Genetic Diversity of the Critically Endangered Thuja sutchuenensis Revealed by ISSR Markers and the Implications for Conservation

Abstract
:1. Introduction
2. Results
2.1. Genetic Diversity
2.2. Genetic Differentiation
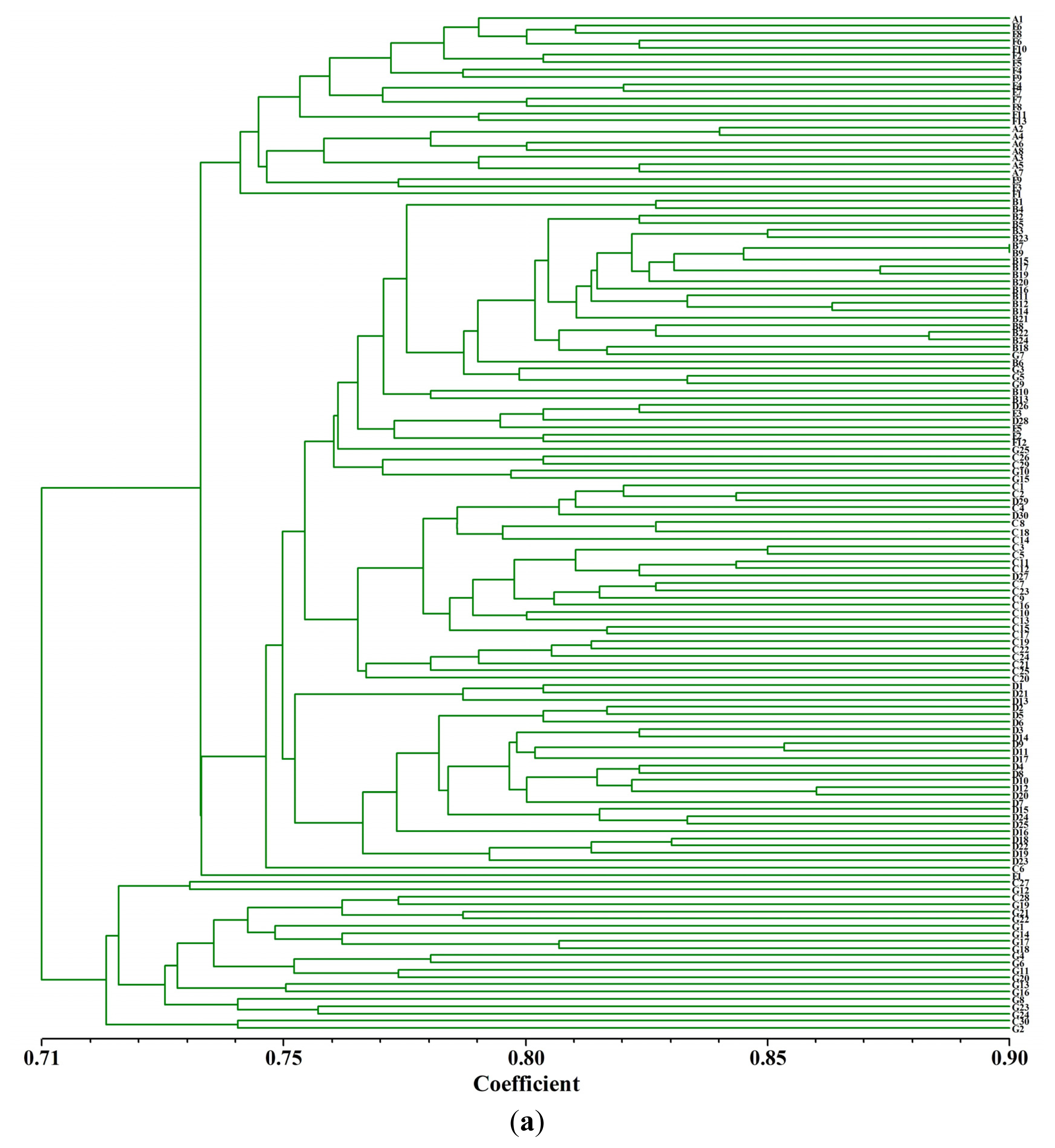
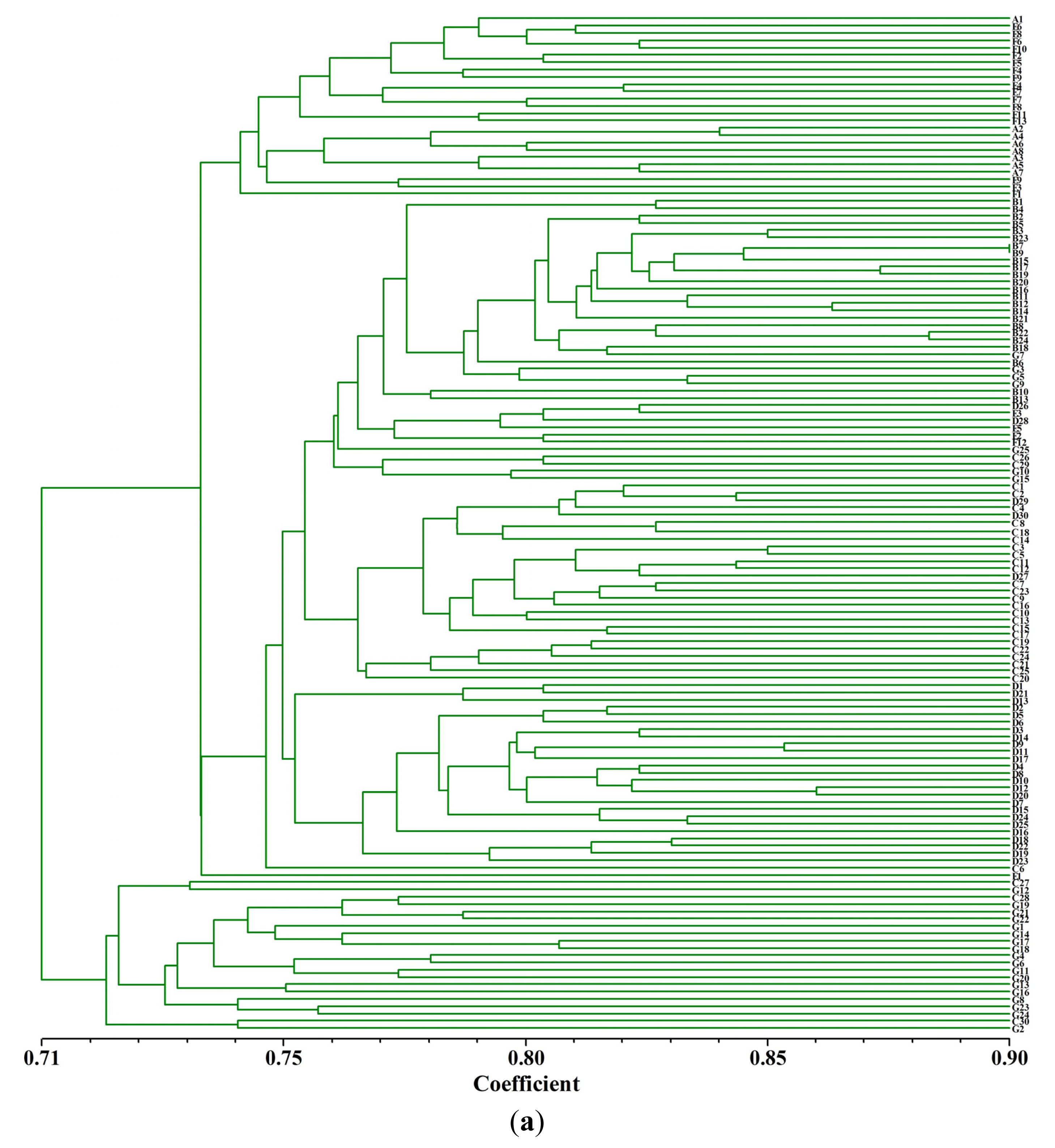
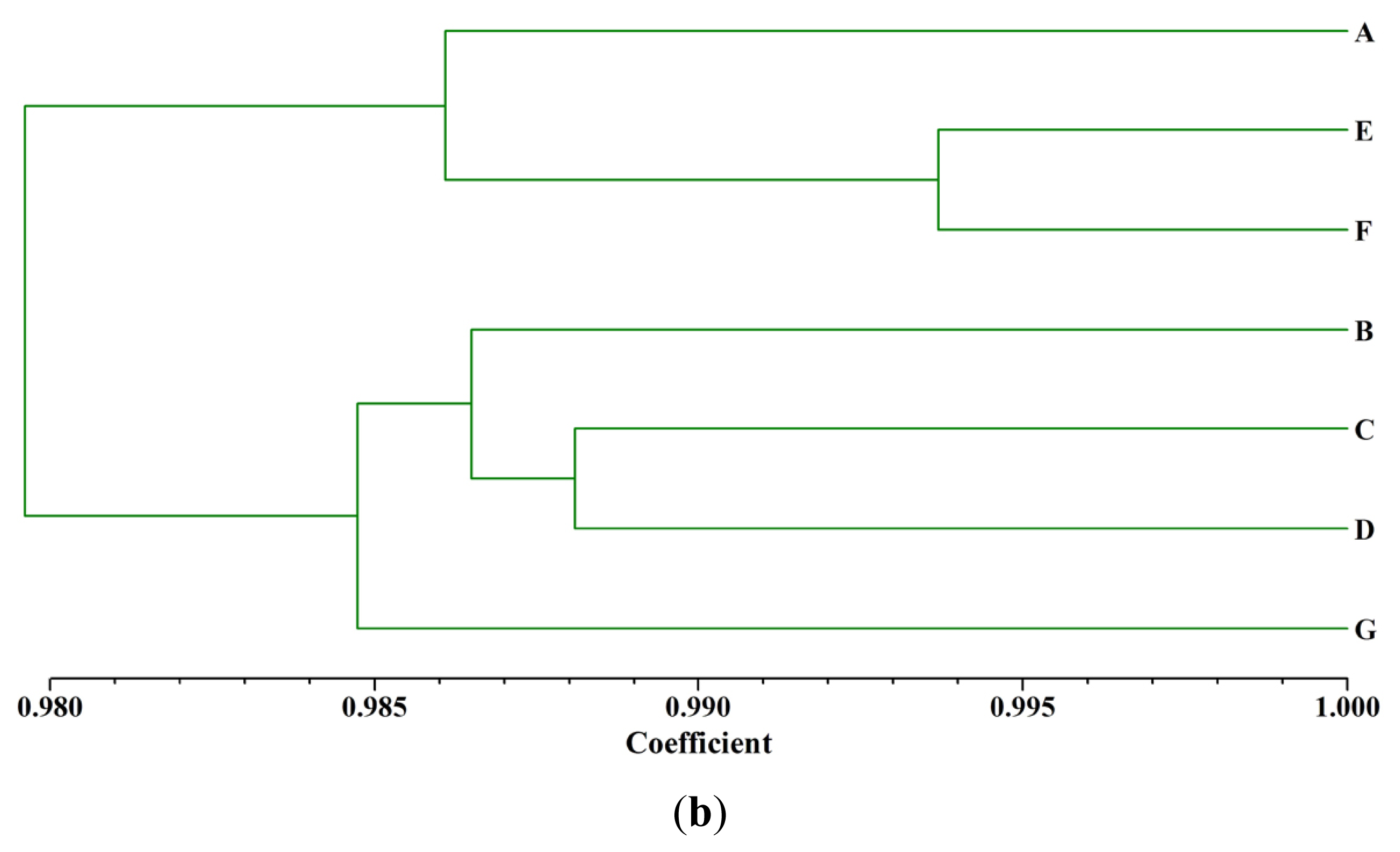
2.3. Cluster Analysis
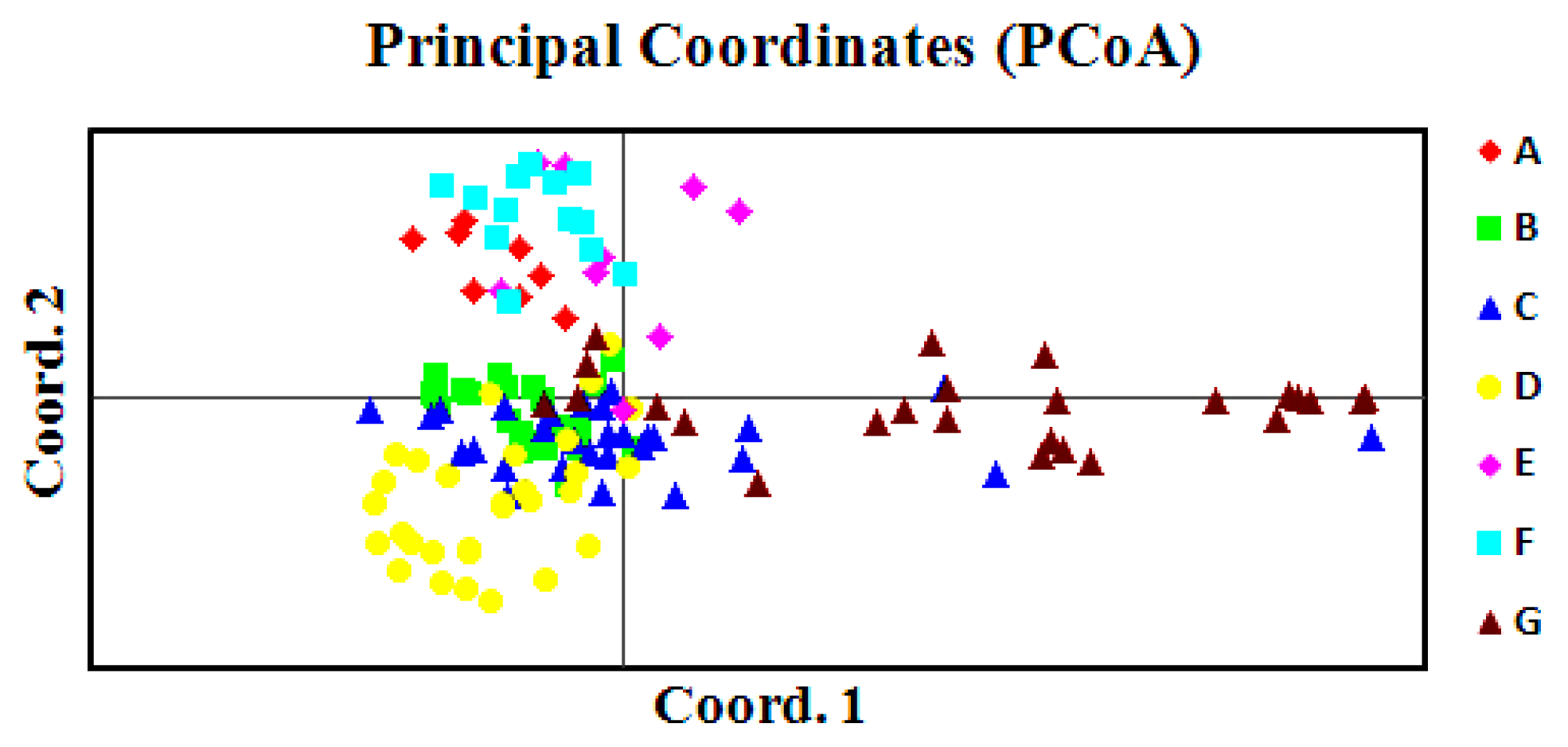
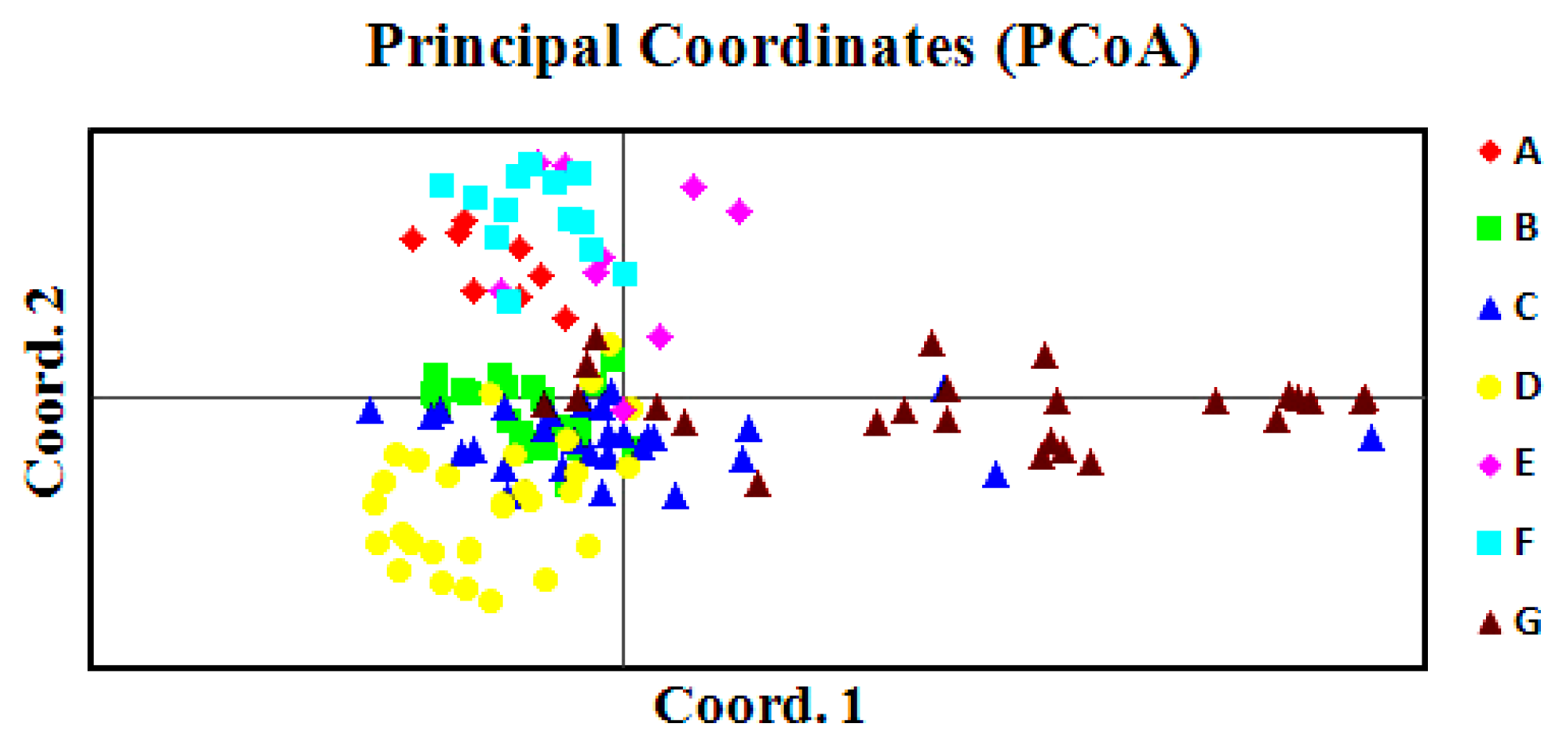
2.4. Principal Coordinate Analysis
3. Discussion
3.1. High Genetic Diversity
3.2. Low Genetic Divergence
3.3. Implication for Conservation
4. Experimental Section
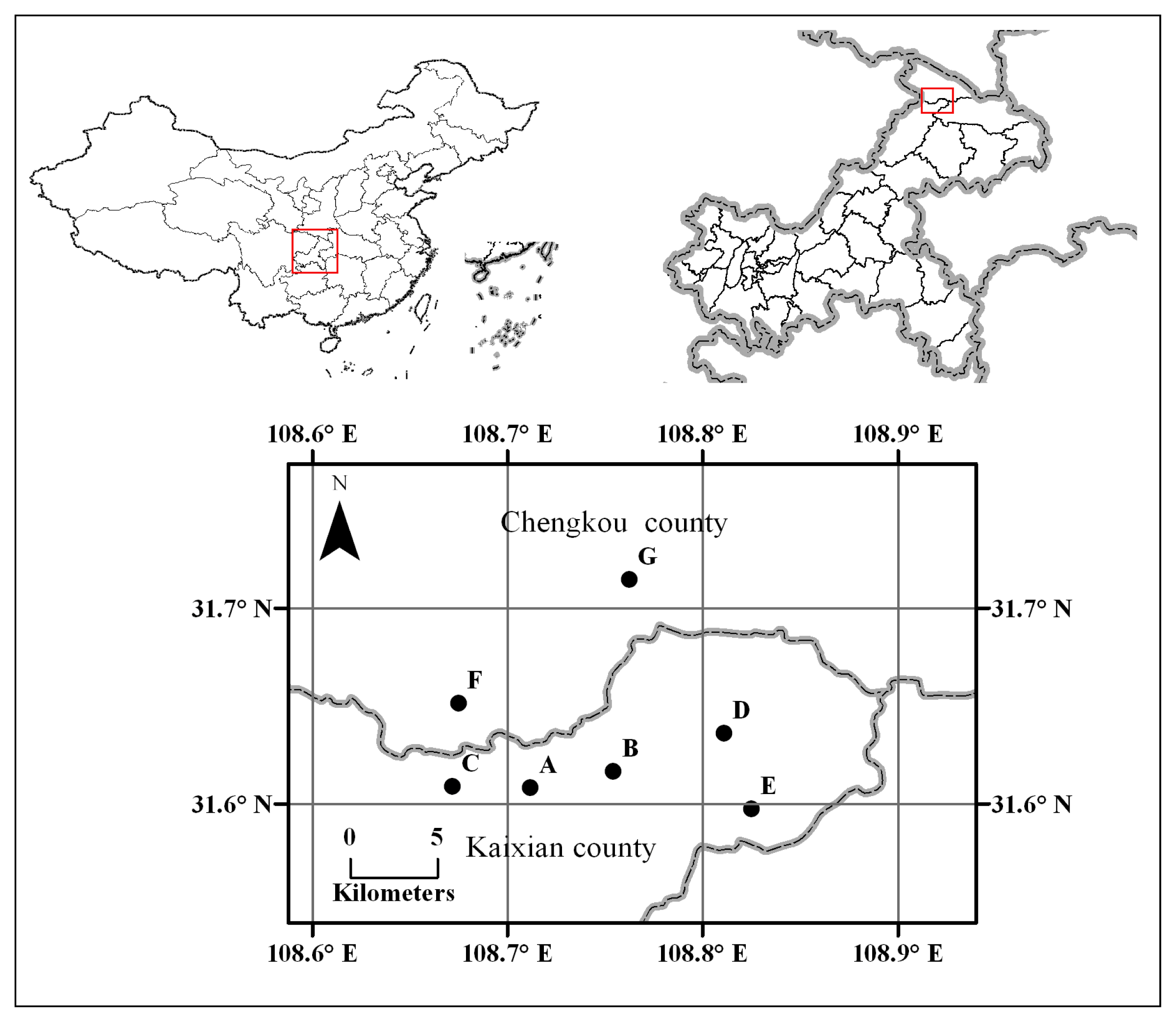
4.1. Study Species and Population Sampling
4.2. DNA Extraction
4.3. ISSR Amplification
4.4. Data Analysis
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Xiang, Q.; Fajon, A.; Li, Z.; Fu, L.; Liu, Z. Thuja Sutchuenensis. In IUCN Red List of Threatened Species, Version 2012.2; The International Union for Conservation of Nature (IUCN): Cambridge, UK, 2012. [Google Scholar]
- Xiang, Q.; Fajon, A.; Li, Z.; Fu, L.; Liu, Z. Thuja sutchuenensis: A rediscovered species of the Cupressaceae. Bot. J. Linn. Soc 2002, 139, 305–310. [Google Scholar]
- Ge, X.J.; Yu, Y.; Zhao, N.X.; Chen, H.S.; Qi, W.Q. Genetic variation in the endangered Inner Mongolia endemic shrub Tetraena mongolica Maxim. (Zygophyllaceae). Biol. Conserv 2003, 111, 427–434. [Google Scholar]
- Zhang, X.P.; Li, X.H.; Qiu, Y.X. Genetic diversity of the endangered species Kirengeshoma palmata (Saxifragaceae) in China. Biochem. Syst. Ecol 2006, 34, 38–47. [Google Scholar]
