Phthalic Acid Chemical Probes Synthesized for Protein-Protein Interaction Analysis


Abstract
:1. Introduction
2. Results and Discussion
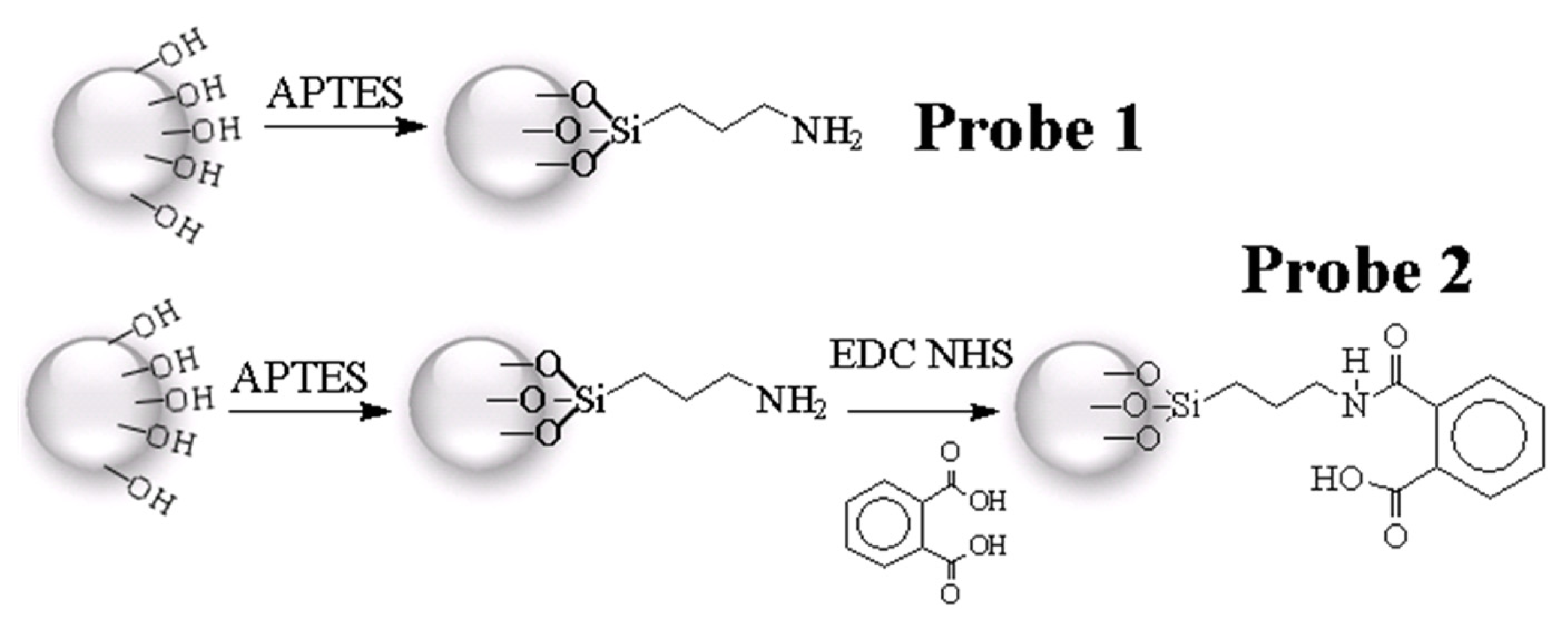
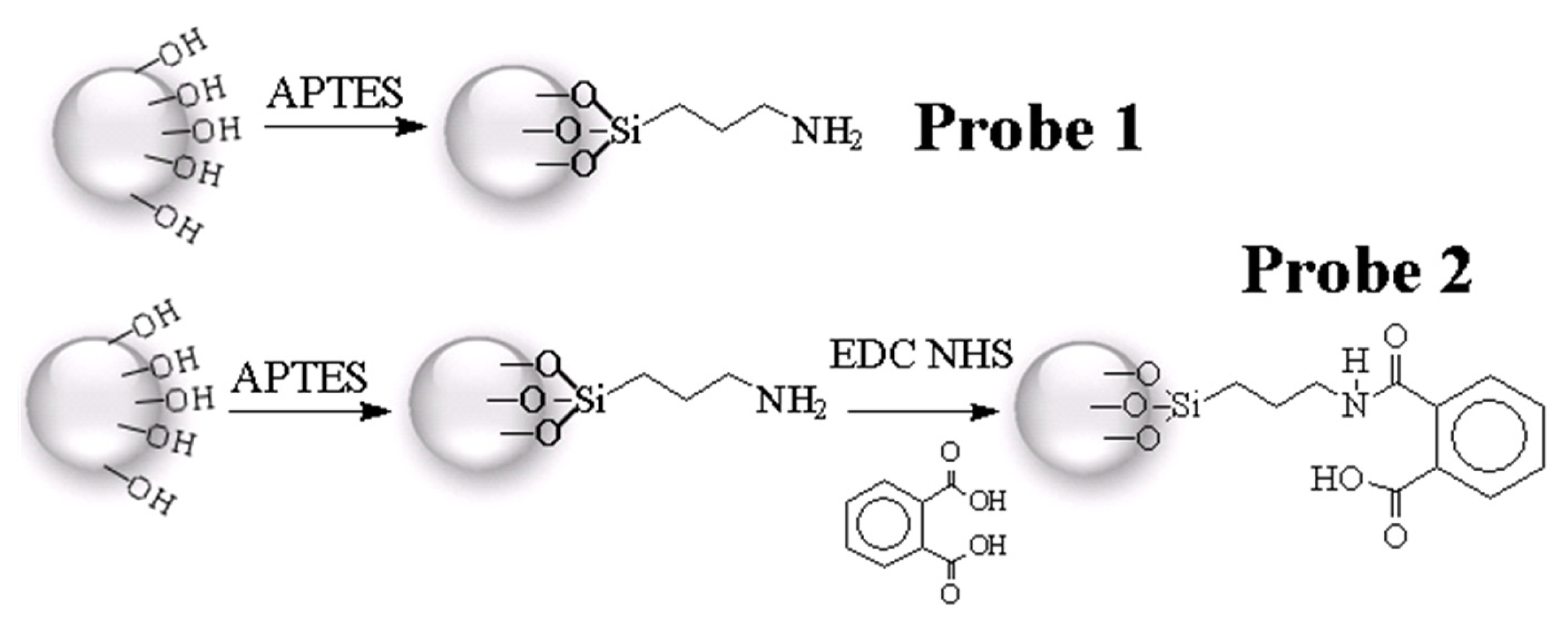
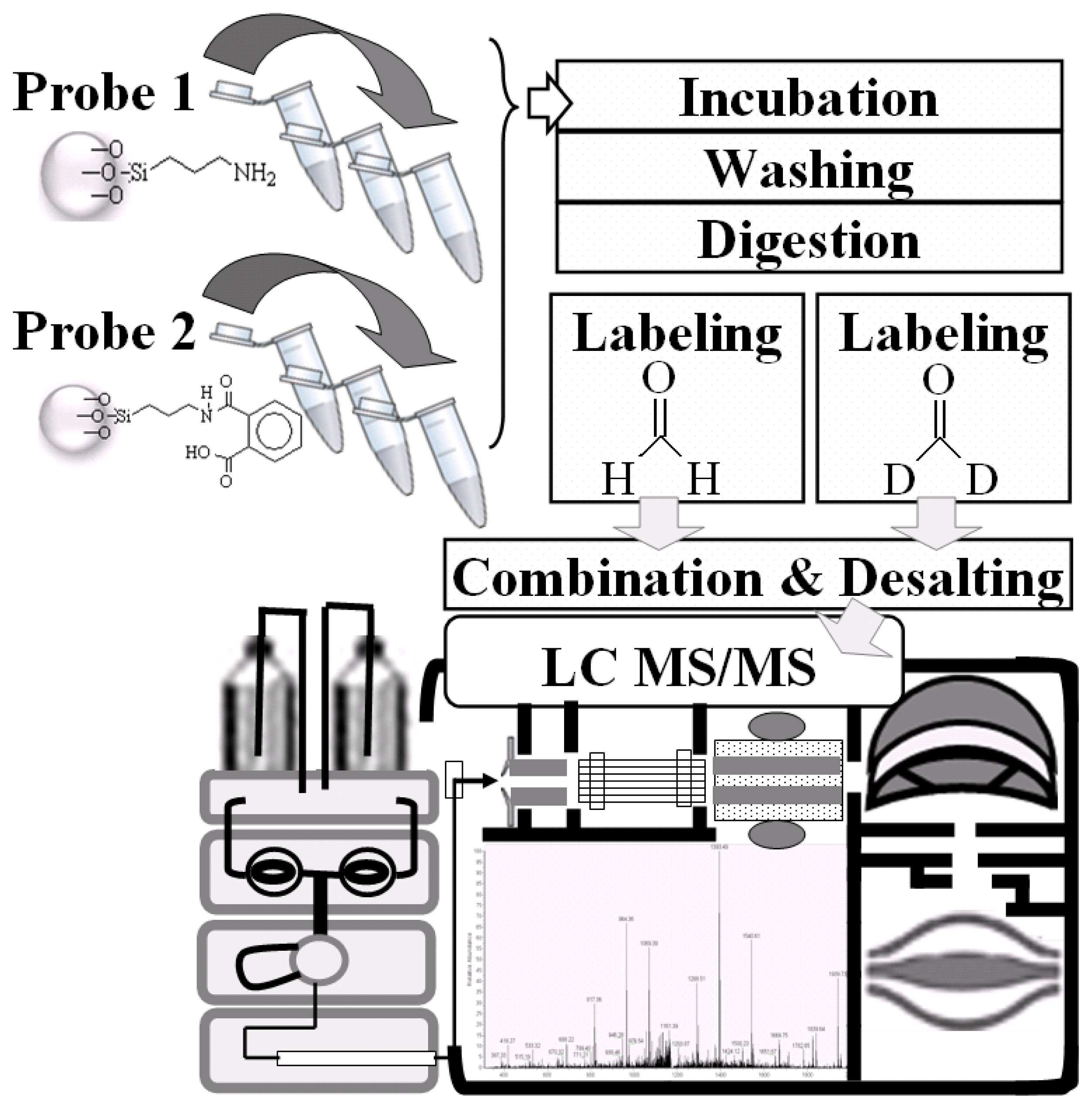
2.1. Procedures for Chemical Probe Synthesis
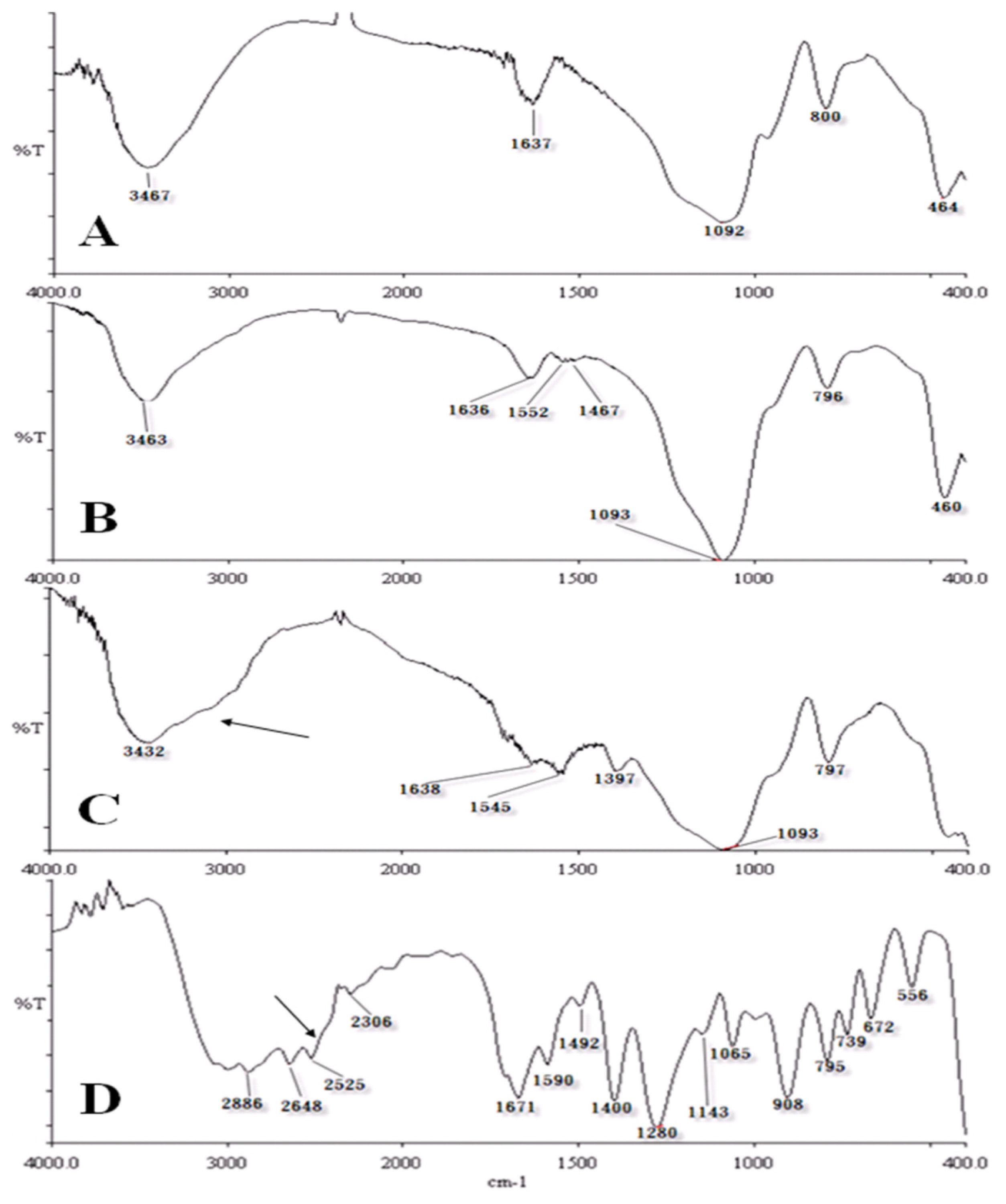
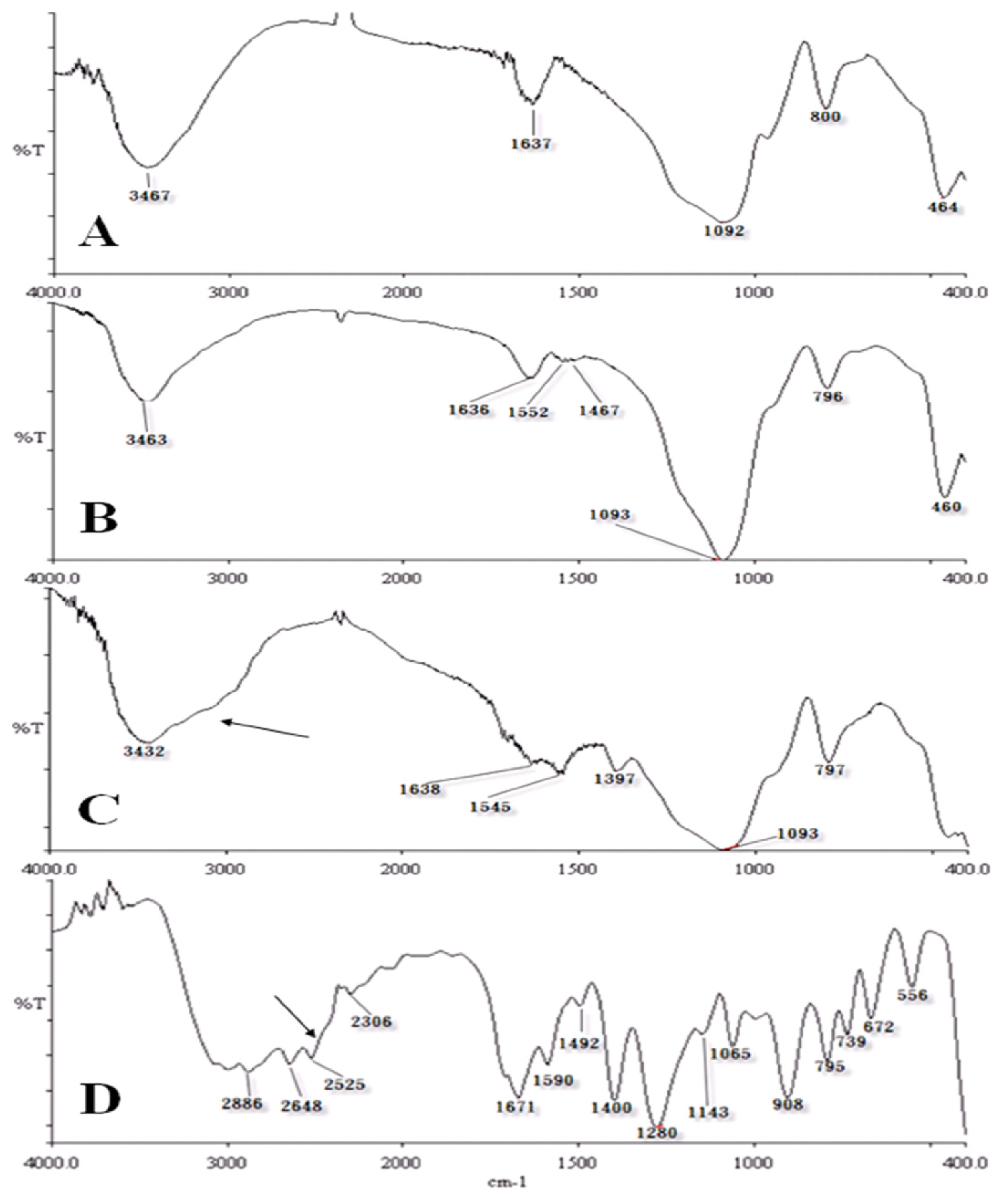
2.2. Characterization of Chemical Probes by IR Spectrometry
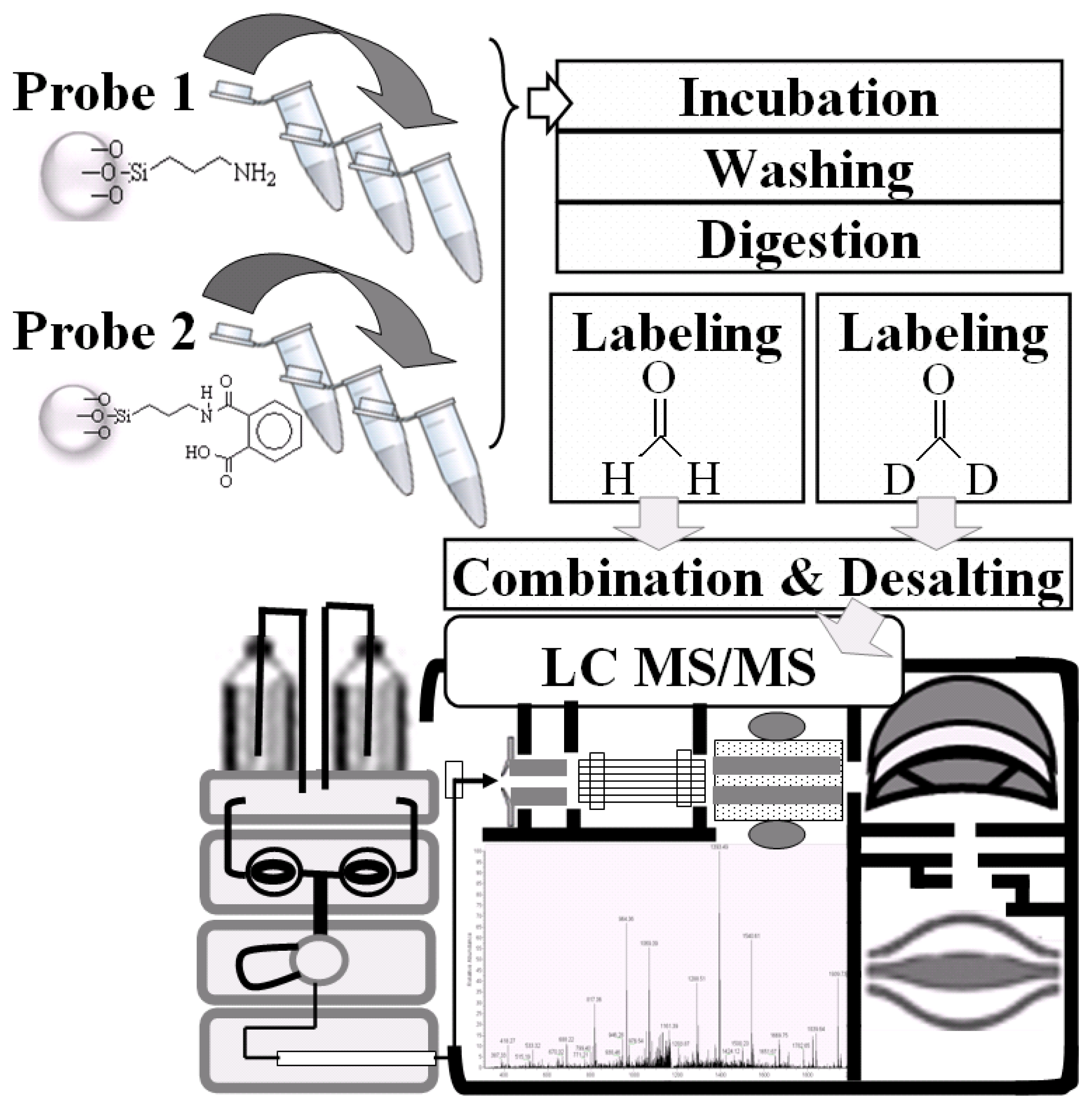
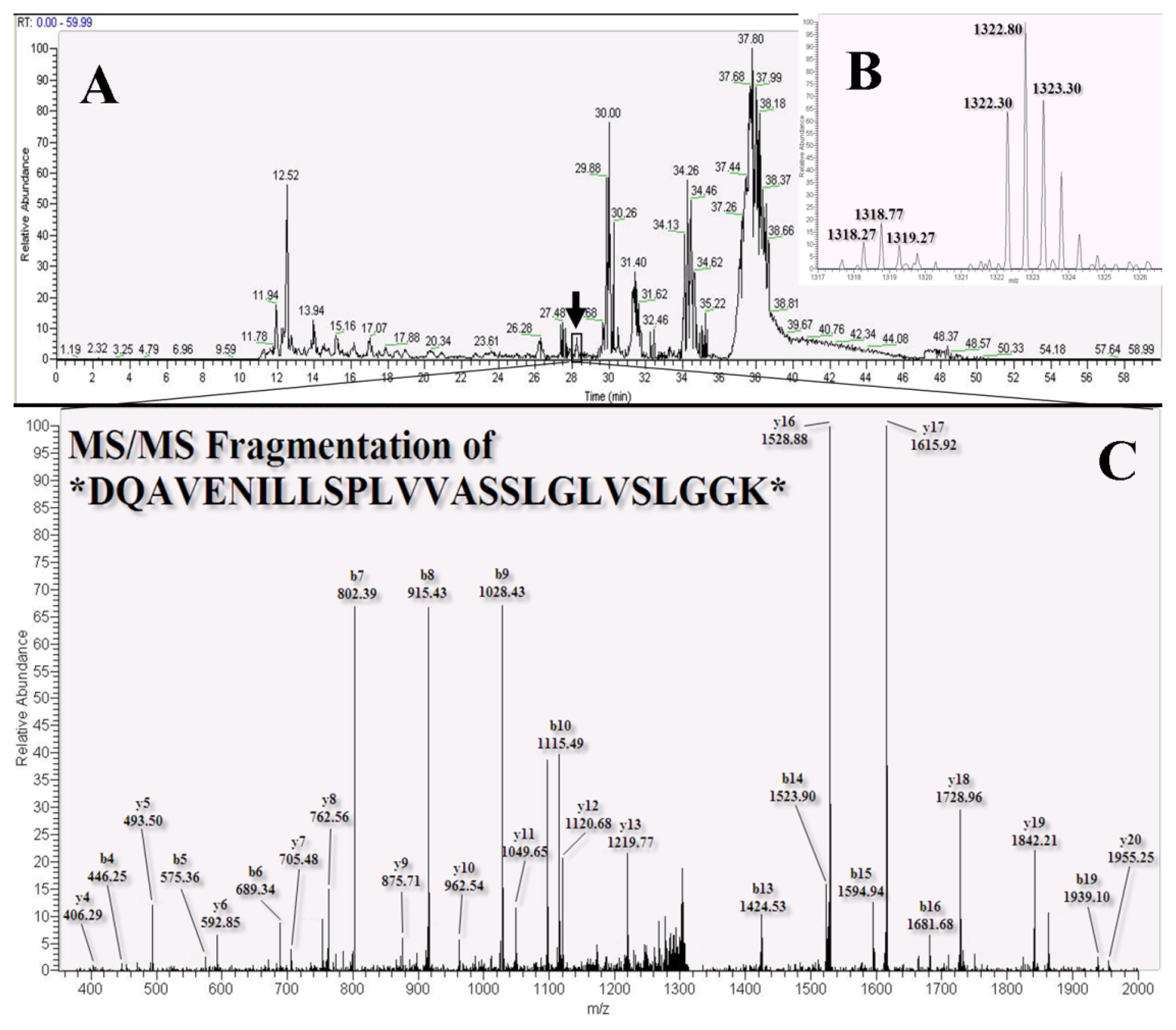
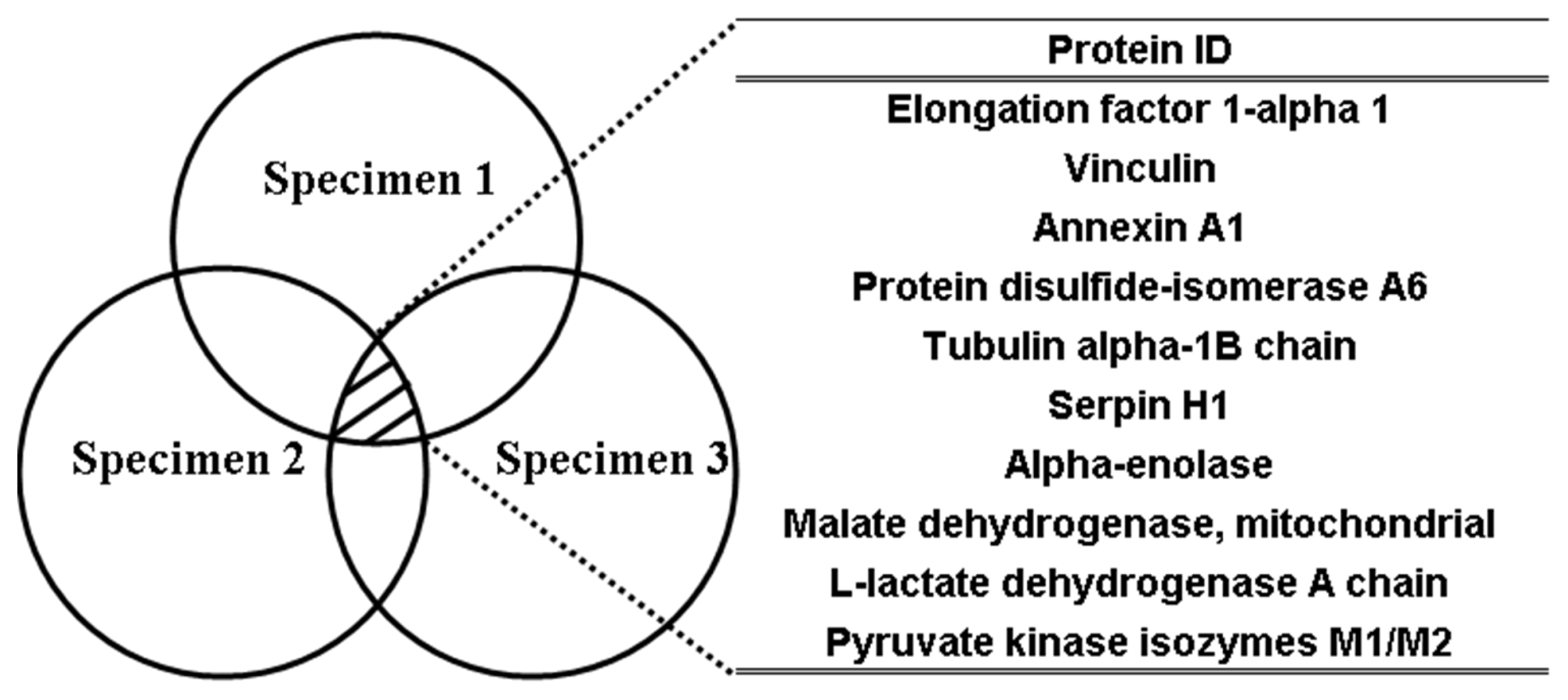
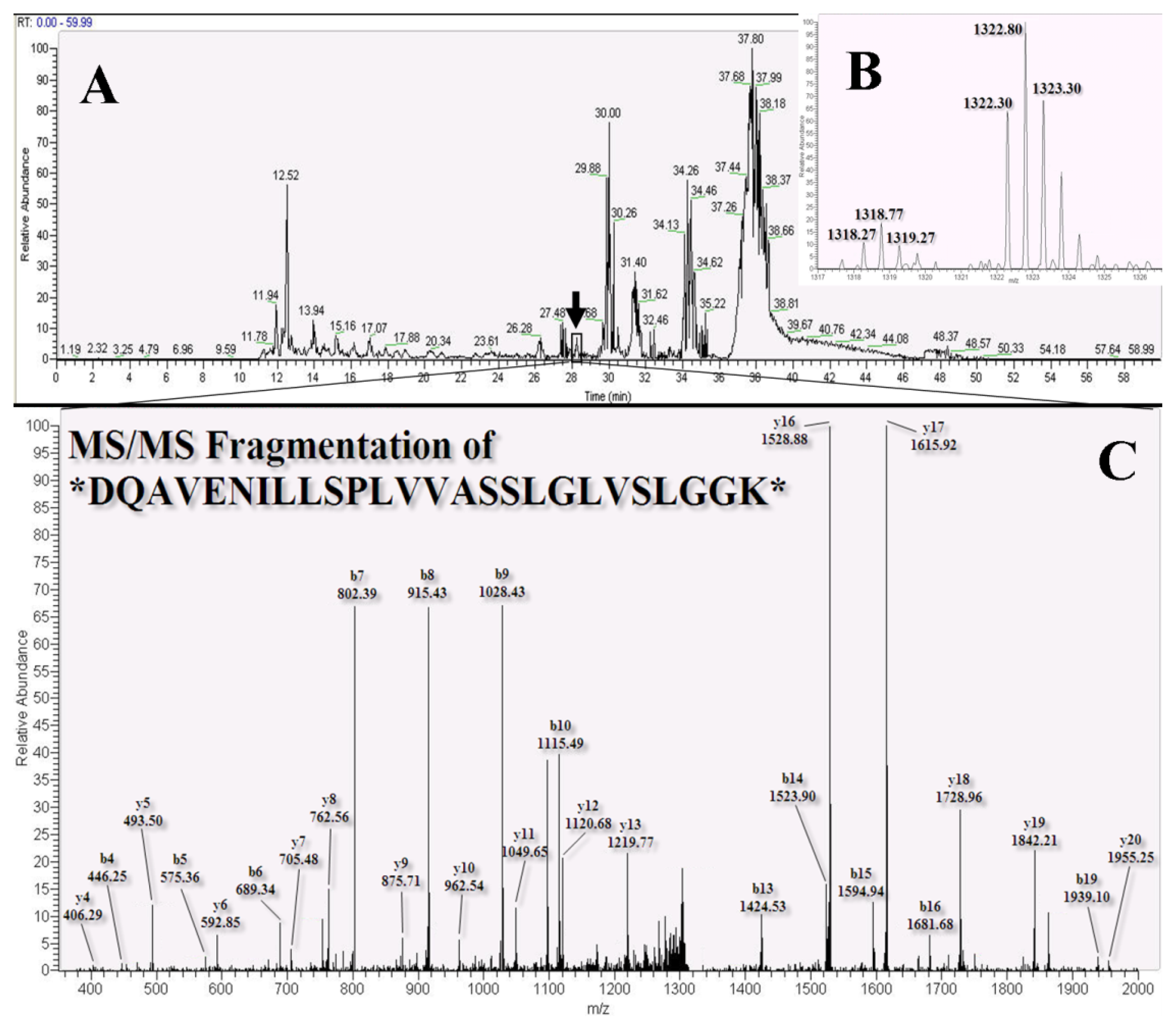
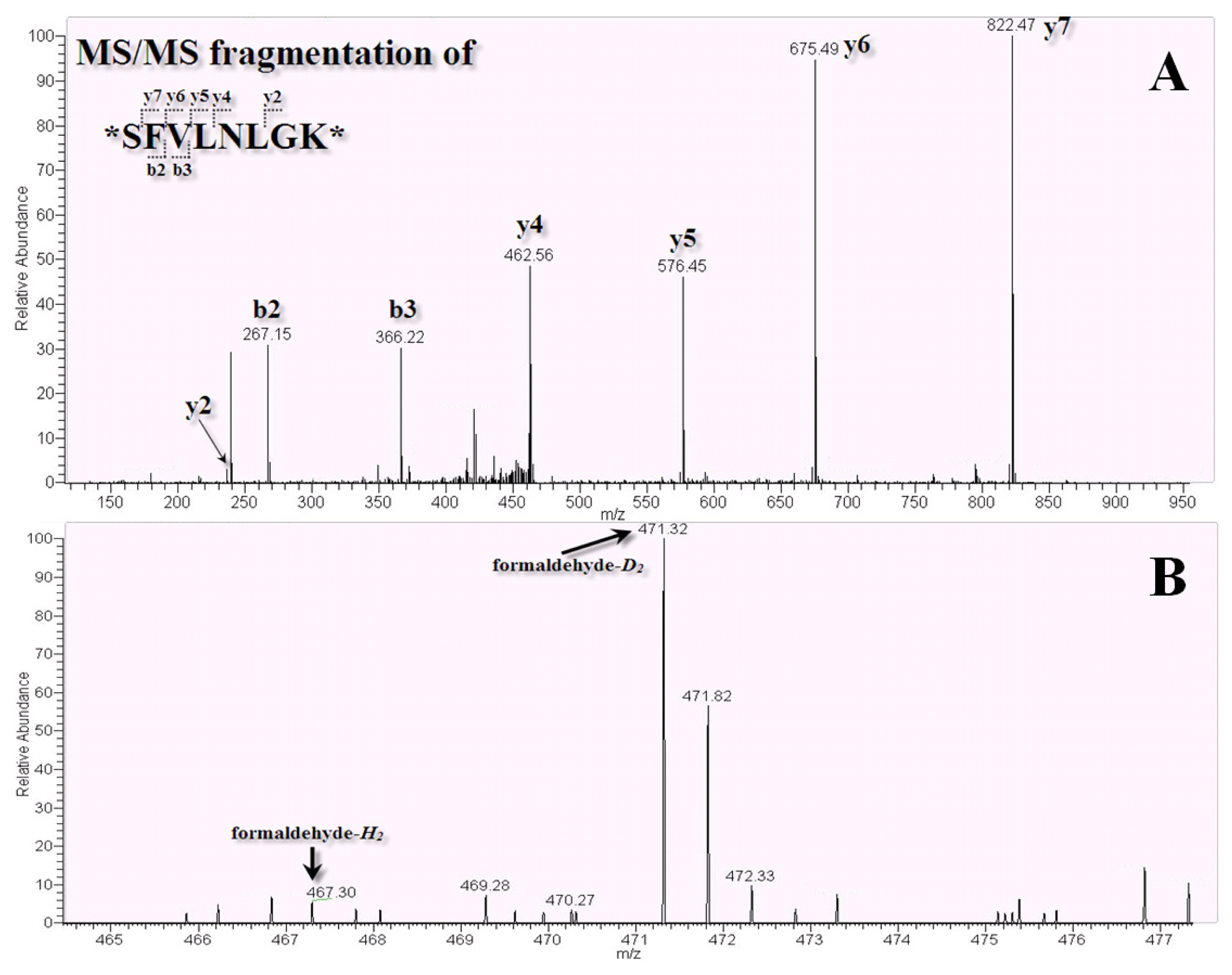
2.3. LC-MS/MS for Identification and Quantitation of Bound Proteins
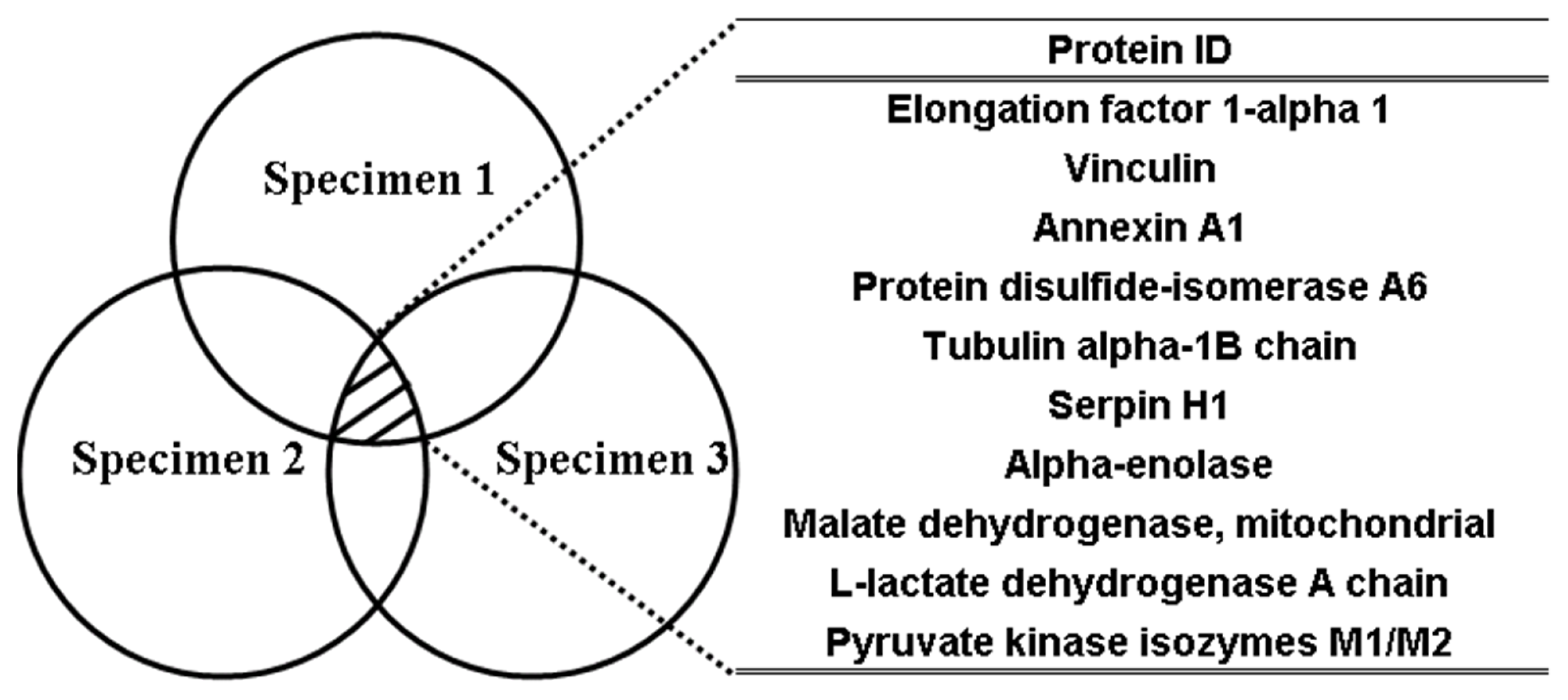
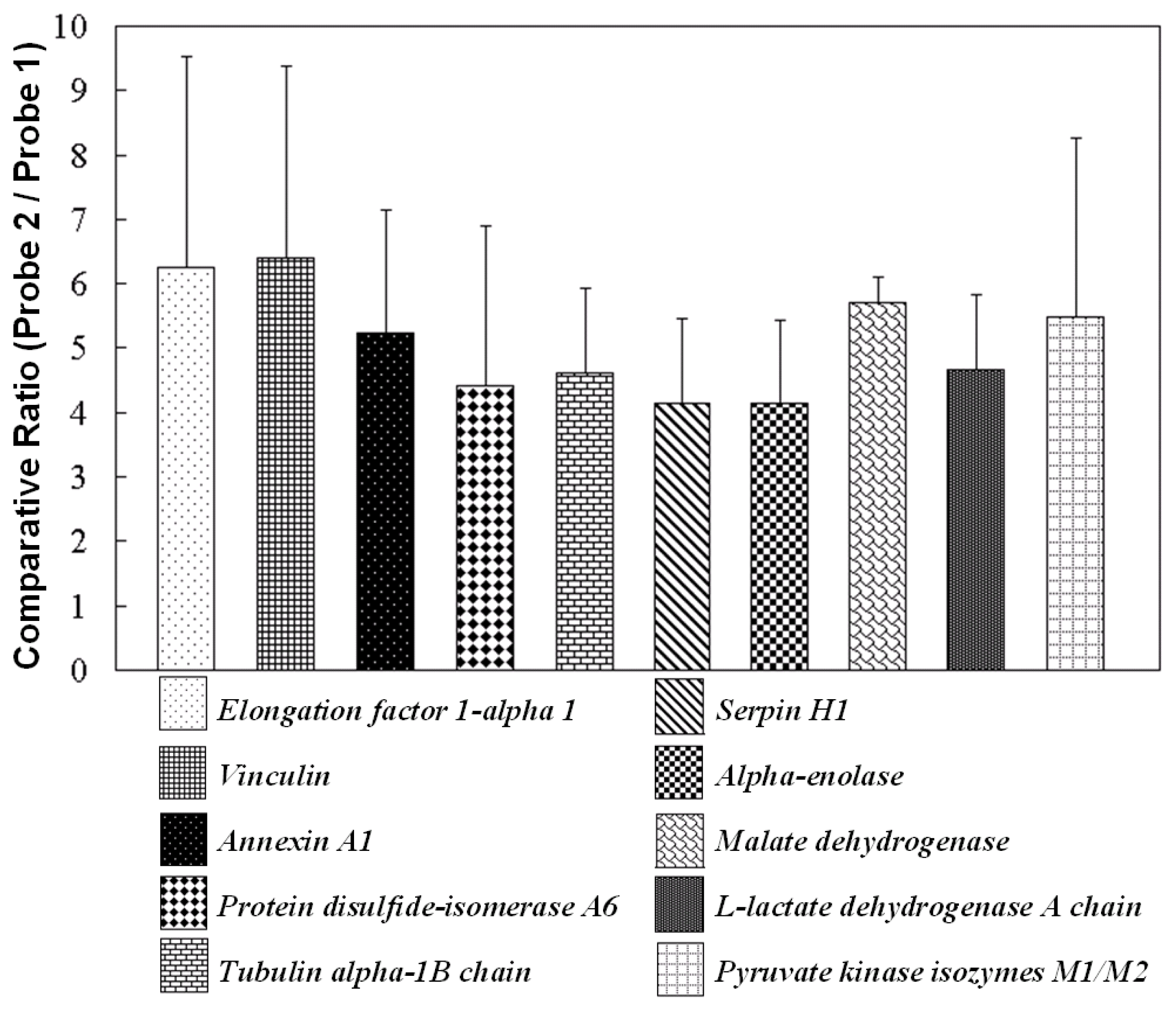
2.4. Functions of Phthalate Chemical Probe Binding Proteins
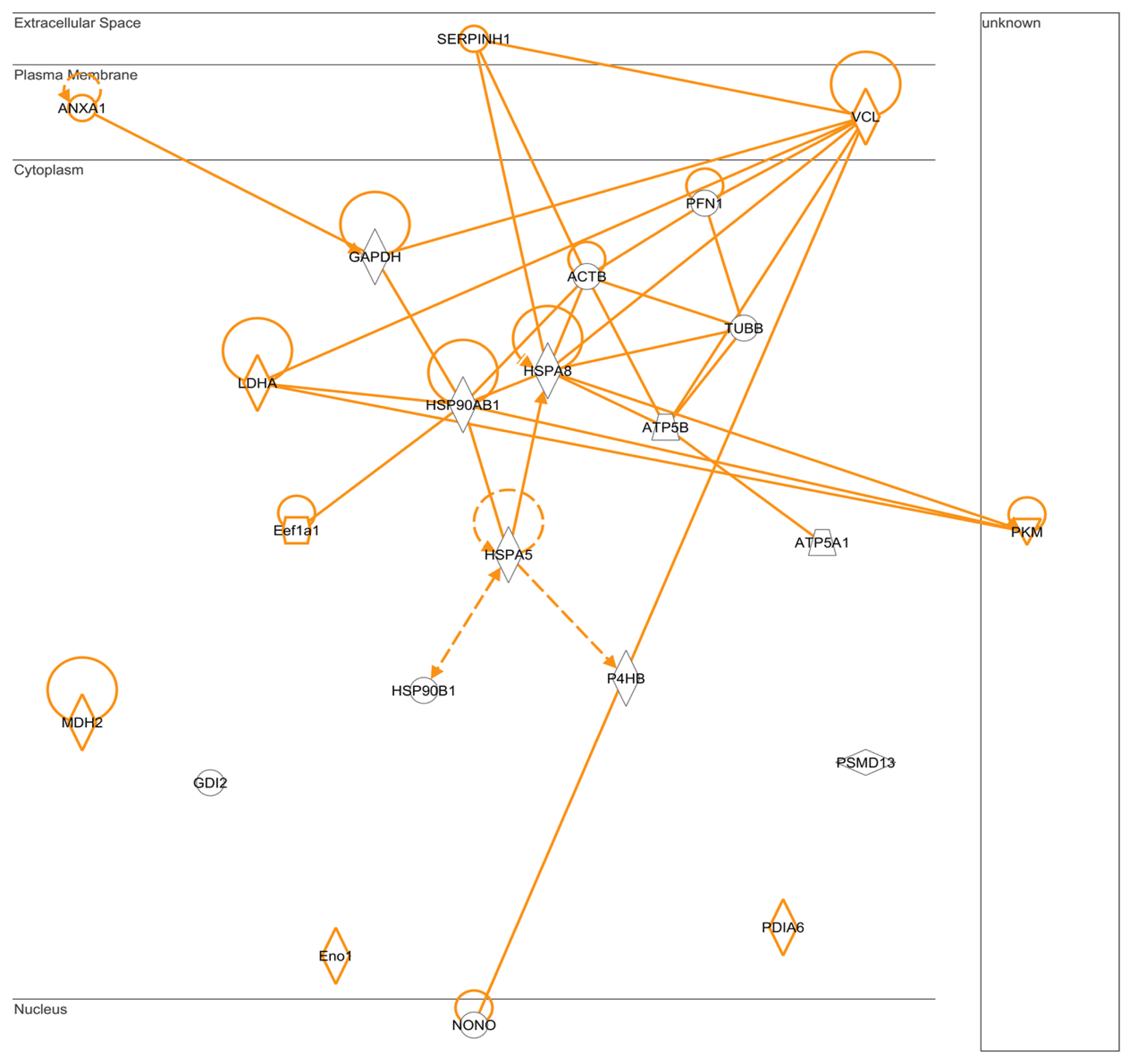
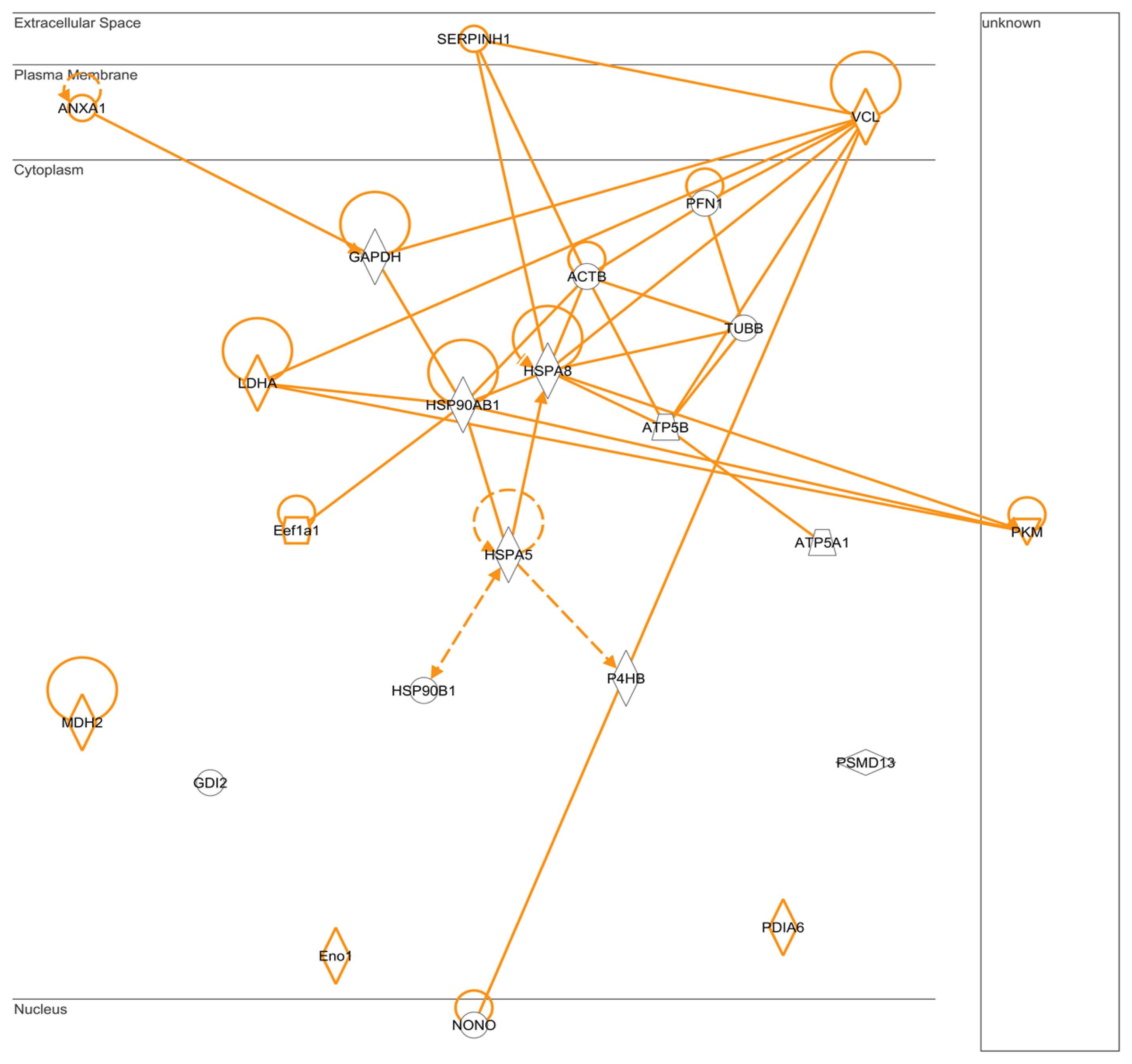
2.5. IPA: Pathways for Protein-Protein Interactions
3. Methods and Experimental Section
3.1. Materials and Chemicals
3.2. Phthalic Acid Chemical Probes Syntheses
3.3. Characterization of Chemical Probes by Infrared (IR) Spectrometry
3.4. NRK-52E Cell Culture and Protein Concentration
