miR-125b Regulates the Early Steps of ESC Differentiation through Dies1 in a TGF-Independent Manner


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
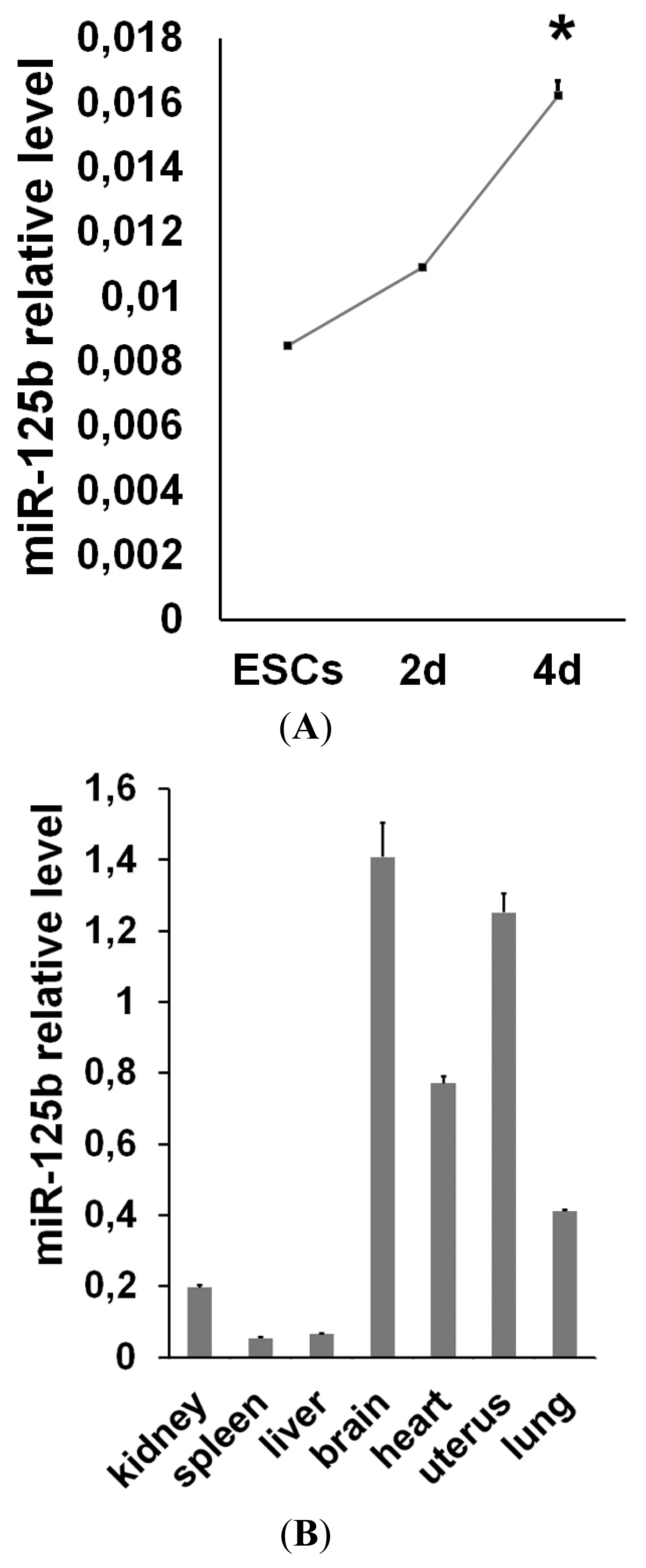
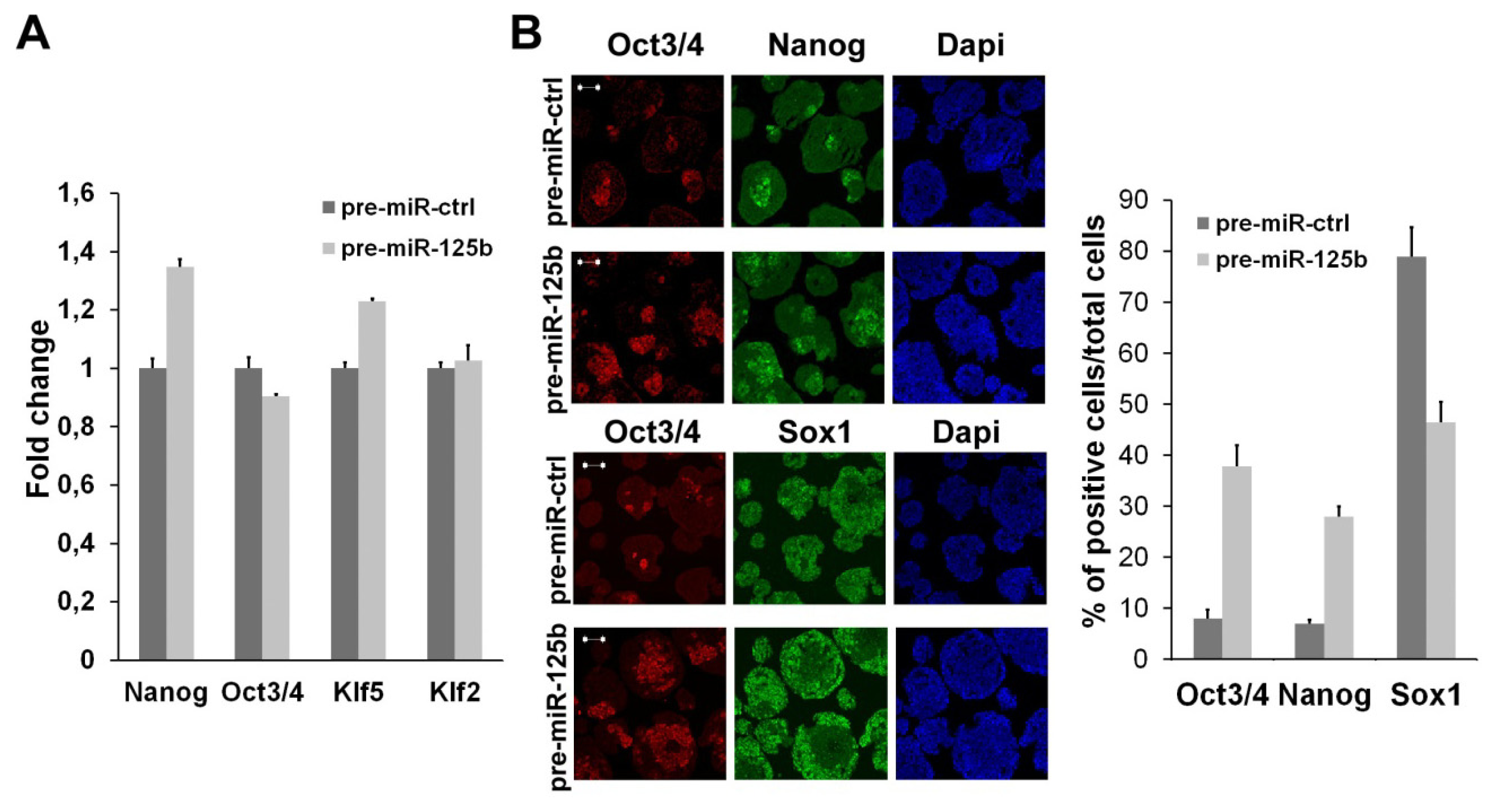
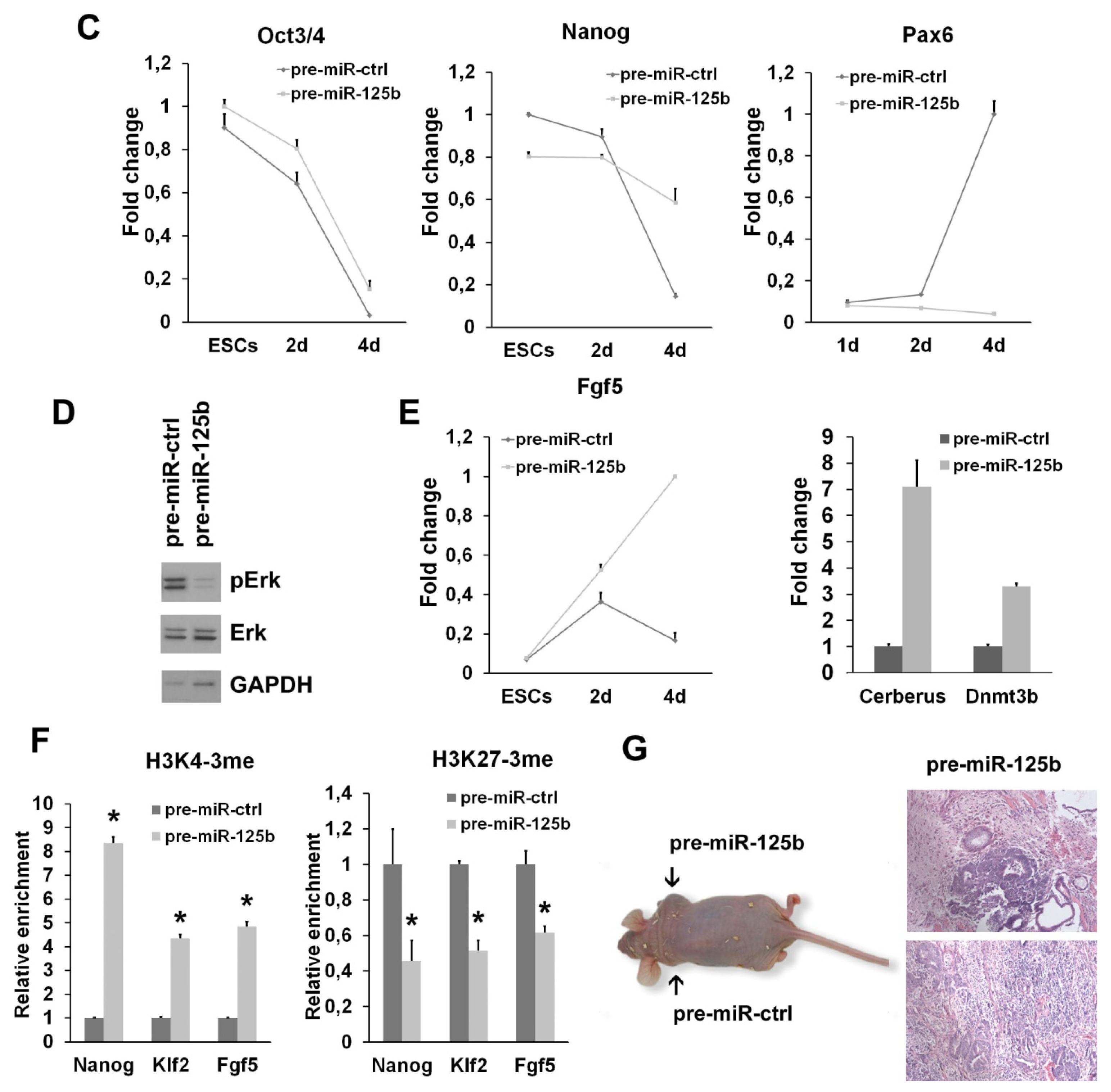
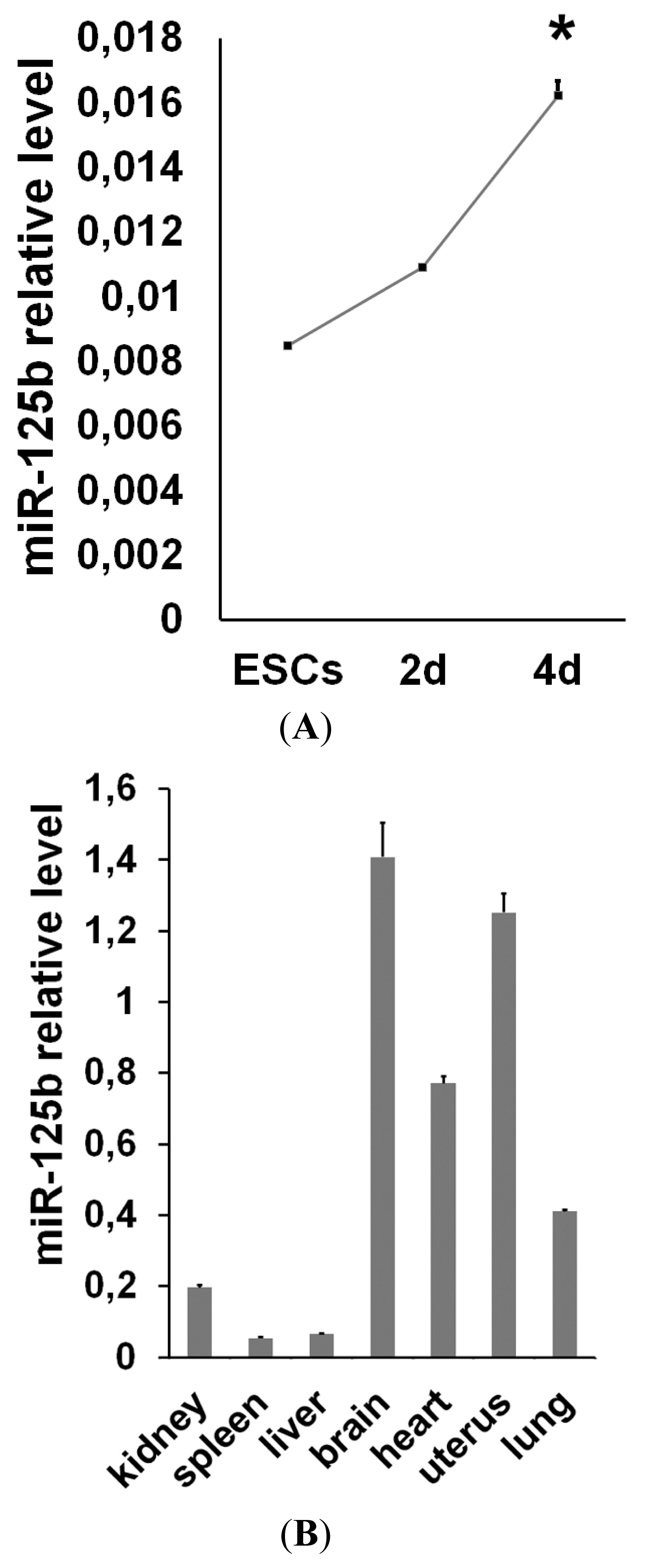
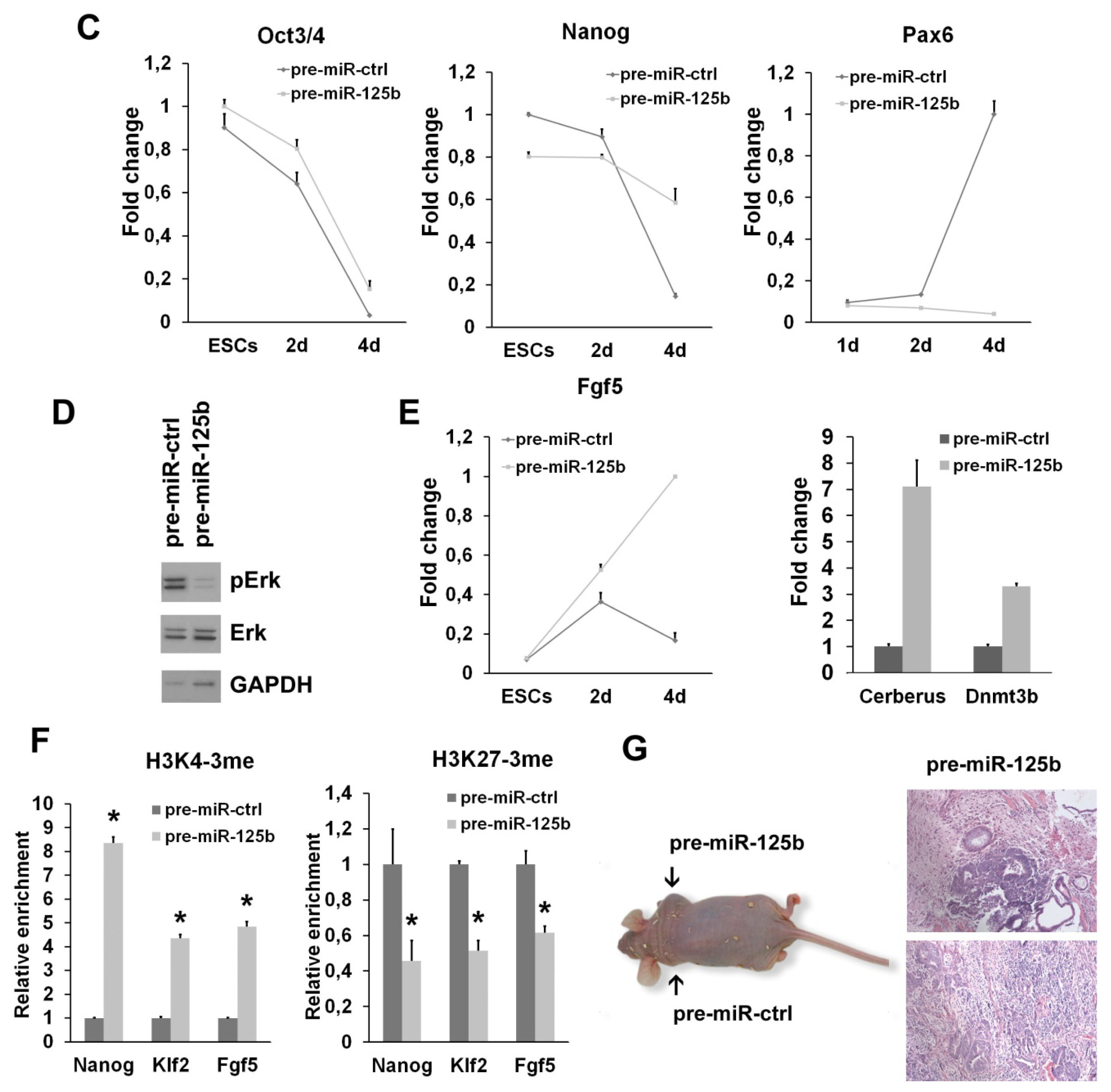
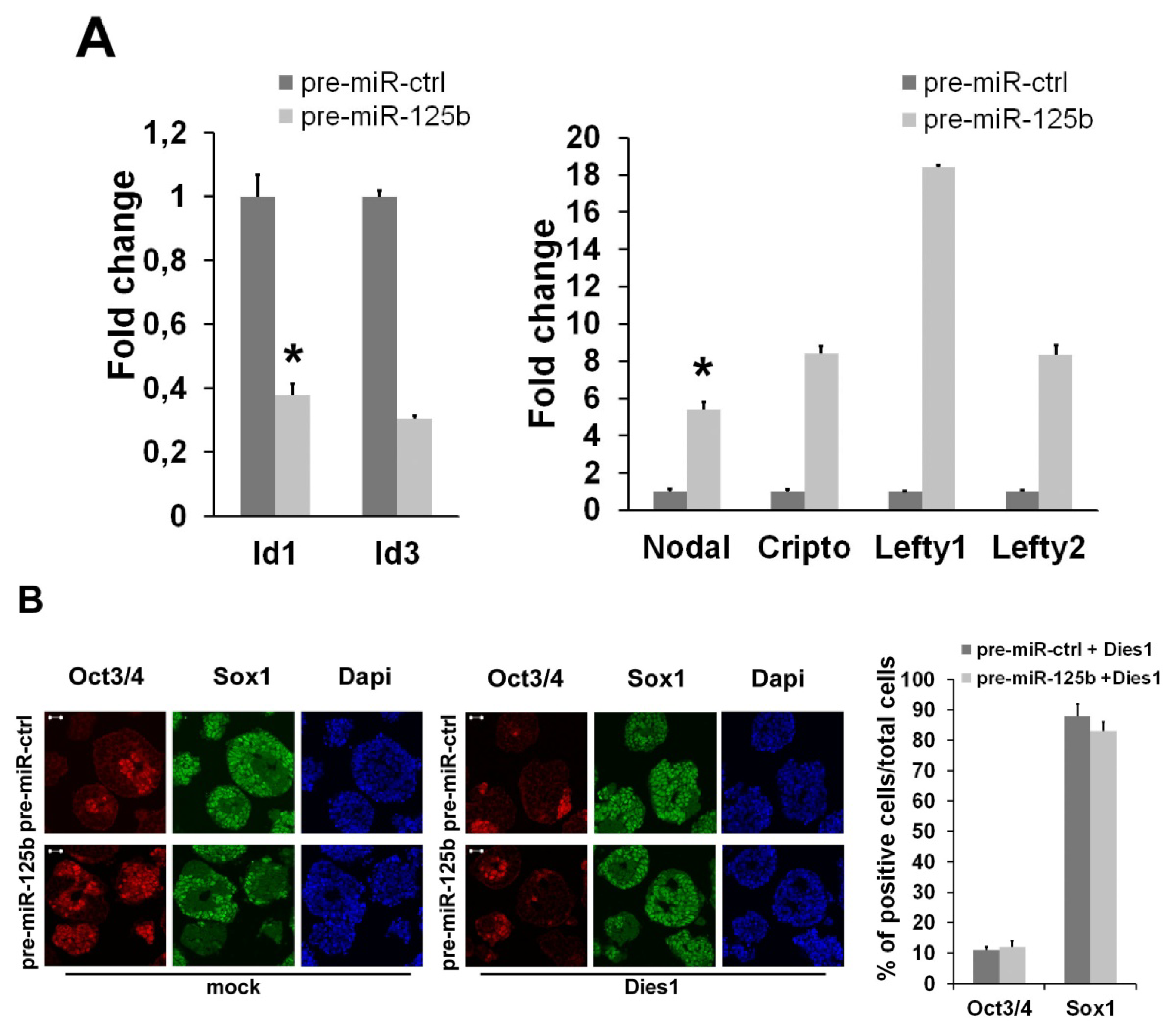
2.1. miR-125b Overexpression Blocks ESC at the Epiblast Stage
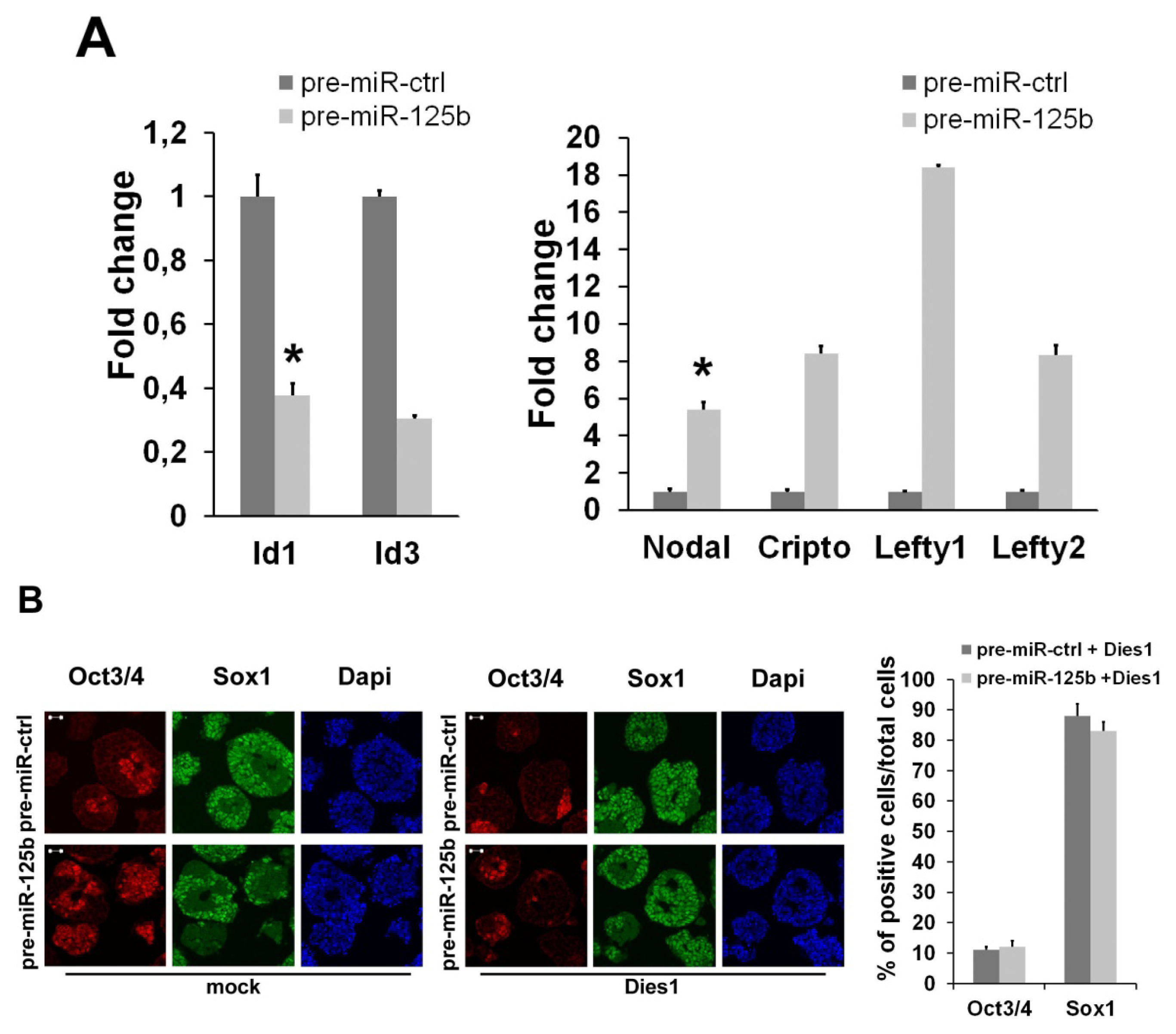
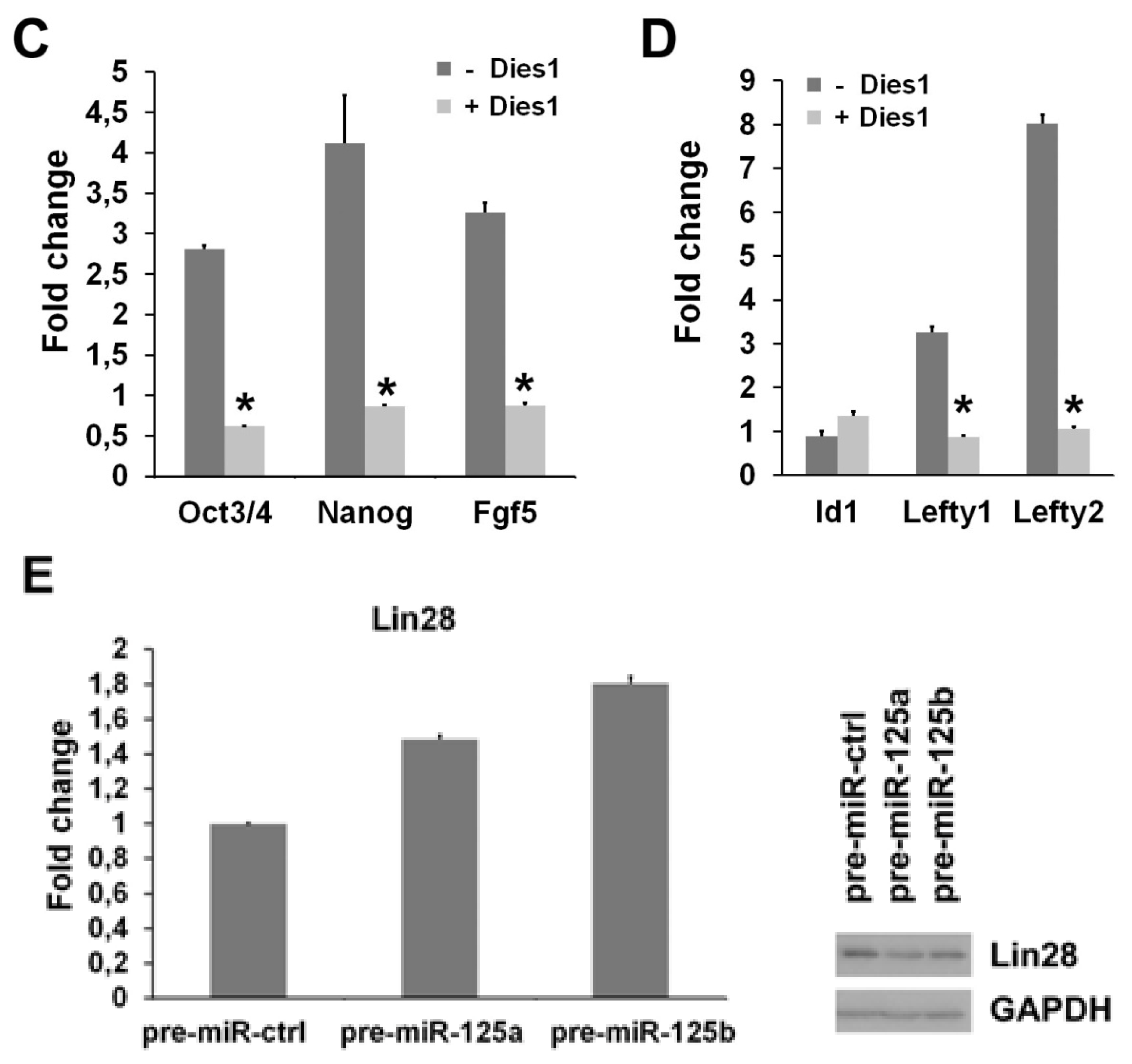
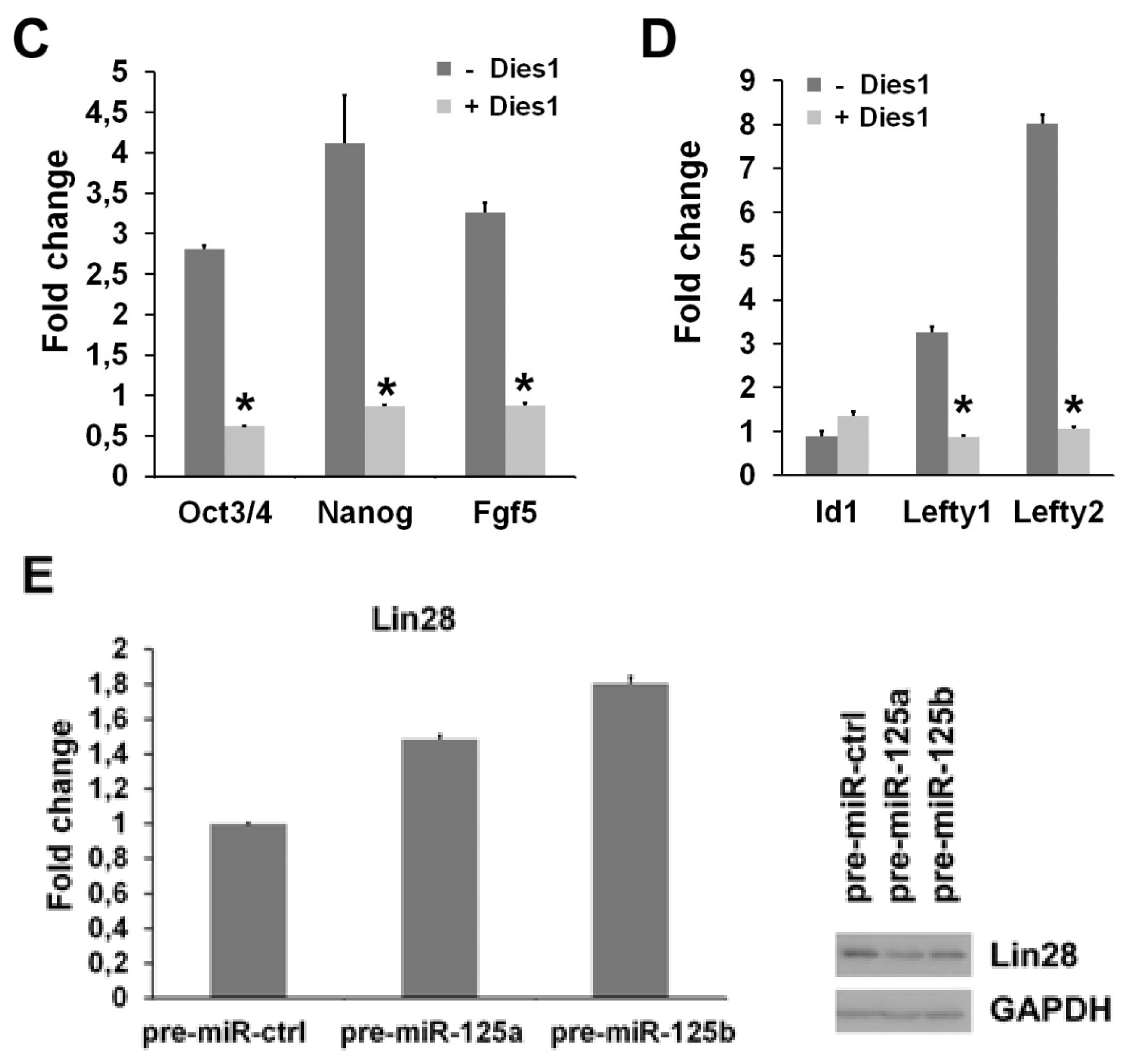
2.2. miR-125b Effects on the ESC-Epiblast Transition Are Due to Dies1
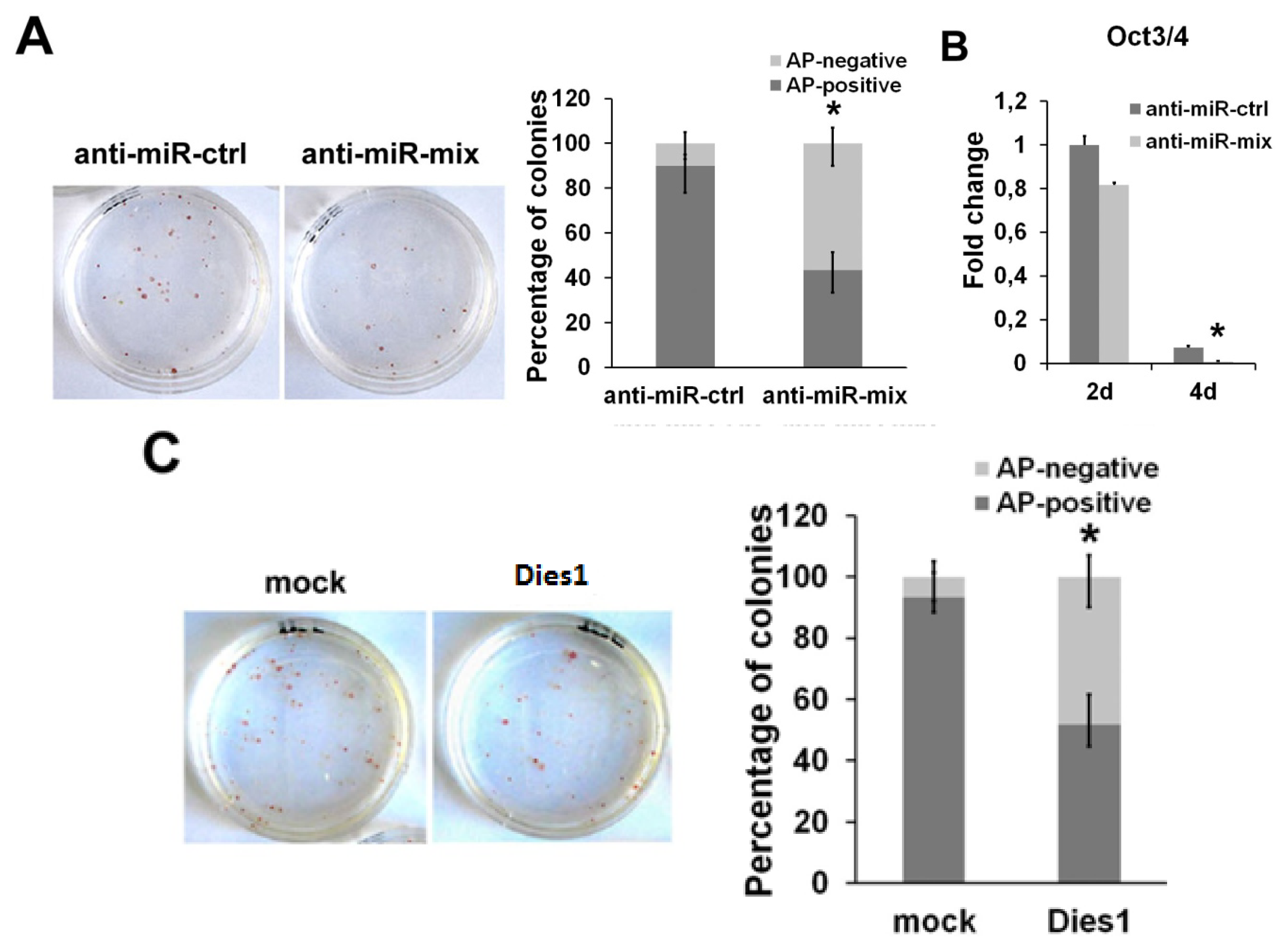
2.3. miR-125 Suppression Promotes ESC Differentiation
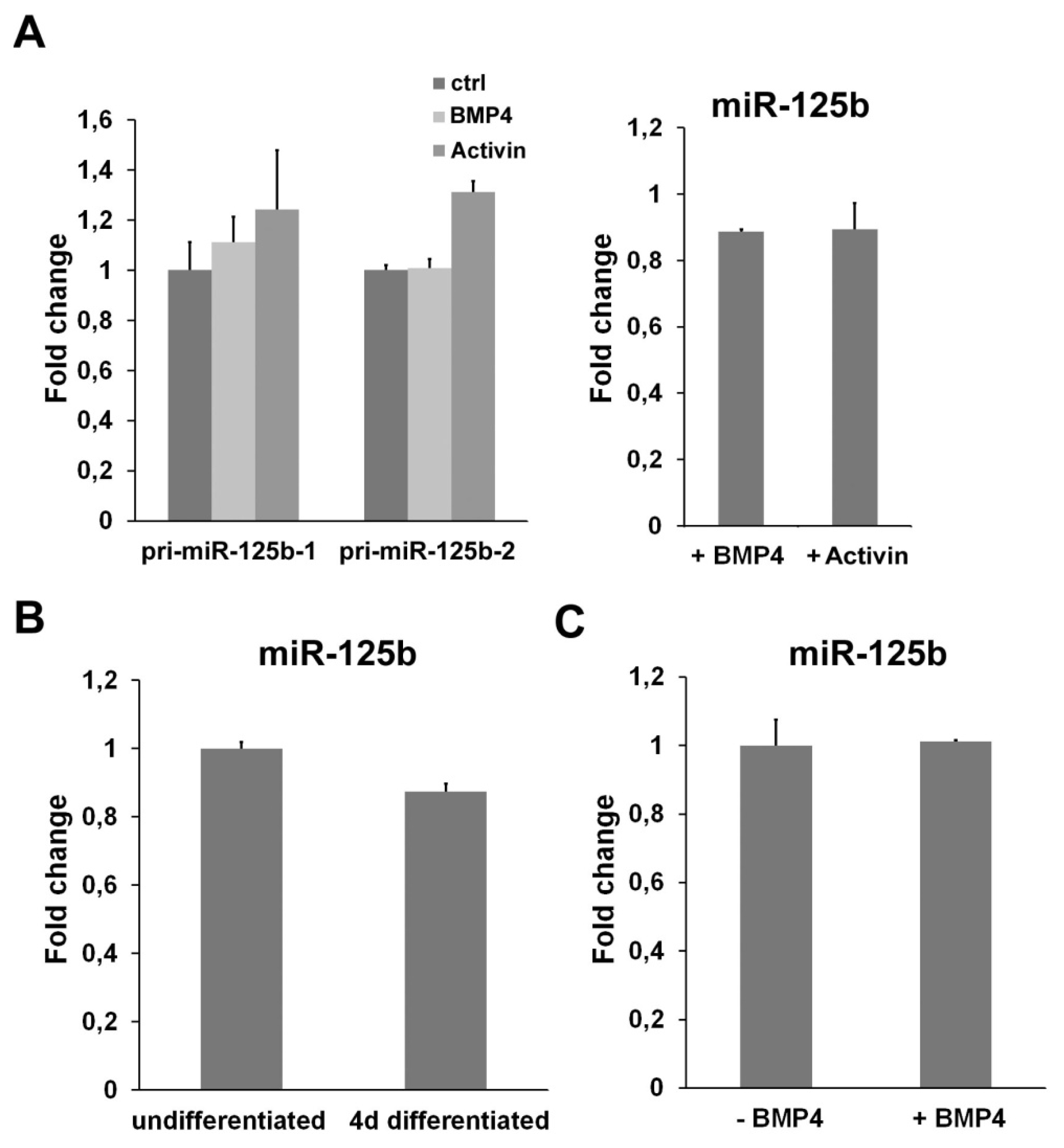
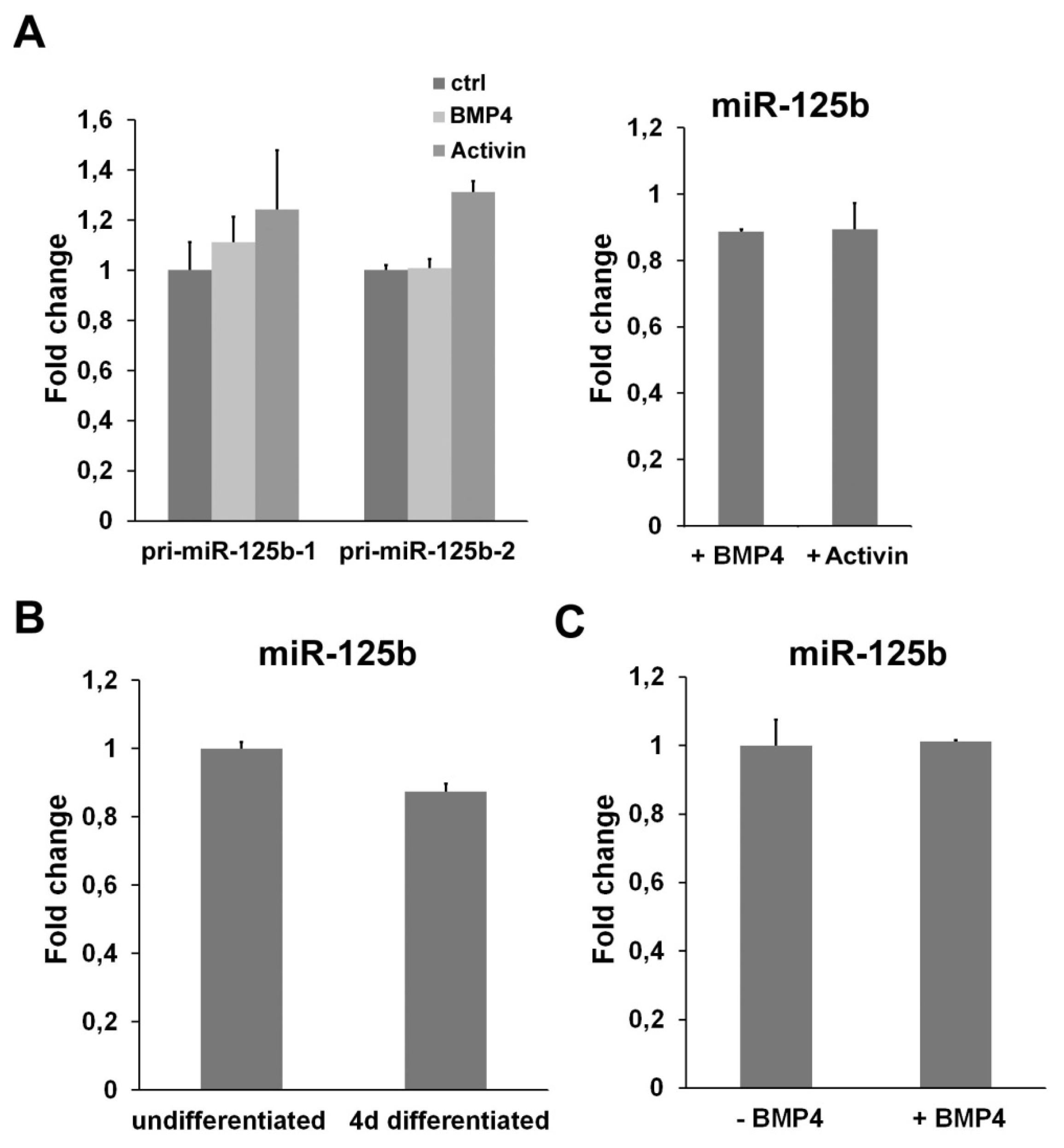
2.4. miR-125b Is Not Regulated by a TGFβ Signaling
3. Experimental Section
3.1. Cell Culture, Transfection and Differentiation
3.2. Cell Treatment and Alkaline Phosphatase Staining
3.3. RNA Isolation, q-PCR and TaqMan Analysis