- Zheng, D.J.; Xie, L.S.; Zhu, J.H.; Zhang, Z.L. Low genetic diversity and local adaptive divergence of Dracaena cambodiana (Liliaceae) populations associated with historical population bottlenecks and natural selection: An endangered long-lived tree endemic to Hainan Island, China. Plant Biol 2012, 14, 828–838. [Google Scholar]
- Ding, G.; Zhang, D.; Yu, Y.; Zhao, L.; Zhang, B. Analysis of genetic variability and population structure of the endemic medicinal Limonium sinense using molecular markers. Gene 2013, 520, 189–193. [Google Scholar]
- Frankham, R.; Briscoe, D.A.; Ballou, J.D. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Culley, T.M.; Wolfe, A.D. Population genetic structure of the cleistogamous plant species Viola pubescens Aiton (Violaceae), as indicated by allozyme and ISSR molecular markers. Heredity 2001, 86, 545–556. [Google Scholar]
- Vicente, M.J.; Segura, F.; Aguado, M.; Migliaro, D.; Franco, J.A.; Martínez-Sánchez, J.J. Genetic diversity of Astragalus nitidiflorus, a critically endangered endemic of SE Spain, and implications for its conservation. Biochem. Syst. Ecol 2011, 39, 175–182. [Google Scholar]
- Rubio-Moraga, A.; Candel-Perez, D.; Lucas-Borja, M.E.; Tiscar, P.A.; Viñegla, B.; Linares, J.C.; Gómez-Gómez, L.; Ahrazem, O. Genetic diversity of Pinus nigra Arn. populations in southern Spain and northern Morocco revealed by inter-simple sequence repeat profiles. Int. J. Mol. Sci 2012, 13, 5645–5658. [Google Scholar]
