Impacts of pr-10a Overexpression at the Molecular and the Phenotypic Level

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Magnitude of pr-10a Expression Induction Is Sensitive to both the Chemical Compound and the Strength of Osmotic Pressure
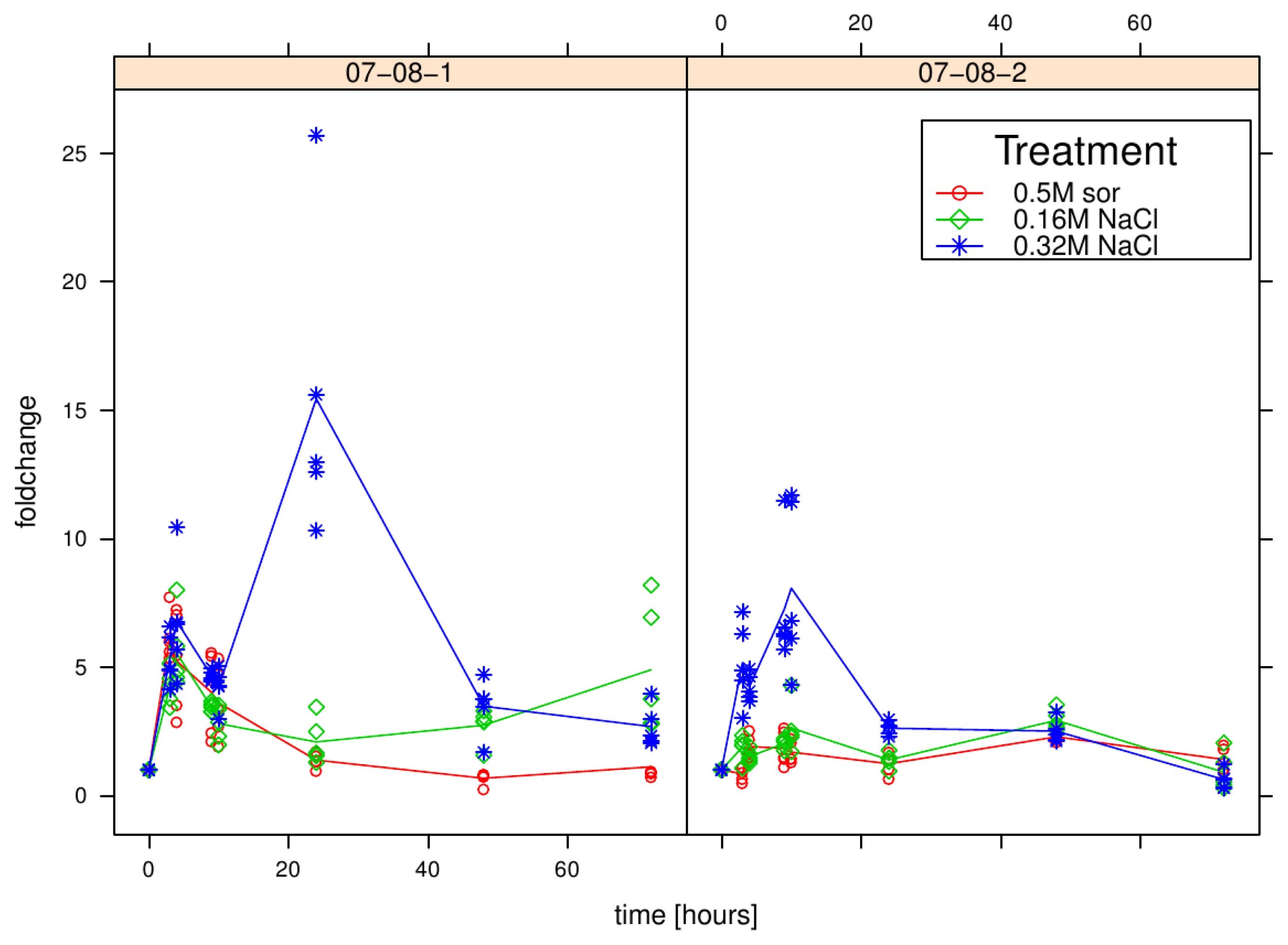