3.5. Conditions for NRK-52E Cell Lysate with APTES-Modified and Phthalic Acid Chemical Probes
3.6. Direct Tryptic Digestion and Dimethyl Labeling
3.7. Nano-LC-MS/MS Analysis and Mascot Database Search
3.8. Pathways and Networks Analyses of Protein-Protein Interactions
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Nassberger, L.; Arbin, A.; Ostelius, J. Exposure of patients to phthalates from polyvinyl chloride tubes and bags during dialysis. Nephron 1987, 45, 286–290. [Google Scholar]
- Mettang, T.; Alscher, D.M.; Pauli-Magnus, C.; Dunst, R.; Kuhlmann, U.; Rettenmeier, A.W. Phthalic acid is the main metabolite of the plasticizer di(2-ethylhexyl) phthalate in peritoneal dialysis patients. Adv. Perit. Dial 1999, 15, 229–233. [Google Scholar]
- Silva, M.J.; Slakman, A.R.; Reidy, J.A.; Preau, J.L., Jr; Herbert, A.R.; Samandar, E.; Needham, L.L.; Calafat, A.M. Analysis of human urine for fifteen phthalate metabolites using automated solid-phase extraction. J. Chromatogr. B 2004, 805, 161–167. [Google Scholar]
- Chen, M.; Tao, L.; Collins, E.M.; Austin, C.; Lu, C. Simultaneous determination of multiple phthalate metabolites and bisphenol-A in human urine by liquid chromatography-tandem mass spectrometry. J. Chromatogr. B 2012, 904, 73–80. [Google Scholar]