- Nagaoka, T.; Ogihara, Y. Applicability of inter-simple sequence repeat polymorphisms in wheat for use as DNA markers in comparison to RFLP and RAPD markers. Theor. Appl. Genet 1997, 94, 597–602. [Google Scholar]
- Qian, W.; Ge, S.; Hong, D.Y. Genetic variation within and among populations of a wild rice Oryza granulata from China detected by RAPD and ISSR markers. Theor. Appl. Genet 2001, 102, 440–449. [Google Scholar]
- Wu, C.J.; Cheng, Z.Q.; Huang, X.Q.; Yin, S.-H.; Cao, K.M.; Sun, C.-R. Genetic diversity among and within populations of Oryza granulata from Yunnan of China revealed by RAPD and ISSR markers: Implications for conservation of the endangered species. Plant Sci 2004, 167, 35–42. [Google Scholar]
- Wang, C.; Zhang, H.; Qian, Z.Q.; Zhao, G.F. Genetic differentiation in endangered Gynostemma pentaphyllum (Thunb.) Makino based on ISSR polymorphism and its implications for conservation. Biochem. Syst. Ecol 2008, 36, 699–705. [Google Scholar]
- Yang, W.; de Oliveira, A.C.; Godwin, I.; Schertz, K.; Bennetzen, J.L. Comparison of DNA marker technologies in characterizing plant genome diversity: Variability in Chinese sorghums. Crop Sci 1996, 36, 1669–1676. [Google Scholar]
- Bornet, B.; Branchard, M. Nonanchored inter simple sequence repeat (ISSR) markers: Reproducible and specific tools for genome fingerprinting. Plant Mol. Biol. Report 2001, 19, 209–215. [Google Scholar]
- Liu, X.L.; Qian, Z.G.; Liu, F.H.; Yang, Y.W.; Pu, C.X. Genetic diversity within and among populations of Neopicrorhiza scrophulariiflora (Scrophulariaceae) in China, an endangered medicinal plant. Biochem. Syst. Ecol 2011, 39, 297–301. [Google Scholar]