2.2. Fast Induction of pr-10a Expression Regulators Sebf and pti4
2.3. Mannopine-synthase Promoter is Induced by Strong Salt Treatments
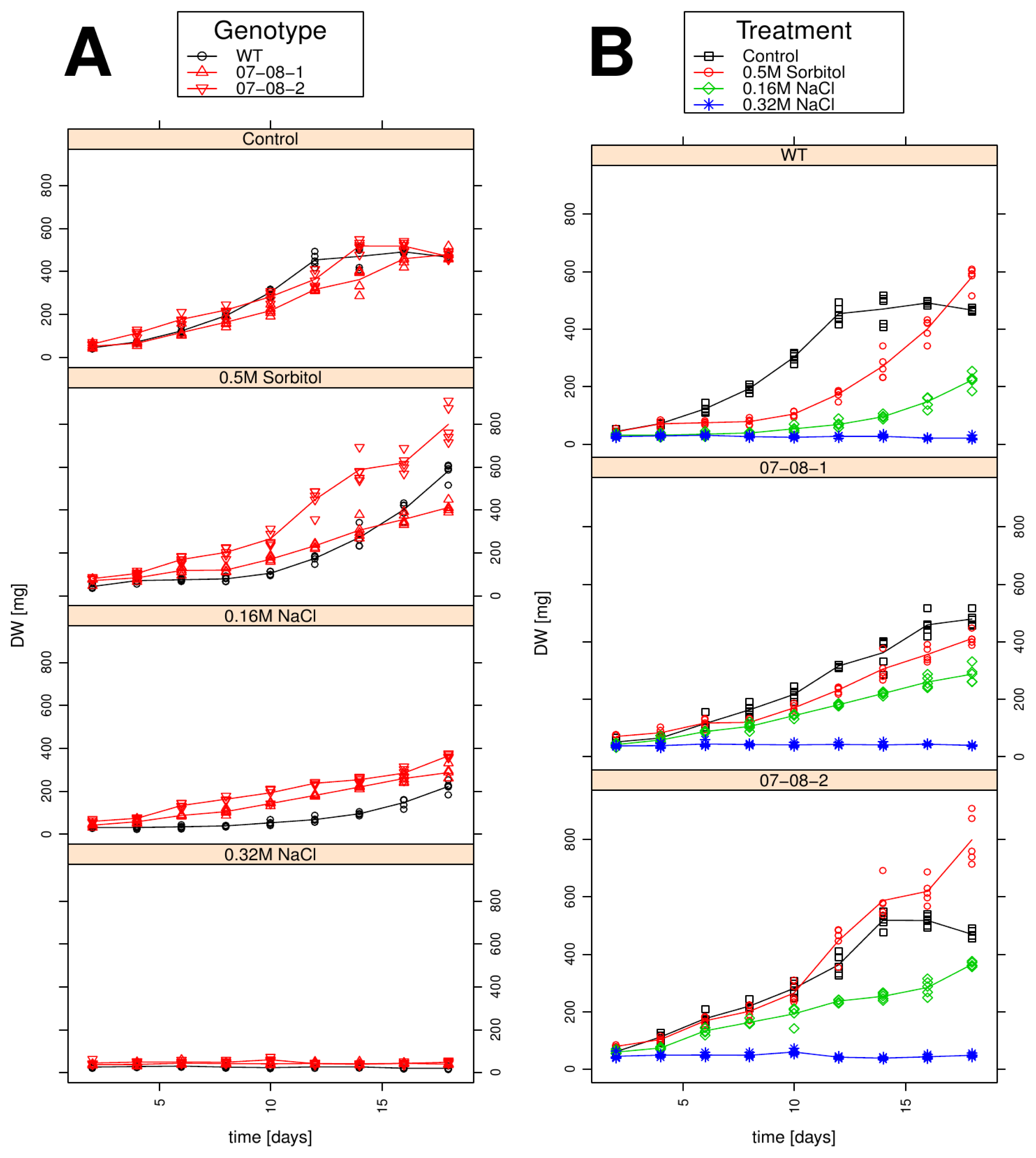
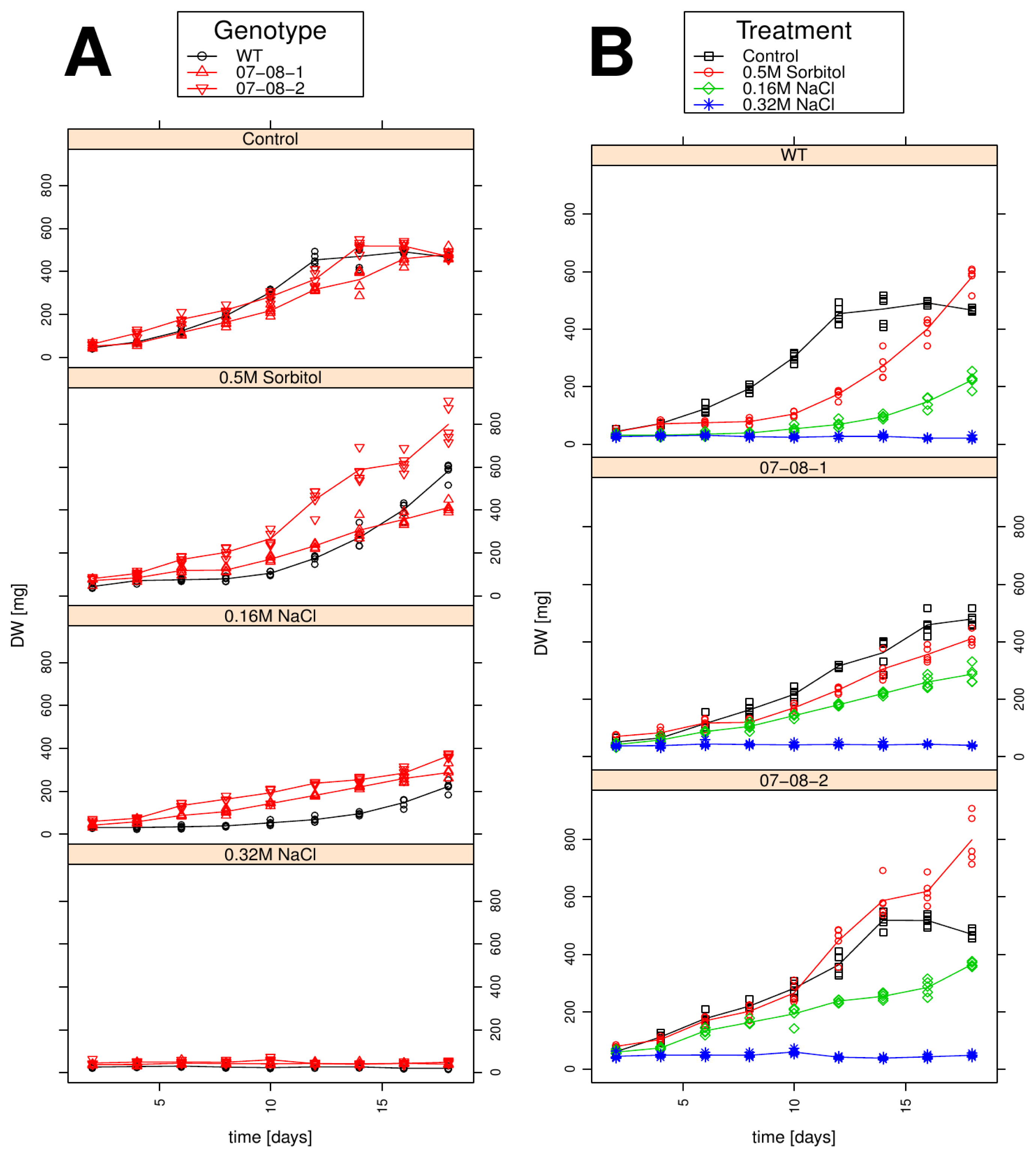
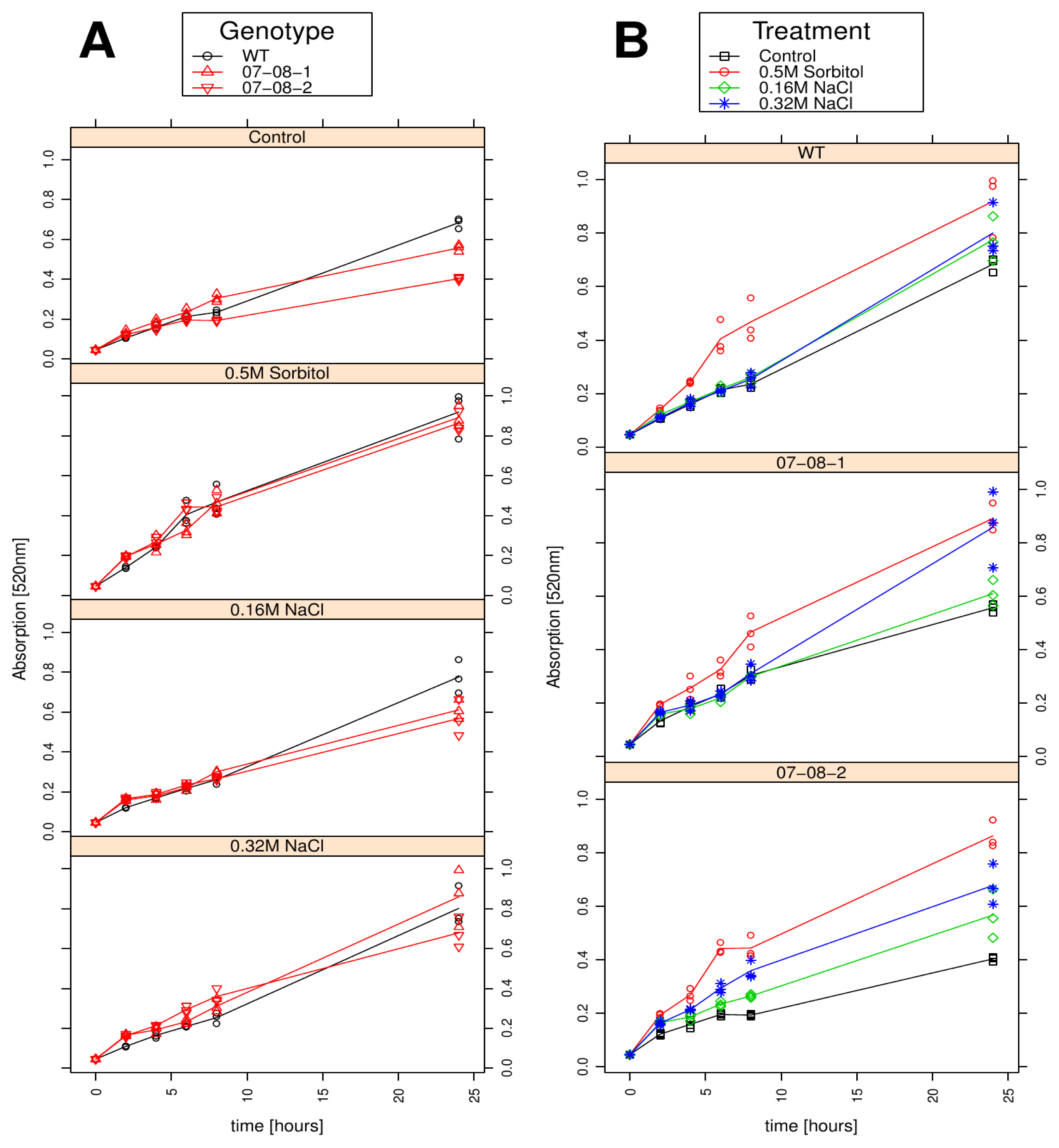
2.4. Growth Behaviour Is Influenced by both the Chemical