- Calafat, A.M.; Ye, X.; Wong, L.Y.; Reidy, J.A.; Needham, L.L. Exposure of the U.S. population to bisphenol A and 4-tertiary-octylphenol: 2003–2004. Environ. Health Perspect 2008, 116, 39–44. [Google Scholar]
- Tsai, M.J.; Kuo, P.L.; Ko, Y.C. The association between phthalate exposure and asthma. Kaohsiung J. Med. Sci 2012, 28, S28–S36. [Google Scholar]
- Hsieh, T.H.; Tsai, C.F.; Hsu, C.Y.; Kuo, P.L.; Hsi, E.; Suen, J.L.; Hung, C.H.; Lee, J.N.; Chai, C.Y.; Wang, S.C.; et al. n-Butyl benzyl phthalate promotes breast cancer progression by inducing expression of lymphoid enhancer factor 1. PLoS One 2012, 7, e42750. [Google Scholar]
- Koo, H.J.; Lee, B.M. Estimated exposure to phthalates in cosmetics and risk assessment. J. Toxicol. Environ. Health A 2004, 67, 1901–1914. [Google Scholar]
- Zhao, B.; Chu, Y.; Huang, Y.; Hardy, D.O.; Lin, S.; Ge, R.S. Structure-dependent inhibition of human and rat 11β-hydroxysteroid dehydrogenase 2 activities by phthalates. Chem. Biol. Interact 2010, 183, 79–84. [Google Scholar]
- Latini, G.; Verrotti, A.; de Felice, C. DI-2-ethylhexyl phthalate and endocrine disruption: A review. Curr. Drug Targets Immune Endocr. Metabol. Disord 2004, 4, 37–40. [Google Scholar]