3.4. Northern Blot Analysis
3.5. Protein Extraction and Western Blot Analysis
3.6. Immunostaining
3.7. Teratoma Formation
3.8. Chromatin Immunoprecipitation (ChIP)-qPCR Analysis
3.9. Statistics
4. Conclusions
Supplementary Information
ijms-14-13482-s001.pdfAcknowledgments
Conflict of Interest
References
- Niwa, H.; Ogawa, K.; Shimosato, D.; Adachi, K. A parallel circuit of LIF signalling pathways maintains pluripotency of mouse ES cells. Nature 2009, 460, 118–122. [Google Scholar]
- Parisi, S.; D’Andrea, D.; Lago, C.T.; Adamson, E.D.; Persico, M.G.; Minchiotti, G. Nodal-dependent Cripto signaling promotes cardiomyogenesis and redirects the neural fate of embryonic stem cells. J. Cell Biol 2003, 163, 303–314. [Google Scholar]
- Seuntjens, E.; Umans, L.; Zwijsen, A.; Sampaolesi, M.; Verfaillie, C.M.; Huylebroeck, D. Transforming growth factor type beta and Smad family signaling in stem cell function. Cytokine Growth Factor Rev 2009, 20, 449–458. [Google Scholar]
- Watabe, T.; Miyazono, K. Roles of TGF-beta family signaling in stem cell renewal and differentiation. Cell Res 2009, 19, 103–115. [Google Scholar]
- Zhang, K.; Li, L.; Huang, C.; Shen, C.; Tan, F.; Xia, C.; Liu, P.; Rossant, J.; Jing, N. Distinct functions of BMP4 during different stages of mouse ES cell neural commitment. Development 2010, 137, 2095–2105. [Google Scholar]
- Lee, K.L.; Lim, S.K.; Orlov, Y.L.; Yit le, Y.; Yang, H.; Ang, L.T.; Poellinger, L.; Lim, B. Graded Nodal/Activin signaling titrates conversion of quantitative phospho-Smad2 levels into qualitative embryonic stem cell fate decisions. PLoS Genet. 2011. [Google Scholar] [CrossRef]
- Galvin, K.E.; Travis, E.D.; Yee, D.; Magnuson, T.; Vivian, J.L. Nodal signaling regulates the bone morphogenic protein pluripotency pathway in mouse embryonic stem cells. J. Biol. Chem 2010, 285, 19747–19756. [Google Scholar]
- Parisi, S.; Battista, M.; Musto, A.; Navarra, A.; Tarantino, C.; Russo, T. A regulatory loop involving Dies1 and miR-125a controls BMP4 signaling in mouse embryonic stem cells. FASEB J 2012, 26, 3957–3968. [Google Scholar]