Compound and the Strength of the Osmotic Pressure
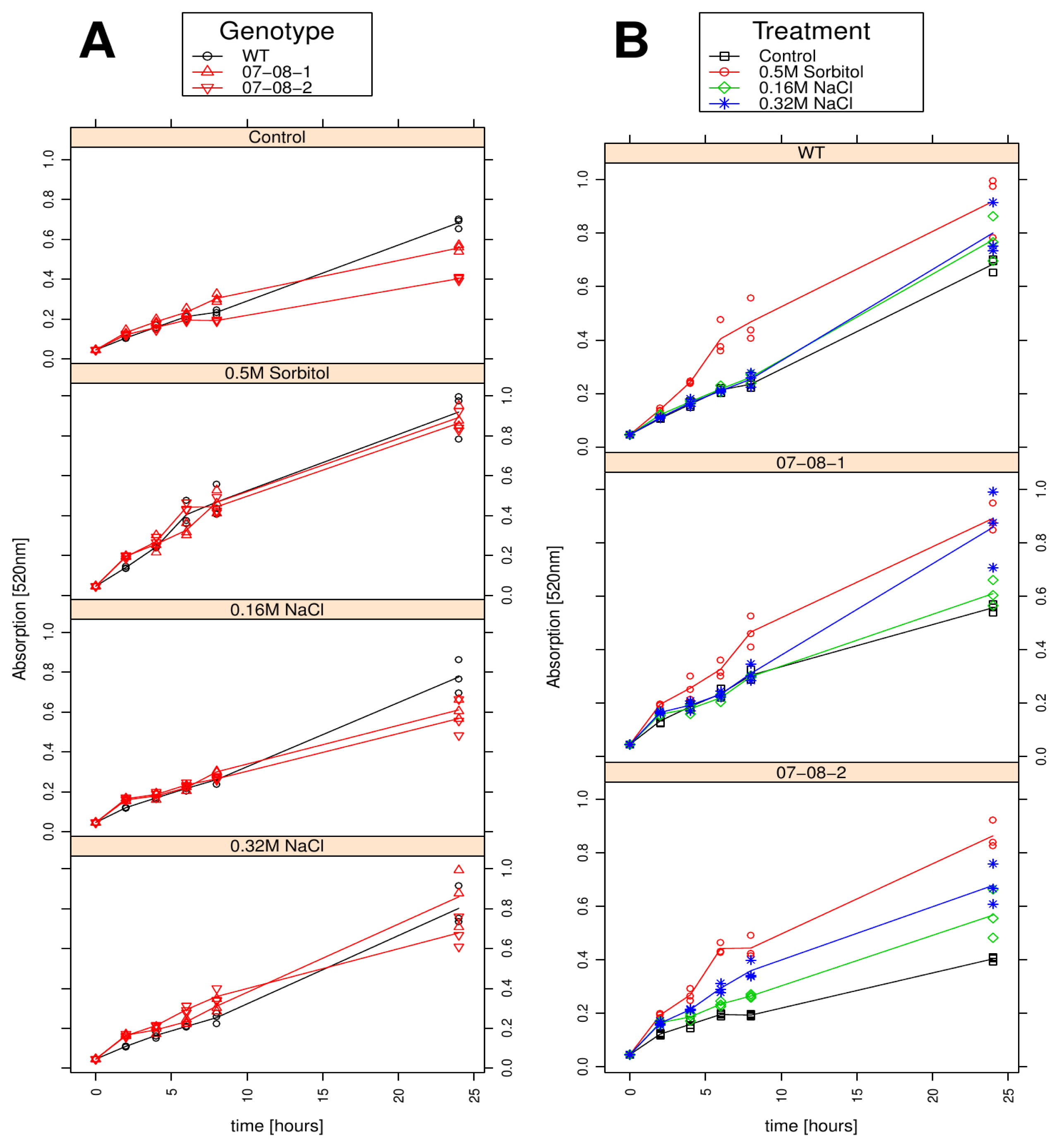
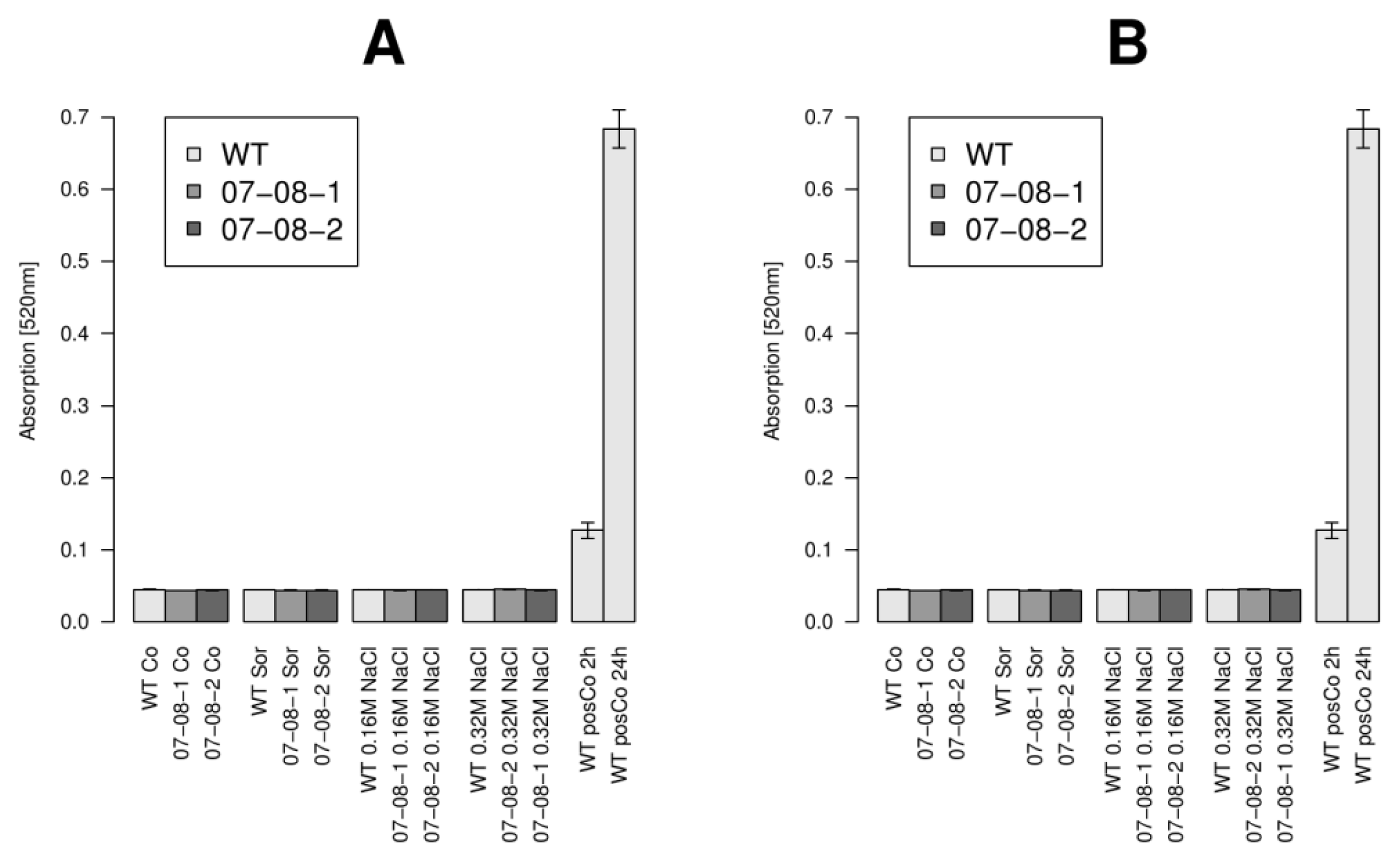
2.5. Cell Respiration Reacts on both the Chemical Compound and the Strength of Osmotic Pressure
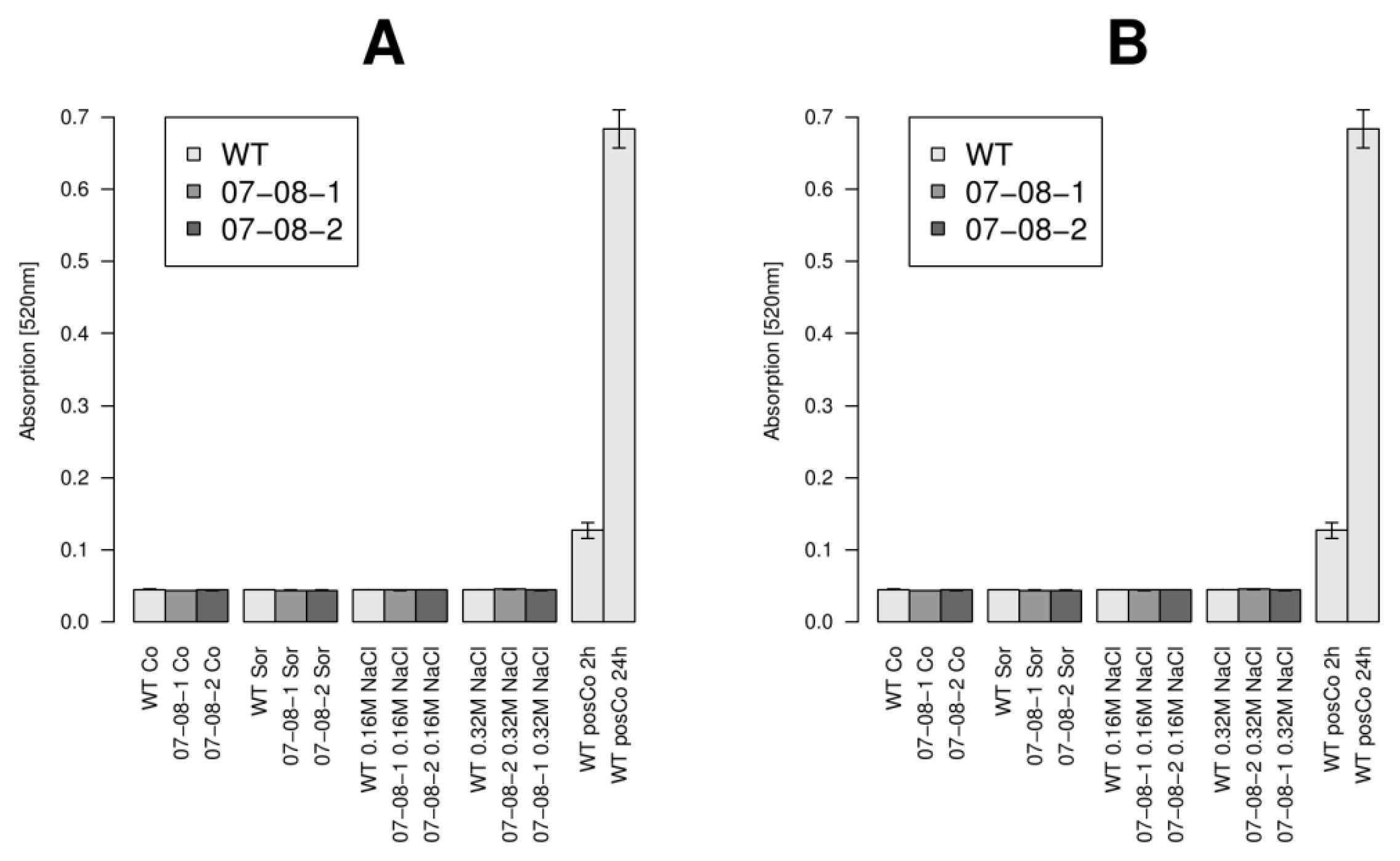
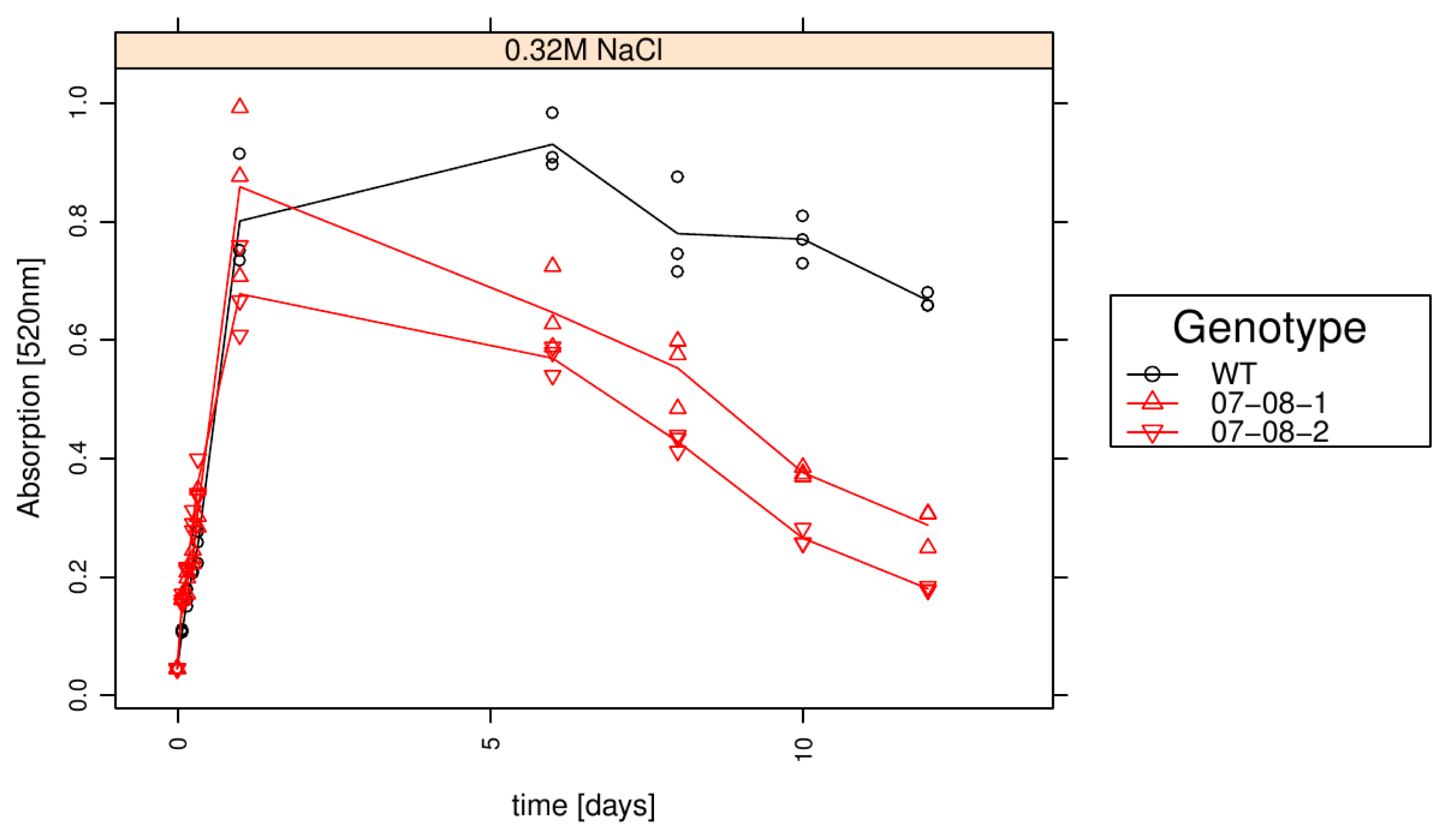