- Singh, S.; Li, S.S. Phthalates: Toxicogenomics and inferred human diseases. Genomics 2011, 97, 148–157. [Google Scholar]
- Kumar, D.M.; Thota, B.; Shinde, S.V.; Prasanna, K.V.; Hegde, A.S.; Arivazhagan, A.; Chandramouli, B.A.; Santosh, V.; Somasundaram, K. Proteomic identification of haptoglobin α2 as a glioblastoma serum biomarker: Implications in cancer cell migration and tumor growth. J. Proteome Res 2010, 9, 5557–5567. [Google Scholar]
- Albrethsen, J.; Bogebo, R.; Moller, C.H.; Olsen, J.A.; Raskov, H.H.; Gammeltoft, S. Candidate biomarker verification: Critical examination of a serum protein pattern for human colorectal cancer. Proteom. Clin. Appl 2012, 6, 182–189. [Google Scholar]
- Payne, P.R.; Huang, K.; Keen-Circle, K.; Kundu, A.; Zhang, J.; Borlawsky, T.B. Multi-dimensional discovery of biomarker and phenotype complexes. BMC Bioinforma 2010, 11, S3. [Google Scholar]
- Zhang, Y.; Li, Y.; Qiu, F.; Qiu, Z. Comparative analysis of the human urinary proteome by 1D SDS-PAGE and chip-HPLC-MS/MS identification of the AACT putative urinary biomarker. J. Chromatogr. B 2010, 878, 3395–3401. [Google Scholar]
- McDonald, W.H.; Yates, J.R., 3rd. Shotgun proteomics and biomarker discovery. Dis. Markers 2002, 18, 99–105. [Google Scholar]