- Chambers, I.; Tomlinson, S.R. The transcriptional foundation of pluripotency. Development 2009, 136, 2311–2322. [Google Scholar]
- Parisi, S.; Russo, T. Regulatory role of Klf5 in early mouse development and in embryonic stem cells. Vitam Horm 2011, 87, 381–397. [Google Scholar]
- Ivey, K.N.; Srivastava, D. MicroRNAs as regulators of differentiation and cell fate decisions. Cell Stem Cell 2010, 7, 36–41. [Google Scholar]
- Martinez, N.J.; Gregory, R.I. MicroRNA gene regulatory pathways in the establishment and maintenance of ESC identity. Cell Stem Cell 2010, 7, 31–35. [Google Scholar]
- Houbaviy, H.B.; Murray, M.F.; Sharp, P.A. Embryonic stem cell specific microRNAs. Dev. Cell 2003, 5, 351–358. [Google Scholar]
- Viswanathan, S.R.; Daley, G.Q.; Gregory, R.I. Selective blockade of microRNA processing by Lin28. Science 2008, 320, 97–100. [Google Scholar]
- Sinkkonen, L.; Hugenschmidt, T.; Berninger, P.; Gaidatzis, D.; Mohn, F.; Artus-Revel, C.G.; Zavolan, M.; Svoboda, P.; Filipowicz, W. MicroRNAs control de novo DNA methylation through regulation of transcriptional repressors in mouse embryonic stem cells. Nat. Struct. Mol. Biol 2008, 15, 259–267. [Google Scholar]