2.6. Wild-Type Cells Showed Sustained Respiration Capacity under Severe 0.32M NaCl
3. Discussion
3.1. pr-10a Expression Induction by a Common Stimulus
3.2. Interaction between Treatments and Transcriptional Activity of the Transgene
3.3. Importance of Phenotypes Derived from Cellular Level
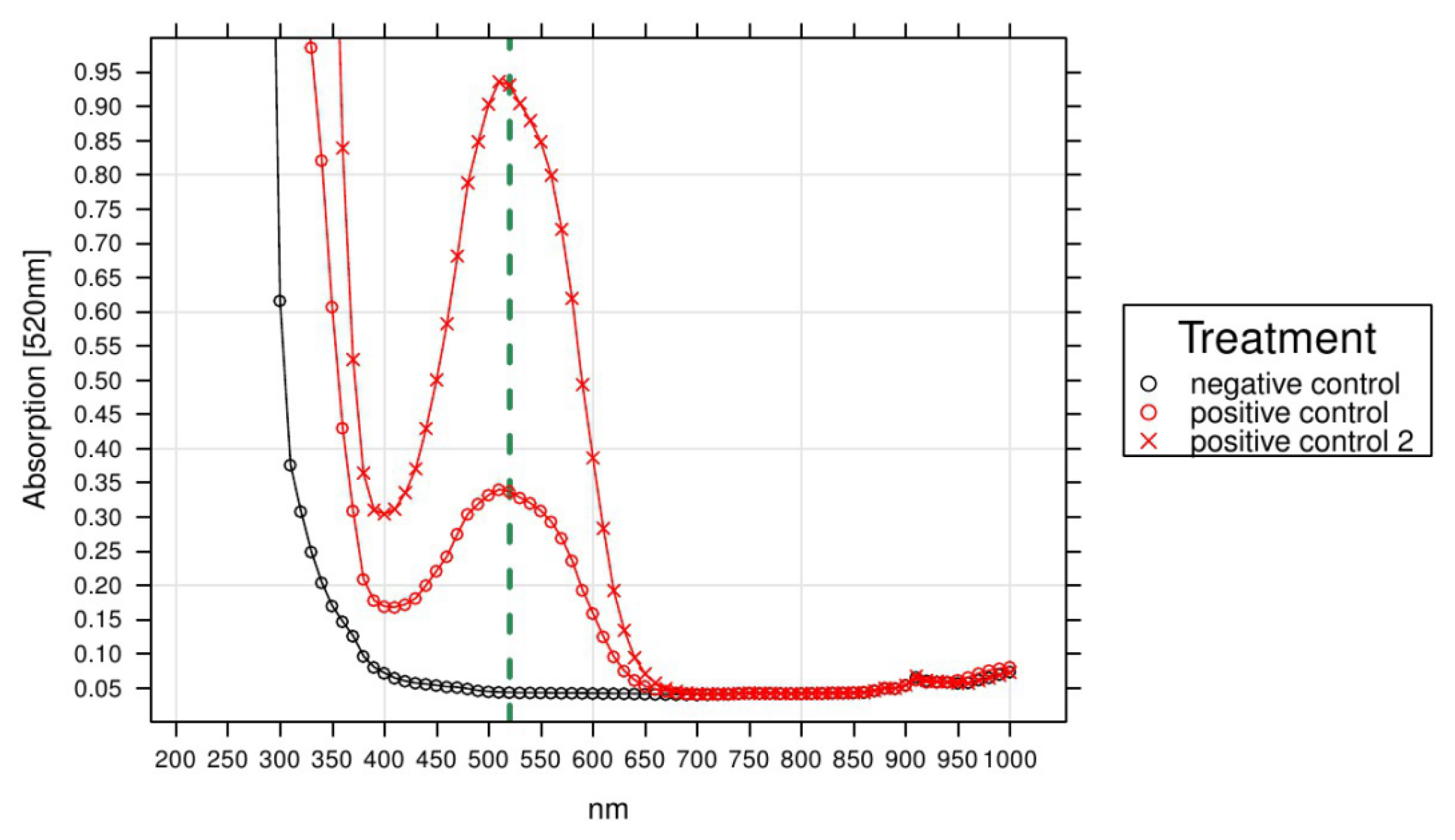
3.4. Technical Aspects of Respiration Measurements in Plant Cell Cultures
4. Experimental Section
4.1. Plant Material and Osmotic Challenge
4.2. RNA Isolation and Quantitative Real-Time PCR
4.3. Dry-Weight Determination of Cell Material
4.4. Respiration Curves