- Yang, L.; Zhou, G.; Chen, G. Genetic diversity and population structure of Swertia tetraptera (Gentianaceae), an endemic species of Qinghai-Tibetan Plateau. Biochem. Syst. Ecol 2011, 39, 302–308. [Google Scholar]
- Zhao, X.; Ma, Y.; Sun, W.; Wen, X.; Milne, R. High genetic diversity and low differentiation of Michelia coriacea (Magnoliaceae), a critically endangered endemic in southeast Yunnan, China. Int. J. Mol. Sci 2012, 13, 4396–4411. [Google Scholar]
- Wang, D.L.; Li, Z.C.; Hao, G.; Chiang, T.Y.; Ge, X.J. Genetic diversity of Calocedrus macrolepis (Cupressaceae) in southwestern China. Biochem. Syst. Ecol 2004, 32, 797–807. [Google Scholar]
- Li, Z.C.; Wang, X.L.; Ge, X.J. Genetic diversity of the relict plant Taiwania cryptomerioides Hayata (Cupressaceae) in mainland China. Silvae Genet 2008, 57, 242–249. [Google Scholar]
- Xia, T.; Meng, L.; Mao, K.; Tian, B.; Miehe, G.; Liu, J. Genetic variation in the Qinghai-Tibetan plateau endemic and endangered conifer Cupressus gigantea, detected using RAPD and ISSR markers. Silvae Genet 2008, 57, 85–92. [Google Scholar]
- Hao, B.; Li, W.; Linchun, M.; Li, Y.; Rui, Z.; Mingxia, T.; Weikai, B. A study of conservation genetics in Cupressus chengiana, an endangered endemic of China, using ISSR markers. Biochem. Genet 2006, 44, 29–43. [Google Scholar]
- Hamrick, J.L.; Godt, M.; Sherman-Broyles, S. Factors influencing levels of genetic diversity in woody plant species. N. For 1992, 6, 95–124. [Google Scholar]
