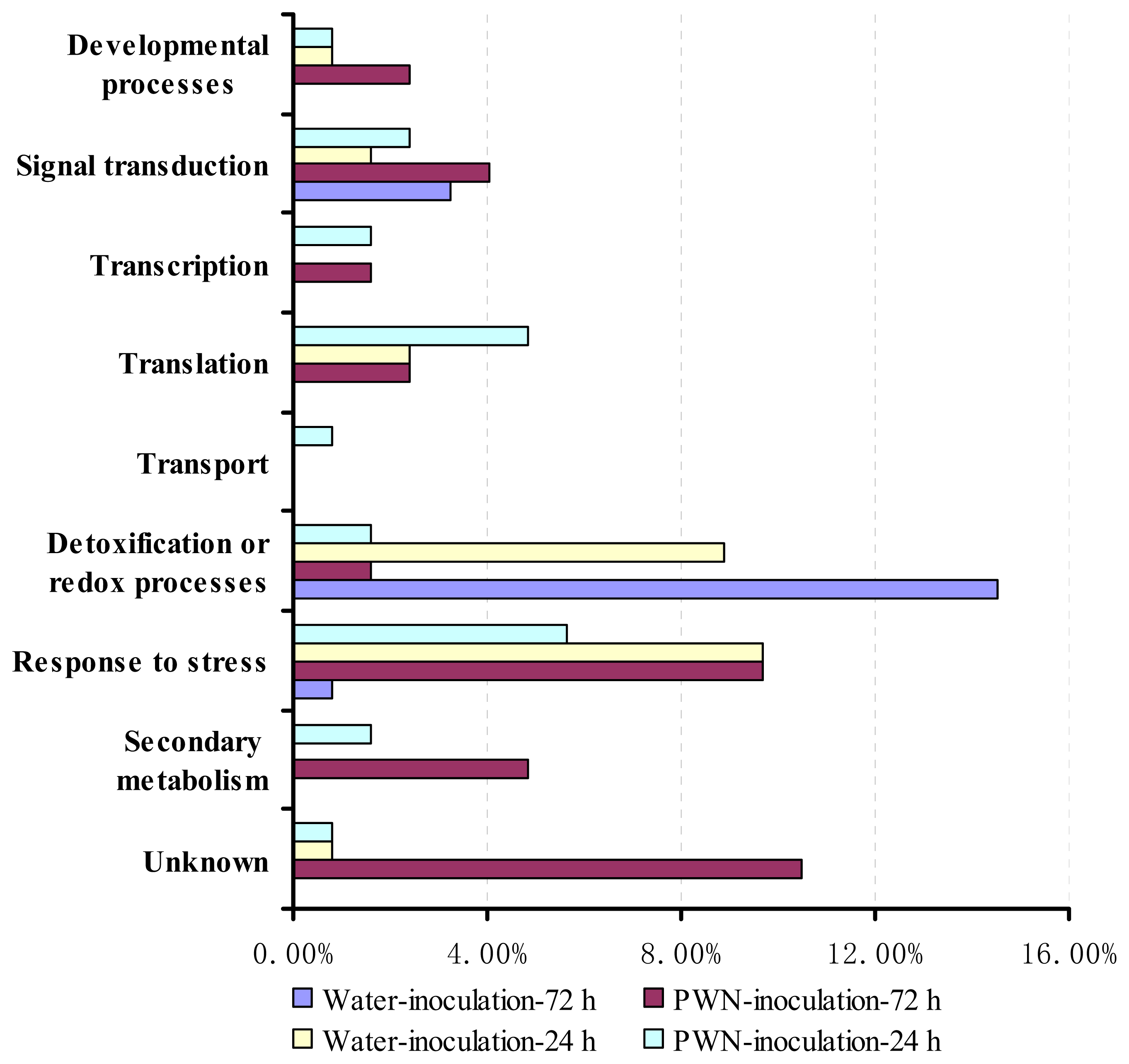

{kind=link}
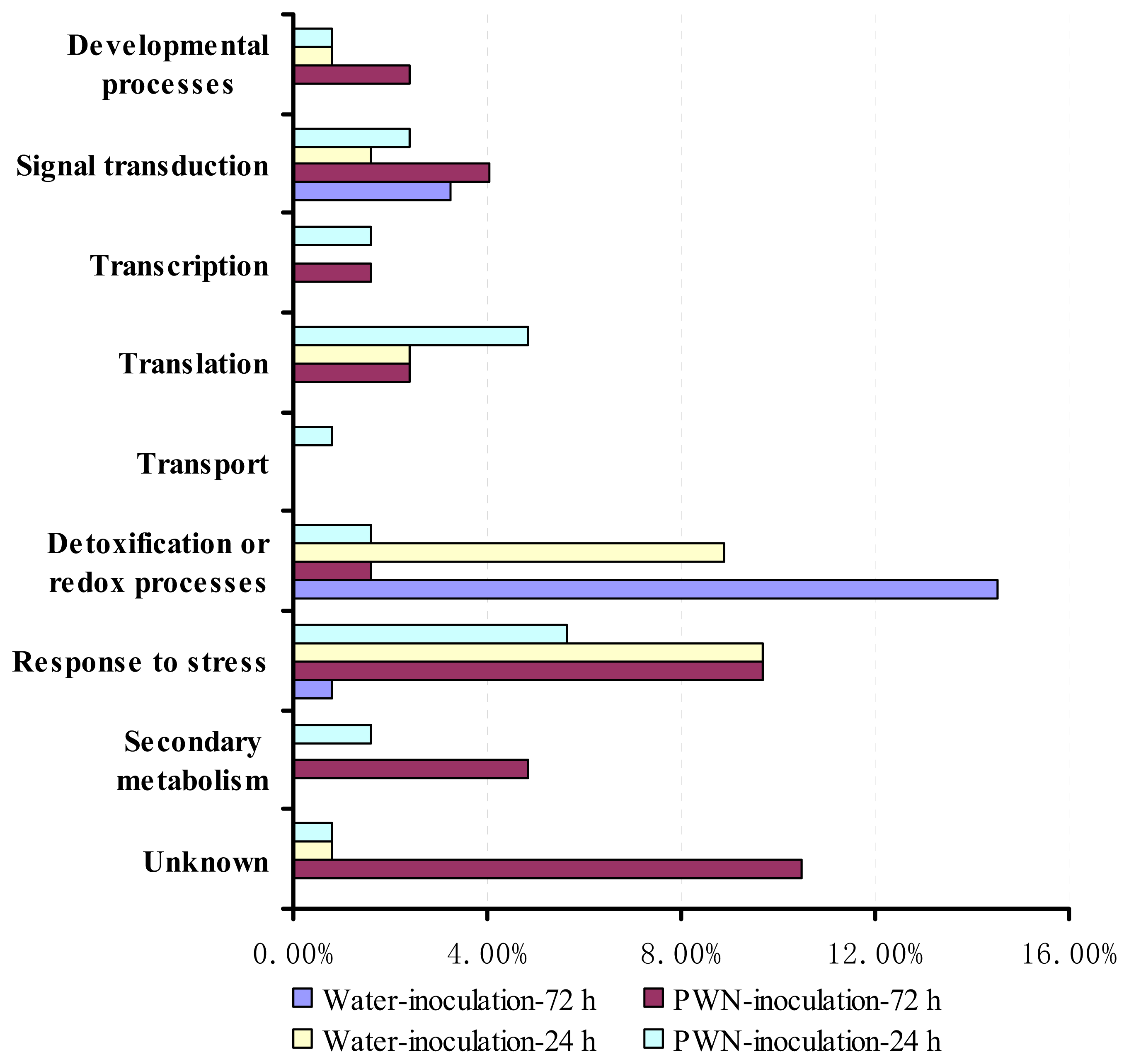{kind=link}
{kind=link}
| PM-24 h | PM-72 h | |
|---|---|---|
| Library and EST summary | ||
| Number of cDNAs sequenced | 61 | 83 |
| Mean read length 1 | 458.5 | 427.6 |
| Number of high-quality ESTs | 55 | 69 |
| Clustering results | ||
| Number of assembled ESTs 2 | 23 | 24 |
| Number of contigs (A) | 5 | 4 |
| Number of singletons (B) | 32 | 45 |
| Number of assembled sequences (A + B) | 37 | 49 |
| Contig sizes | ||
| 2–4 ESTs 3 | 3 | 2 |
| 5–7 ESTs | 1 | 1 |
| 8–13 ESTs | 1 | 0 |
| >14 ESTs | 0 | 1 |
| PM-24h | PM-72h | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Clone ID | BLASTX | E Value | Signal intensity ratio | Clustered EST No. | Clone ID | BLASTX | E Value | Signal intensity ratio | Clustered EST No. |
| Developmental process | |||||||||
| PMXY 148 | Pectin methylesterase 6 Arabidopsis AT4G02330.1 | 6 × 10−84 | 2.07 (Upregulated) | PMXY 165 | No pollen germination 1 Arabidopsis AT2G43040.1 | 1 × 10−7 | 2.06 (Upregulated) | ||
| PMXY 120 | Polygalacturonase Arabidopsis AT1G80170.1 | 4 × 10−18 | 0.10 (Downregulated) | PMXY 155 | Acyl carrier protein 4 Arabidopsis AT4G25050.1 | 9 × 10−28 | 2.30 (Upregulated) | ||
| PMXY 186 | Aldehyde dehydrogenase 11A3 Arabidopsis AT2G24270.4 | 2 × 10−15 | 2.02 (Upregulated) | ||||||
| Signal transduction | |||||||||
| PMXY 121 | Jasmonate ZIM domain-containing Arabidopsis AT1G74950.1 | 2 × 10−3 | 2.14 (Upregulated) | Contig 01 | Calcium-binding protein Arabidopsis AT2G46600.1 | 2 × 10−25 | 0.11 (Downregulated) | 4 | |
| PMXY 131 | Calreticulin 1a Arabidopsis AT1G56340.2 | 1 × 10−123 | 2.10 (Upregulated) | PMXY 153 | Calcium-binding protein Arabidopsis AT2G46600.1 | 3 × 10−12 | 2.48 (Upregulated) | ||
| PMXY 130 | Phosphate-responsive protein Arabidopsis AT4G08950.1 | 1 × 10−36 | 2.10 (Upregulated) | PMXY 205 | Jasmonate ZIM domain-containing Arabidopsis AT1G19180.2 | 8 × 10−3 | 2.47 (Upregulated) | ||