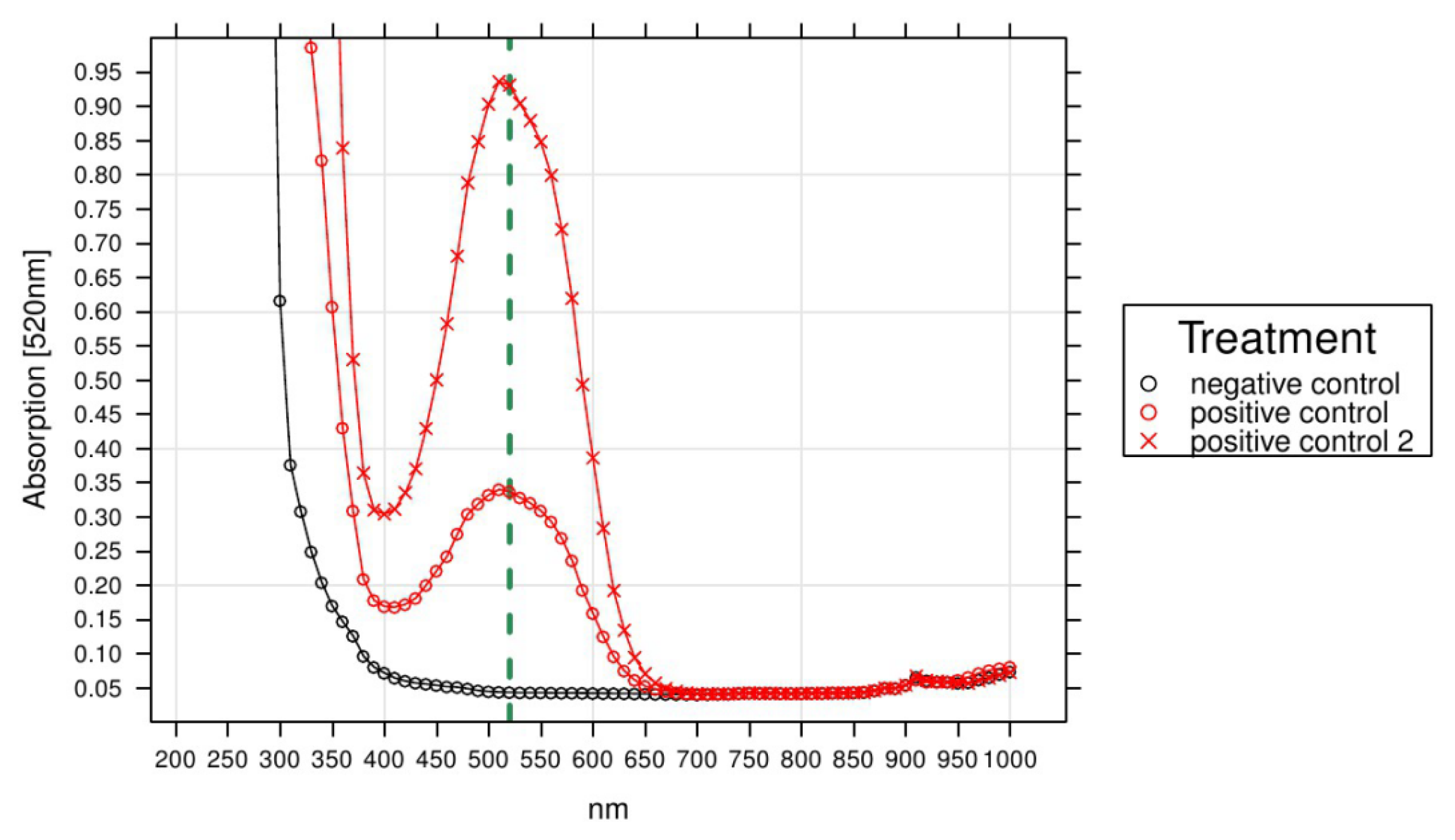
4.5. Assessment of Cell Survival via Respiration Measurements
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
1. Monitoring of Gene-Expression
2. Validation of TTC-Based Respiration Curves
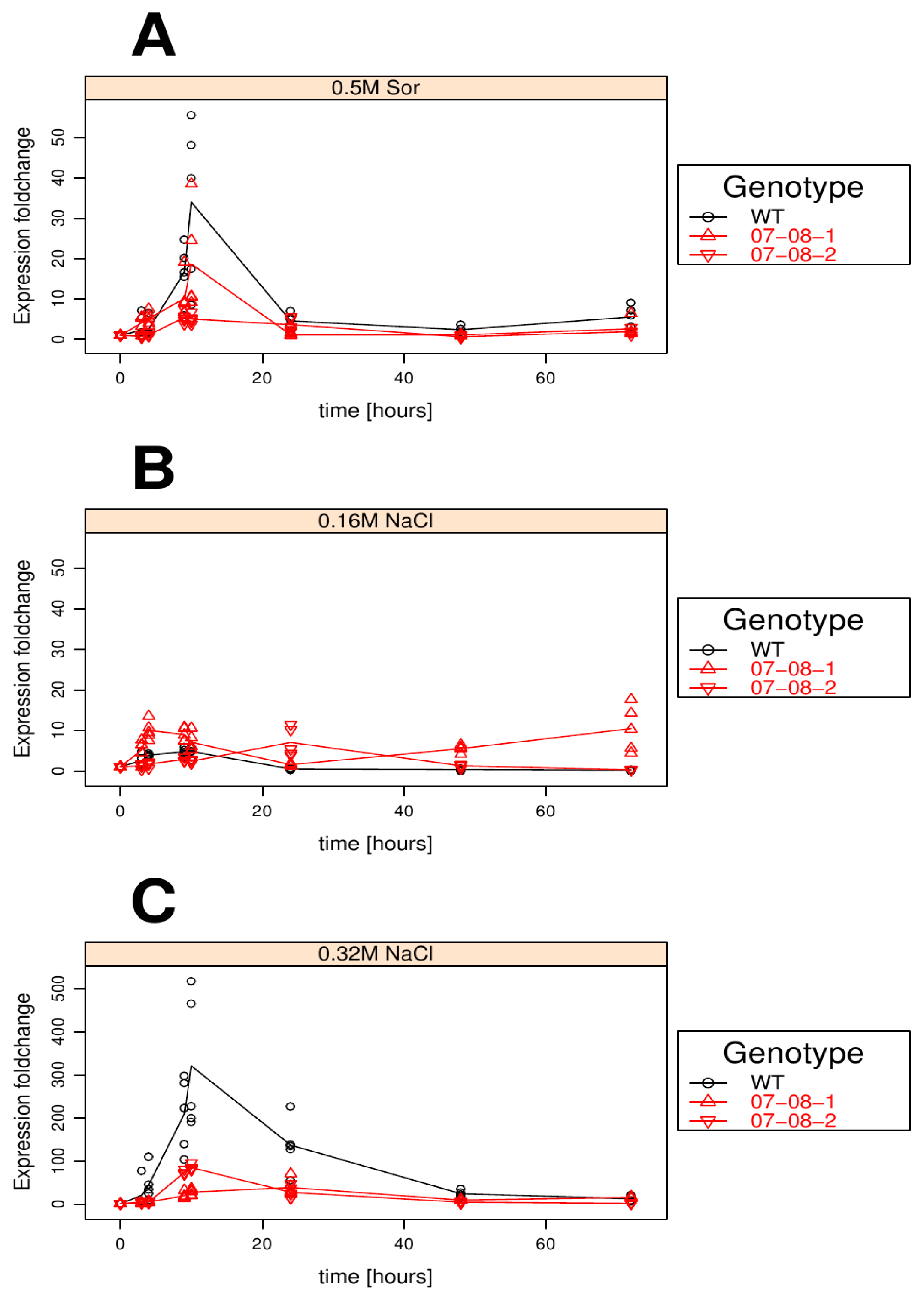
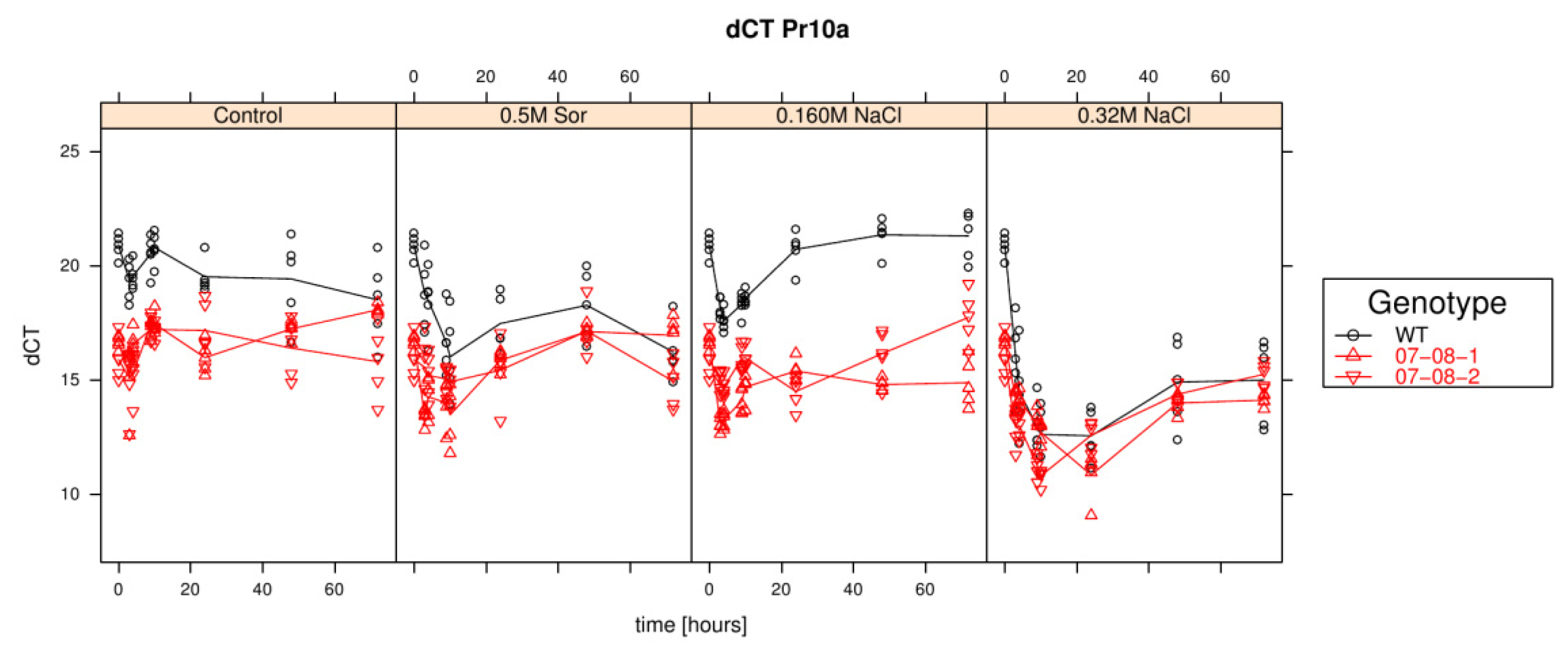