- Luan, S.; Chiang, T.Y.; Gong, X. High genetic diversity vs. low genetic differentiation in Nouelia insignis (Asteraceae), a narrowly distributed and endemic species in China, revealed by ISSR fingerprinting. Ann. Bot 2006, 98, 583–589. [Google Scholar]
- Wang, X.M.; Hou, X.Q.; Zhang, Y.Q.; Yang, R.; Feng, S.F.; Li, Y.; Ren, Y. Genetic diversity of the endemic and medicinally important plant Rheum officinale as revealed by inter-simpe sequence repeat (ISSR) markers. Int. J. Mol. Sci 2012, 13, 3900–3915. [Google Scholar]
- Leimu, R.; Mutikainen, P.I.A.; Koricheva, J.; Fischer, M. How general are positive relationships between plant population size, fitness and genetic variation? J. Ecol 2006, 94, 942–952. [Google Scholar]
- Qin, Q.; Zhang, C.; Milne, R. Population genetics and breeding system of Tupistra pingbianensis (Liliaceae), a naturally rare plant endemic to SW China. J. Syst. Evol 2010, 48, 47–57. [Google Scholar]
- Hilde, N.; Bartish, I.V. Effects of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants. Perspect. Plant Ecol. Evol. Syst 2000, 3, 93–114. [Google Scholar]
- Boratyński, A.; Lewandowski, A.; Boratyńska, K.; Montserrat, J.M.; Romo, A. High level of genetic differentiation of Juniperus phoenicea (Cupressaceae) in the Mediterranean region: Geographic implications. Plant Syst. Evol 2009, 277, 163–172. [Google Scholar]
- Li, F.; Xia, N. Population structure and genetic diversity of an endangered species, Glyptostrobus pensilis (Cupressaceae). Bot. Bull. Acad. Sin 2005, 46, 155–162. [Google Scholar]
- Pfeifer, M.; Jetschke, G. Influence of geographical isolation on genetic diversity of Himantoglossum hircinum (Orchidaceae). Folia Geobot 2006, 41, 3–20. [Google Scholar]