| PMXY 099 | Phosphoesterase family protein Arabidopsis AT2G26870.1 | 4 × 10−28 | 0.08 (Downregulated) | PMXY 160 | Response regulator 3 Arabidopsis AT1G59940.1 | 1 × 10−35 | 2.01 (Upregulated) | ||
| PMXY 108 | Phosphatidylinositol n-acetylglucosaminyltransferase subunit p Arabidopsis AT2G39445.1 | 2 × 10−37 | 0.12 (Downregulated) | PMXY 167 | RAB GTPase homolog A2D Arabidopsis AT5G59150.1 | 3 × 10−29 | 2.19 (Upregulated) | ||
| PMXY 210 | Phosphate-responsive protein Arabidopsis AT4G08950.1 | 2 × 10−40 | 2.05 (Up-regulated) | ||||||
| Transcription | |||||||||
| PMXY 151 | PHD finger family protein Arabidopsis AT2G02470.2 | 3 × 10−48 | 2.03 (Upregulated) | PMXY 173 | C3HC4-type RING finger protein Arabidopsis AT5G22920.1 | 4 × 10−20 | 2.15 (Upregulated) | ||
| PMXY 147 | Zinc finger protein (C2H2 type) Arabidopsis AT2G28200.1 | 1 × 10−3 | 2.43 (Upregulated) | PMXY 192 | La domain-containing protein Arabidopsis AT4G32720.2 | 8 × 10−17 | 2.30 (Upregulated) | ||
| Translation | |||||||||
| PMXY 139 | 60S ribosomal protein L10 Arabidopsis AT1G26910.1 | 2 × 10−38 | 2.17 (Upregulated) | PMXY 204 | 60S ribosomal protein L38 Arabidopsis AT3G59540.1 | 2 × 10−8 | 2.65 (Upregulated) | ||
| PMXY 126 | 60S ribosomal protein L24 Arabidopsis AT2G36620.1 | 1 × 10−51 | 2.33 (Upregulated) | PMXY 212 | Serine-rich proteins Arabidopsis AT3G56500.1 | 8 × 10−7 | 2.15 (Upregulated) | ||
| PMXY 141 | 60S ribosomal protein L29 Arabidopsis AT5G02610.1 | 5 × 10−54 | 2.08 (Upregulated) | PMXY 185 | GTP-binding Elongation factor Tu family protein Arabidopsis AT5G60390.3 | 6 × 10−86 | 2.04 (Upregulated) | ||
| PMXY 142 | 60S ribosomal protein L27a-2 Arabidopsis AT1G23290.1 | 1 × 10−51 | 2.37 (Upregulated) | ||||||
| PMXY 133 | 40S ribosomal protein S11-2 Arabidopsis AT3G48930.1 | 1 × 10−41 | 2.23 (Upregulated) | ||||||
| PMXY 140 | Translation initiation factor eIF-4A Arabidopsis AT3G13920.4 | 1 × 10−142 | 2.50 (Upregulated) | ||||||
| PMXY 098 | RNA-binding protein Nova-1-like Arabidopsis AT5G04430.1 | 5 × 10−9 | 0.09 (Down-regulated) | ||||||