- Jou, Y.J.; Lin, C.D.; Lai, C.H.; Chen, C.H.; Kao, J.Y.; Chen, S.Y.; Tsai, M.H.; Huang, S.H.; Lin, C.W. Proteomic identification of salivary transferrin as a biomarker for early detection of oral cancer. Anal. Chim. Acta 2010, 681, 41–48. [Google Scholar]
- Chu, W.; Ghahramani, Z.; Falciani, F.; Wild, D.L. Biomarker discovery in microarray gene expression data with Gaussian processes. Bioinformatics 2005, 21, 3385–3393. [Google Scholar]
- Lee, H.J.; Wark, A.W.; Corn, R.M. Microarray methods for protein biomarker detection. Analyst 2008, 133, 975–983. [Google Scholar]
- Liu, Y.; Wang, H.; Huang, J.; Yang, J.; Liu, B.; Yang, P. Microchip-based ELISA strategy for the detection of low-level disease biomarker in serum. Anal. Chim. Acta 2009, 650, 77–82. [Google Scholar]
- Zangar, R.C.; Daly, D.S.; White, A.M. ELISA microarray technology as a high-throughput system for cancer biomarker validation. Expert Rev. Proteomics 2006, 3, 37–44. [Google Scholar]
- Devonshire, A.S.; Sanders, R.; Wilkes, T.M.; Taylor, M.S.; Foy, C.A.; Huggett, J.F. Application of next generation qPCR and sequencing platforms to mRNA biomarker analysis. Methods 2013, 59, 89–100. [Google Scholar]