- Slatkin, M. Gene flow and the geographic structure of natural populations. Science 1987, 236, 787–792. [Google Scholar]
- Fischer, M.; Husi, R.; Prati, D.; Peintinger, M.; van Kleunen, M.; Schmid, B. RAPD variation among and within small and large populations of the rare clonal plant Ranunculus reptans (Ranunculaceae). Am. J. Bot 2000, 87, 1128–1137. [Google Scholar]
- Franklin, I.R. Evolutionary Change in Small Populations. In Conservation biology: An Evolutionary-Ecological Perspective; Soule, M., Wilcox, B., Eds.; Sinauer Associates: Sunderland, MA, USA, 1980; pp. 135–149. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull 1987, 19, 11–15. [Google Scholar]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar]
- Yeh, F.C.; Yang, R.; Boyle, T.; Ye, Z.; Mao, J.X. Popgene Version 1.32: The User Friendly Software for Population Genetic Analysis. In Molecular Biology and Biotechnology Centre; University of Alberta: Edmonton, AB, Canada, 1999. [Google Scholar]
- McDermott, J.M.; McDonald, B.A. Gene flow in plant pathosystems. Annu. Rev. Phytopathol 1993, 31, 353–373. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res 1967, 27, 209–220. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pop | N | Locality | Latitude (° N) | Longitude (° E) | Altitude (m) |
|---|---|---|---|---|---|
| A | 8 | Erchongyan, Kaixian county | 31.608 | 108.712 | 2290 |
| B | 24 | Anjiacao, Kaixian county | 31.616 | 108.754 | 2150 |
| C | 30 | Wangjiayan, Kaixian county | 31.609 | 108.672 | 1840 |