| PMXY 089 | RNA recognition motif (RRM)-containing protein Arabidopsis AT1G14340.1 | 3 × 10−5 | 0.08 (Down-regulated) | ||||||
| PMXY 107 | 60S ribosomal protein L15 Arabidopsis AT4G16720.1 | 2 × 10−7 | 0.05 (Down-regulated) | ||||||
| Transport | |||||||||
| PMXY 124 | TOM1-like protein 2-like isoform 2 Arabidopsis AT4G32760.2 | 4 × 10−24 | 2.31 (Upregulated) | ||||||
| Detoxification or redox processes | |||||||||
| Contig 02 | Metallothionein 3 Arabidopsis AT3G15353.1 | 6 × 10−9 | 0.04 (Downregulated) | 11 | Contig 03 | Heavy-metal-associated domaincontaining protein Arabidopsis AT1G01490.2 | 1 × 10−6 | 0.10 (Downregulated) | 3 |
| PMXY 134 | Glutathione peroxidase Arabidopsis AT4G11600.1 | 2 × 10−71 | 2.48 (Upregulated) | Contig 04 | Metallothionein 3 Arabidopsis AT3G15353.1 | 6 × 10−9 | 0.09 (Downregulated) 15 | ||
| PMXY 136 | 2-alkenal reductase Arabidopsis AT5G16970 | 3 × 10−23 | 2.05 (Upregulated) | PMXY 168 | Glyoxalase 1 family protein Arabidopsis AT1G15380.2 | 3 × 10−12 | 2.29 (Upregulated) | ||
| PMXY 170 | Cyt_b561_FRRS1-likecontaining protein Picea sitchensis ADM73751.1 | 2 × 10−4 | 2.04 (Upregulated) | ||||||
| Response to stress | |||||||||
| Contig 05 | Thaumatin-like family protein Arabidopsis AT4G11650.1 | 4 × 10−15 | 2.55 (Upregulated) | 2 | Contig 09 | Mannose/glucose-specific lectin Arabidopsis AT1G19715.3 | 8 × 10−14 | 1.73 (Upregulated) | 2 |
| Contig 06 | Heat shock protein 18.2 Arabidopsis AT5G59720.1 | 6 × 10−42 | 0.10 (Downregulated) | 2 | PMXY 172 | Heat shock protein 70 Arabidopsis AT5G02500.1 | 8 × 10−59 | 2.06 (Upregulated) | |
| Contig 07 | Calcium-binding protein lp8 Arabidopsis AT1G24620.1 | 9 × 10−20 | 0.04 (Downregulated) | 5 | PMXY 190 | AAA-type ATPase family protein Arabidopsis AT4G02480.1 | 2 × 10−21 | 2.21 (Upregulated) | |
| Contig 08 | Drought stress responsive protein Pinus pinaster AJ309123.1 | 6 × 10−97 | 0.10 (Downregulated) | 3 | PMXY 189 | Cell-wall protein (lp5) Pinus sylvestris EU394126.1 | 2 × 10−4 | 2.42 (Upregulated) | |