- Kaur, P.; Rizk, N.M.; Ibrahim, S.; Younes, N.; Uppal, A.; Dennis, K.; Karve, T.; Blakeslee, K.; Kwagyan, J.; Zirie, M.; et al. iTRAQ-based quantitative protein expression profiling and MRM verification of markers in type 2 diabetes. J. Proteome Res 2012, 11, 5527–5539. [Google Scholar]
- Ahn, Y.H.; Shin, P.M.; Oh, N.R.; Park, G.W.; Kim, H.; Yoo, J.S. A lectin-coupled, targeted proteomic mass spectrometry (MRM MS) platform for identification of multiple liver cancer biomarkers in human plasma. J. Proteomics 2012, 75, 5507–5515. [Google Scholar]
- Ji, C.; Tschantz, W.R.; Pfeifer, N.D.; Ullah, M.; Sadagopan, N. Development of a multiplex UPLC-MRM MS method for quantification of human membrane transport proteins OATP1B1, OATP1B3 and OATP2B1 in in vitro systems and tissues. Anal. Chim. Acta 2012, 717, 67–76. [Google Scholar]
- Weerapana, E.; Simon, G.M.; Cravatt, B.F. Disparate proteome reactivity profiles of carbon electrophiles. Nat. Chem. Biol 2008, 4, 405–407. [Google Scholar]
- Simon, G.M.; Cravatt, B.F. Activity-based proteomics of enzyme superfamilies: Serine hydrolases as a case study. J. Biol. Chem 2010, 285, 11051–11055. [Google Scholar]