| D | 30 | Yangcun, Kaixian county | 31.636 | 108.811 | 1770 |
| E | 9 | Jigongliang, Kaixian county | 31.597 | 108.825 | 1270 |
| F | 13 | Geteng, Chengkou county | 31.651 | 108.675 | 1615 |
| G | 25 | Longmen, Chengkou county | 31.715 | 108.762 | 1005 |
| Primer | Sequence (5′–3′) | Primer | Sequence (5′–3′) |
|---|---|---|---|
| UBC807 | AGA GAG AGA GAG AGA GT | UBC841 | GAG AGA GAG AGA GAG AYC |
| UBC808 | AGA GAG AGA GAG AGA GC | UBC842 | GAG AGA GAG AGA GAG AYG |
| UBC809 | AGA GAG AGA GAG AGA GG | UBC848 | CAC ACA CAC ACA CAC AR*G |
| UBC810 | GAG AGA GAG AGA GAG AT | UBC878 | GGA TGG ATG GAT GGA T |
| UBC811 | GAG AGA GAG AGA GAG AC | UBC880 | GGA GAG GAG AGG AGA |
| UBC820 | GTG TGT GTG TGT GTG TC | UBC886 | VDV CTC TCT CTC TCT CT |
| UBC826 | ACA CAC ACA CAC ACA CC | UBC895 | AGA GTT GGT AGC TCT TGA TC |
| UBC836 | AGA GAG AGA GAG AGA GY*A |
| A | B | C | D | E | F | G | Mean | Total | |
|---|---|---|---|---|---|---|---|---|---|
| Na | 1.597 | 1.739 | 1.913 | 1.881 | 1.648 | 1.690 | 1.861 | 1.761 | 2.000 |
| Ne | 1.214 | 1.180 | 1.226 | 1.193 | 1.215 | 1.237 | 1.220 | 1.212 | 1.220 |
| P | 59.7 | 73.9 | 91.3 | 88.1 | 64.8 | 69.0 | 86.1 | 76.1 | 100.0 |
| H | 0.152 | 0.130 | 0.161 | 0.142 | 0.157 | 0.163 | 0.178 | 0.155 | 0.165 |
| I | 0.232 | 0.180 | 0.272 | 0.247 | 0.245 | 0.256 | 0.291 | 0.252 | 0.295 |
| HT | 0.167 | ||||||||
| Hs | 0.150 | ||||||||
| Gst | 0.102 | ||||||||
| Nm | 4.407 |
| Source of variation | d.f.* | Sum of squares | Variance component | Percentage of variance | p value |
|---|---|---|---|---|---|