| PMXY 127 | Thaumatin-like family protein Arabidopsis AT4G11650.1 | 3 × 10−16 | 5.60 (Upregulated) | PMXY 156 | BCL-2-associated athanogene 6 Arabidopsis AT2G46240.1 | 3 × 10−6 | 2.14 (Upregulated) | ||
| PMXY 135 | Dirigent-like protein Arabidopsis AT4G11190.1 | 0.77 | 2.39 (Upregulated) | PMXY 195 | Ubiquitin family protein Arabidopsis AT4G02890.4 | 1 × 10−111 | 2.19 (Upregulated) | ||
| PMXY 128 | Chitin recognition protein Arabidopsis AT3G04720.1 | 1 × 10−15 | 8.41 (Upregulated) | PMXY 196 | Ubiquitin-like conjugating enzyme Arabidopsis AT1G64230.5 | 6 × 10−52 | 2.23 (Upregulated) | ||
| PMXY 146 | Chitinase Arabidopsis AT2G43590.1 | 2 × 10−20 | 2.45 (Upregulated) | PMXY 158 | Lipid transfer protein Arabidopsis AT5G59310.1 | 1 × 10−11 | 2.47 (Upregulated) | ||
| PMXY 145 | Heat shock protein 90 Arabidopsis AT5G52640.1 | 1 × 10−65 | 2.33 (Upregulated) | PMXY 209 | Adenine nucleotide alpha hydrolase-like superfamily protein Arabidopsis AT5G14680.1 | 6 × 10−43 | 2.11 (Upregulated) | ||
| PMXY 103 | High mobility group B3 Arabidopsis AT1G20696.3 | 1 × 10−31 | 0.03 (Downregulated) | PMXY 201 | RPS 2 protein Arabidopsis AT4G26090.1 | 6 × 10−6 | 2.04 (Upregulated) | ||
| PMXY 088 | Heat shock protein 101 Arabidopsis AT1G74310.1 | 6 × 10−10 | 0.11 (Downregulated) | PMXY 181 | Pentatricopeptide (PPR) repeatcontaining protein Arabidopsis AT2G22070.1 | 3 × 10−22 | 2.35 (Upregulated) | ||
| PMXY 214 | Aluminum-induced protein with YGL and LRDR motifs Arabidopsis AT5G19140.1 | 1 × 10−33 | 0.14 (Downregulated) | ||||||
| Secondary metabolism | |||||||||
| PMXY 149 | Methionine synthase 2 Arabidopsis AT3G03780.3 | 1 × 10−120 | 2.28 (Upregulated) | PMXY 166 | Methionine synthase 2 Arabidopsis AT3G03780.3 | 3 × 10−59 | 2.52 (Upregulated) | ||
| PMXY 129 | FAD/NAD(P)-binding oxidoreductase family protein Arabidopsis AT2G35660.1 | 1 × 10−22 | 2.11 (Upregulated) | PMXY 202 | Caffeic acid Omethyltransferase Pinus taeda AAC49708.1 | 2 × 10−12 | 2.08 (Upregulated) | ||