- Sohn, C.H.; Agnew, H.D.; Lee, J.E.; Sweredoski, M.J.; Graham, R.L.; Smith, G.T.; Hess, S.; Czerwieniec, G.; Loo, J.A.; Heath, J.R.; et al. Designer reagents for mass spectrometry-based proteomics: Clickable cross-linkers for elucidation of protein structures and interactions. Anal. Chem 2012, 84, 2662–2669. [Google Scholar]
- Cheng, P.C.; Chang, H.K.; Chen, S.H. Quantitative nanoproteomics for protein complexes (QNanoPX) related to estrogen transcriptional action. Mol. Cell Proteomics 2010, 9, 209–224. [Google Scholar]
- Tian, D.; Zhang, H.; Chai, Y.; Cui, H. Synthesis of N-(aminobutyl)-N-(ethylisoluminol) functionalized gold nanomaterials for chemiluminescent bio-probe. Chem. Commun 2011, 47, 4959–4961. [Google Scholar]
- Koch, H.M.; Preuss, R.; Angerer, J. Di(2-ethylhexyl)phthalate (DEHP): Human metabolism and internal exposure—An update and latest results. Int. J. Androl 2006, 29, 155–165. [Google Scholar]
- Berggard, T.; Linse, S.; James, P. Methods for the detection and analysis of protein-protein interactions. Proteomics 2007, 7, 2833–2842. [Google Scholar]
- Rong, Y.; Chen, H.Z.; Wu, G.; Wang, M. Preparation and characterization of titanium dioxide nanoparticle/polystyrene composites via radical polymerization. Mater. Chem. Phys 2005, 91, 370–374. [Google Scholar]
- Matrix Science Inc. 2013. Available online: http://www.matrixscience.com (on accessed 28 September 2012).