| Among population | 6 | 783.993 | 4.938 | 12.092 | <0.01 |
| Within population | 132 | 4738.597 | 35.898 | 87.908 | <0.01 |
| Pop | A | B | C | D | E | F | G |
|---|---|---|---|---|---|---|---|
| A | ---- | 0.0218 | 0.0188 | 0.0210 | 0.0160 | 0.0121 | 0.0250 |
| B | 4.12 | ---- | 0.0134 | 0.0138 | 0.0220 | 0.0186 | 0.0167 |
| C | 3.78 | 7.84 | ---- | 0.0120 | 0.0186 | 0.0183 | 0.0130 |
| D | 9.87 | 5.77 | 13.49 | ---- | 0.0212 | 0.0201 | 0.0163 |
| E | 10.79 | 7.04 | 14.55 | 4.48 | ---- | 0.0064 | 0.0202 |
| F | 5.92 | 8.43 | 4.72 | 12.97 | 15.41 | ---- | 0.0215 |
| G | 12.76 | 10.93 | 14.54 | 9.89 | 14.33 | 10.85 | ---- |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, J.; Shi, S.; Chang, E.; Yang, W.; Jiang, Z. Genetic Diversity of the Critically Endangered Thuja sutchuenensis Revealed by ISSR Markers and the Implications for Conservation. Int. J. Mol. Sci. 2013, 14, 14860-14871. https://doi.org/10.3390/ijms140714860
Liu J, Shi S, Chang E, Yang W, Jiang Z. Genetic Diversity of the Critically Endangered Thuja sutchuenensis Revealed by ISSR Markers and the Implications for Conservation. International Journal of Molecular Sciences. 2013; 14(7):14860-14871. https://doi.org/10.3390/ijms140714860
Chicago/Turabian StyleLiu, Jianfeng, Shengqing Shi, Ermei Chang, Wenjuan Yang, and Zeping Jiang. 2013. "Genetic Diversity of the Critically Endangered Thuja sutchuenensis Revealed by ISSR Markers and the Implications for Conservation" International Journal of Molecular Sciences 14, no. 7: 14860-14871. https://doi.org/10.3390/ijms140714860