- Zovoilis, A.; Smorag, L.; Pantazi, A.; Engel, W. Members of the miR-290 cluster modulate in vitro differentiation of mouse embryonic stem cells. Differentiation 2009, 78, 69–78. [Google Scholar]
- Wang, Y.; Baskerville, S.; Shenoy, A.; Babiarz, J.E.; Baehner, L.; Blelloch, R. Embryonic stem cell-specific microRNAs regulate the G1-S transition and promote rapid proliferation. Nat. Genet 2008, 40, 1478–1483. [Google Scholar]
- Tay, Y.; Zhang, J.; Thomson, A.M.; Lim, B.; Rigoutsos, I. MicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiation. Nature 2008, 455, 1124–1128. [Google Scholar]
- Juan, A.H.; Kumar, R.M.; Marx, J.G.; Young, R.A.; Sartorelli, V. Mir-214-dependent regulation of the polycomb protein Ezh2 in skeletal muscle and embryonic stem cells. Mol. Cell 2009, 36, 61–74. [Google Scholar]
- Delaloy, C.; Liu, L.; Lee, J.A.; Su, H.; Shen, F.; Yang, G.Y.; Young, W.L.; Ivey, K.N.; Gao, F.B. MicroRNA-9 coordinates proliferation and migration of human embryonic stem cell-derived neural progenitors. Cell Stem Cell 2010, 6, 323–335. [Google Scholar]
- Krichevsky, A.M.; Sonntag, K.C.; Isacson, O.; Kosik, K.S. Specific microRNAs modulate embryonic stem cell-derived neurogenesis. Stem Cells 2006, 24, 857–864. [Google Scholar]
- Tarantino, C.; Paolella, G.; Cozzuto, L.; Minopoli, G.; Pastore, L.; Parisi, S.; Russo, T. miRNA-34a, 100, and 137 modulate differentiation of mouse embryonic stem cells. FASEB J 2010, 24, 3255–3263. [Google Scholar]
- Wang, J.; Cao, N.; Yuan, M.; Cui, H.; Tang, Y.; Qin, L.; Huang, X.; Shen, N.; Yang, H.T. MicroRNA-125b/Lin28 pathway contributes to the mesendodermal fate decision of embryonic stem cells. Stem Cells Dev 2012, 21, 1524–1537. [Google Scholar]
- Ge, Y.; Sun, Y.; Chen, J. IGF-II is regulated by microRNA-125b in skeletal myogenesis. J. Cell Biol 2011, 192, 69–81. [Google Scholar]