- Ingenuity Systems, Inc. 2013. Available online: http://www.ingenuity.com (on accessed 6 February 2013).
- Richert, L.; Boulmedais, F.; Lavalle, P.; Mutterer, J.; Ferreux, E.; Decher, G.; Schaaf, P.; Voegel, J.C.; Picart, C. Improvement of stability and cell adhesion properties of polyelectrolyte multilayer films by chemical cross-linking. Biomacromolecules 2004, 5, 284–294. [Google Scholar]
- Hsu, J.L.; Huang, S.Y.; Chow, N.H.; Chen, S.H. Stable-isotope dimethyl labeling for quantitative proteomics. Anal. Chem 2003, 75, 6843–6852. [Google Scholar]
- Liu, P.S.; Chen, Y.Y. Butyl benzyl phthalate blocks Ca2+ signaling coupled with purinoceptor in rat PC12 cells. Toxicol. Appl. Pharmacol 2006, 210, 136–141. [Google Scholar]
- Liu, F.T. Regulatory roles of galectins in the immune response. Int. Arch. Allergy Immunol 2005, 136, 385–400. [Google Scholar]
- Universal Protein Resource (UniProt). 2013. Available online: http://www.uniprot.org/uniprot (on accessed 21 September 2011).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UNIPROT accession No. a | Protein identification | No. of peptides b | Molecular mass (kDa) b | H/L(I) c | H/L(II) c | H/L(III) c | Average | Std |
|---|---|---|---|---|---|---|---|---|
| Score > 90 and at least the ratio (D/H) > 3 of two experiments | ||||||||
| P85972 | Vinculin | 2 | 119.5 | 9.8 | 5.0 | 4.4 | 6.4 | 3.0 |
| P62630 | Elongation factor 1-alpha 1 | 13 | 51.9 | 10.0 | 4.1 | 4.6 | 6.3 | 3.3 |
| P04636 | Malate dehydrogenase | 3 | 36.9 | 5.3 | 5.9 | 6.0 | 5.7 | 0.4 |
| P11980 | Pyruvate kinase isozymes M1/M2 | 4 | 59.4 | 4.9 | 8.5 | 3.0 | 5.5 | 2.8 |
| P07150 | Annexin A1 | 7 | 40.1 | 7.3 | 3.6 | 4.8 | 5.2 | 1.9 |
| P04642 | l-lactate dehydrogenase A chain | 3 | 37.6 | 5.1 | 3.3 | 5.5 | 4.7 | 1.2 |
| Q6P9V9 | Tubulin alpha-1B chain | 11 | 51.4 | 6.1 | 3.5 | 4.3 | 4.6 | 1.3 |
| Q63081 | Protein disulfide-isomerase A6 | 5 | 49.6 | 7.2 | 3.7 | 2.4 | 4.4 | 2.5 |
| P04764 | Alpha-enolase | 13 | 48.6 | 5.4 | 2.9 | 4.1 | 4.1 | 1.3 |
| P29457 | Serpin H1 | 12 | 47.7 | 5.6 | 3.6 | 3.1 | 4.1 | 1.3 |
| Score > 80, and at least the ratio (D/H) > 3 of one experiment | ||||||||
| P15999 | ATP synthase subunit alpha | 3 | 60.8 | 17.9 | 1.9 | 1.5 | ||
| P34058 | Heat shock protein HSP 90-beta | 3 | 85.9 | 13.3 | 2.9 | 3.5 | ||
| P69897 | Tubulin beta-5 chain | 2 | 50.5 | 12.8 | 4.3 | n.a.d | ||
| P04797 | Glyceraldehyde-3-phosphate dehydrogenase | 3 | 36.9 | 12.0 | 5.8 | n.a. | ||
| P62963 | Profilin-1 | 3 | 15.4 | 7.5 | 4.0 | 2.2 | ||
| Q5FVM4 | Non-POU domain-containing octamer-binding protein | 2 | 55.7 | 6.7 | 0.5 | 1.2 | ||
| P63018 | Heat shock cognate 71 kDa protein | 4 | 72.7 | 5.1 | 3.5 | 5.7 | ||
| P04785 | Protein disulfide-isomerase | 5 | 58.9 | 5.0 | 1.7 | 1.8 | ||
| P06761 | 78 kDa glucose-regulated protein | 5 | 74.4 | 4.9 | 2.9 | 2.1 | ||
| Q66HD0 | Endoplasmin | 4 | 95.3 | 4.7 | 3.6 | n.a. | ||
| B0BN93 | 26S proteasome non-ATPase regulatory subunit 13 | 5 | 43.8 | 3.6 | 6.9 | 3.5 | ||
| P10719 | ATP synthase subunit beta | 6 | 57.0 | 3.2 | 1.1 | 2.1 | ||
| P50399 | Rab GDP dissociation inhibitor beta | 3 | 52.2 | 3.2 | 4.1 | n.a. | ||
| Score < 80 and at least the ratio (D/H) > 10 of one experiment | ||||||||
| P48500 | Triosephosphate isomerase | 2 | 28.0 | n.a. | 27.2 | n.a. | ||
| Q5RKI1 | Eukaryotic initiation factor 4A-II | 2 | 47.3 | n.a. | 14.7 | n.a. | ||
| P08699 | Galectin-3 | 2 | 27.4 | n.a. | 12.0 | n.a. | ||
| P11762 | Galectin-1 | 2 | 15.4 | 39.9 | 9.2 | 14.0 | ||
| Q923J6 | Dynein heavy chain 12 | 2 | 366.7 | 27.6 | n.a. | n.a. | ||
| P63326 | 40S ribosomal protein S10 | 2 | 19.3 | 14.2 | 11.3 | 7.6 | ||
| P05964 | Protein S100-A6 | 2 | 10.4 | 11.4 | n.a. | 5.8 | ||
| P16617 | Phosphoglycerate kinase 1 | 2 | 46.2 | 10.8 | 6.9 | 5.6 | ||
| P11884 | Aldehyde dehydrogenase | 2 | 57.9 | n.a. | n.a. | 13.1 | ||
| P16638 | ATP-citrate synthase | 2 | 123.6 | n.a. | 13.9 | n.a. | ||
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liang, S.-S.; Liao, W.-T.; Kuo, C.-J.; Chou, C.-H.; Wu, C.-J.; Wang, H.-M. Phthalic Acid Chemical Probes Synthesized for Protein-Protein Interaction Analysis. Int. J. Mol. Sci. 2013, 14, 12914-12930. https://doi.org/10.3390/ijms140712914
Liang S-S, Liao W-T, Kuo C-J, Chou C-H, Wu C-J, Wang H-M. Phthalic Acid Chemical Probes Synthesized for Protein-Protein Interaction Analysis. International Journal of Molecular Sciences. 2013; 14(7):12914-12930. https://doi.org/10.3390/ijms140712914
Chicago/Turabian StyleLiang, Shih-Shin, Wei-Ting Liao, Chao-Jen Kuo, Chi-Hsien Chou, Chin-Jen Wu, and Hui-Min Wang. 2013. "Phthalic Acid Chemical Probes Synthesized for Protein-Protein Interaction Analysis" International Journal of Molecular Sciences 14, no. 7: 12914-12930. https://doi.org/10.3390/ijms140712914