| PMXY 152 | Cinnamyl alcohol dehydrogenase 5 Arabidopsis AT4G34230.1 | 9 × 10−48 | 2.42 (Upregulated) | ||||||
| PMXY 187 | Chalcone and stilbene synthase family protein Arabidopsis AT5G13930.1 | 1 × 10−35 | 2.17 (Upregulated) | ||||||
| PMXY 211 | Caffeoyl-CoA O-methyltransferase Pinus taeda AF036095.1 | 3 × 10−76 | 2.19 (Upregulated) | ||||||
| PMXY 183 | Phenylcoumaran benzylic ether reductase Pinus taeda AF242490.2 | 1 × 10−94 | 2.26 (Upregulated) | ||||||
| Candidate gene | Primer sequence |
|---|---|
| Heat-shock protein 18.2 (HSP 18.2) | 5′-TCATACCGCGTGAGAGGTCAA-3′ 5′-AAGGCGAGCATGGAAAACG-3′ |
| Cinnamyl alcohol dehydrogenase (CAD) | 5′-AGCATGGAGGAAGCACAGGAA-3′ 5′-TCCATGGCCGTGTTGATGTAG-3′ |
| No pollen germination 1 (NPG1) | 5′-TTGGAGCTGTATCATGCAGCC-3′ 5′-CACCAGTTGACCAATGGAAGC-3′ |
| Chalcone and stilbene synthase family protein (CHS) | 5′-GGATCCAGATTCAACTTCGCC-3′ 5′-TGGTTGAGGCATTCCAGCA-3′ |
| 18S rRNA | 5′-CGGCTACCACATCCAAGGAA-3′ 5′-GCTGGAATTACCGCGGCT-3′ |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xu, L.; Liu, Z.-Y.; Zhang, K.; Lu, Q.; Liang, J.; Zhang, X.-Y. Characterization of the Pinus massoniana Transcriptional Response to Bursaphelenchus xylophilus Infection Using Suppression Subtractive Hybridization. Int. J. Mol. Sci. 2013, 14, 11356-11375. https://doi.org/10.3390/ijms140611356
Xu L, Liu Z-Y, Zhang K, Lu Q, Liang J, Zhang X-Y. Characterization of the Pinus massoniana Transcriptional Response to Bursaphelenchus xylophilus Infection Using Suppression Subtractive Hybridization. International Journal of Molecular Sciences. 2013; 14(6):11356-11375. https://doi.org/10.3390/ijms140611356
Chicago/Turabian StyleXu, Liang, Zhen-Yu Liu, Kai Zhang, Quan Lu, Jun Liang, and Xing-Yao Zhang. 2013. "Characterization of the Pinus massoniana Transcriptional Response to Bursaphelenchus xylophilus Infection Using Suppression Subtractive Hybridization" International Journal of Molecular Sciences 14, no. 6: 11356-11375. https://doi.org/10.3390/ijms140611356