- Dahlqvist, C.; Blokzijl, A.; Chapman, G.; Falk, A.; Dannaeus, K.; Ibâñez, C.F.; Lendahl, U. Functional Notch signaling is required for BMP4-induced inhibition of myogenic differentiation. Development 2003, 130, 6089–6099. [Google Scholar]
- Sequence Detection Systems (SDA), version 1.4; software for detecting nucleic acid sequence. Available online: http://www.lifetechnologies.com/global/en/home/technical-resources/software-downloads/applied-biosystems-7500-fast-real-time-pcr-system.html (on accessed 31 January, 2008).
- LSM 510 Meta software. Available online: http://microscopy.zeiss.com/microscopy/n_us/downloads/lsm-5-series.html (on accessed 27 February 2007).
- Parisi, S.; Cozzuto, L.; Tarantino, C.; Passaro, F.; Ciriello, S.; Aloia, L.; Antonini, D.; de Simone, V.; Pastore, L.; Russo, T. Direct targets of Klf5 transcription factor contribute to the maintenance of mouse embryonic stem cell undifferentiated state. BMC Biol. 2010. [Google Scholar] [CrossRef]
- Aloia, L.; Parisi, S.; Fusco, L.; Pastore, L.; Russo, T. Differentiation of embryonic stem cells 1 (Dies1) is a component of bone morphogenetic protein 4 (BMP4) signaling pathway required for proper differentiation of mouse embryonic stem cell. J. Biol. Chem 2010, 285, 7776–7783. [Google Scholar]


© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Battista, M.; Musto, A.; Navarra, A.; Minopoli, G.; Russo, T.; Parisi, S. miR-125b Regulates the Early Steps of ESC Differentiation through Dies1 in a TGF-Independent Manner. Int. J. Mol. Sci. 2013, 14, 13482-13496. https://doi.org/10.3390/ijms140713482
Battista M, Musto A, Navarra A, Minopoli G, Russo T, Parisi S. miR-125b Regulates the Early Steps of ESC Differentiation through Dies1 in a TGF-Independent Manner. International Journal of Molecular Sciences. 2013; 14(7):13482-13496. https://doi.org/10.3390/ijms140713482
Chicago/Turabian StyleBattista, Marica, Anna Musto, Angelica Navarra, Giuseppina Minopoli, Tommaso Russo, and Silvia Parisi. 2013. "miR-125b Regulates the Early Steps of ESC Differentiation through Dies1 in a TGF-Independent Manner" International Journal of Molecular Sciences 14, no. 7: 13482-13496. https://doi.org/10.3390/ijms140713482