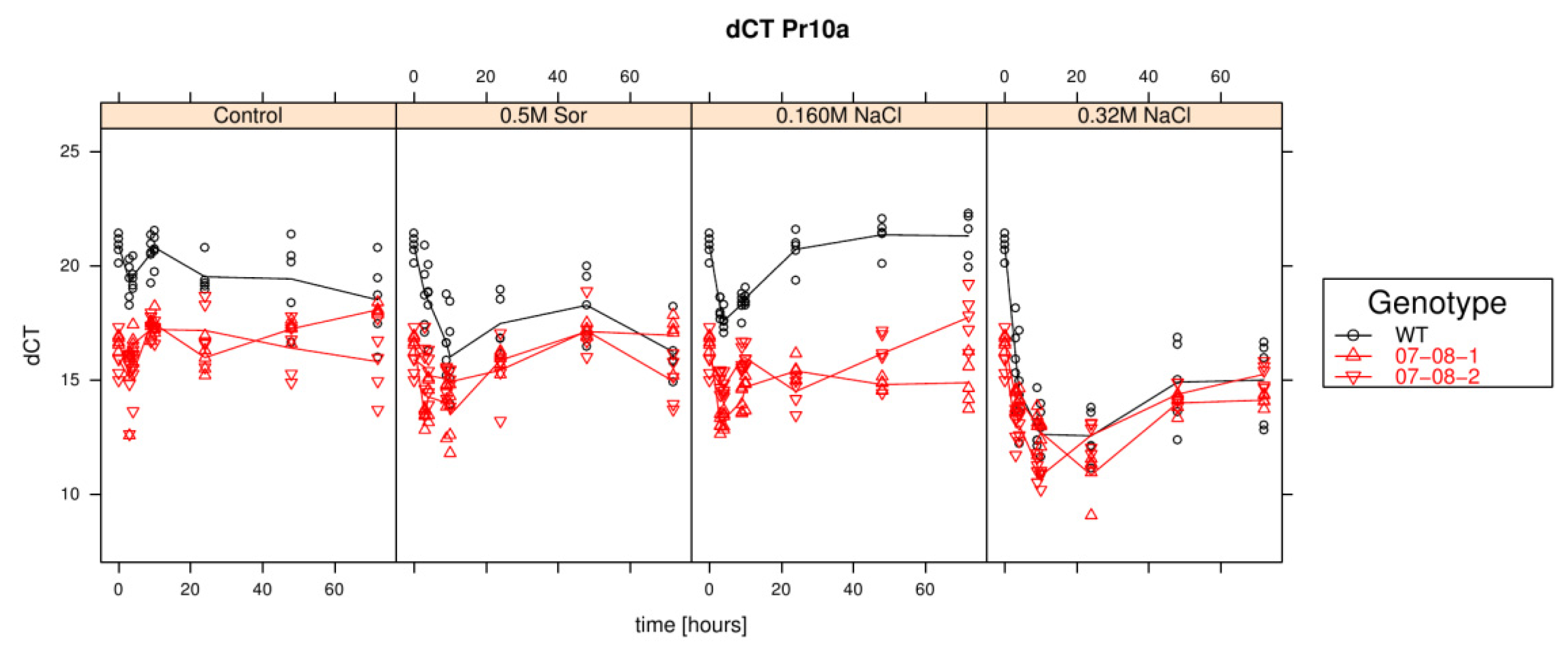
Results of relative pr-10a expression analysis of cells treated with differently supplemented medium (control, 0.5 M sorbitol, 0.32 M NaCl, 0.16 M NaCl). Given are the expressions levels standardized to their 18S rRNA threshold cycle (ΔCt). Wild type (WT) is encoded by black lines and circles, transgenic cell cultures (07-08-1 and 07-08-2, respectively) by red lines and triangles, while symbols indicate single measurement points and lines corresponding group means. Note that smaller ΔCt values indicate a stronger expression [1].
{kind=link}
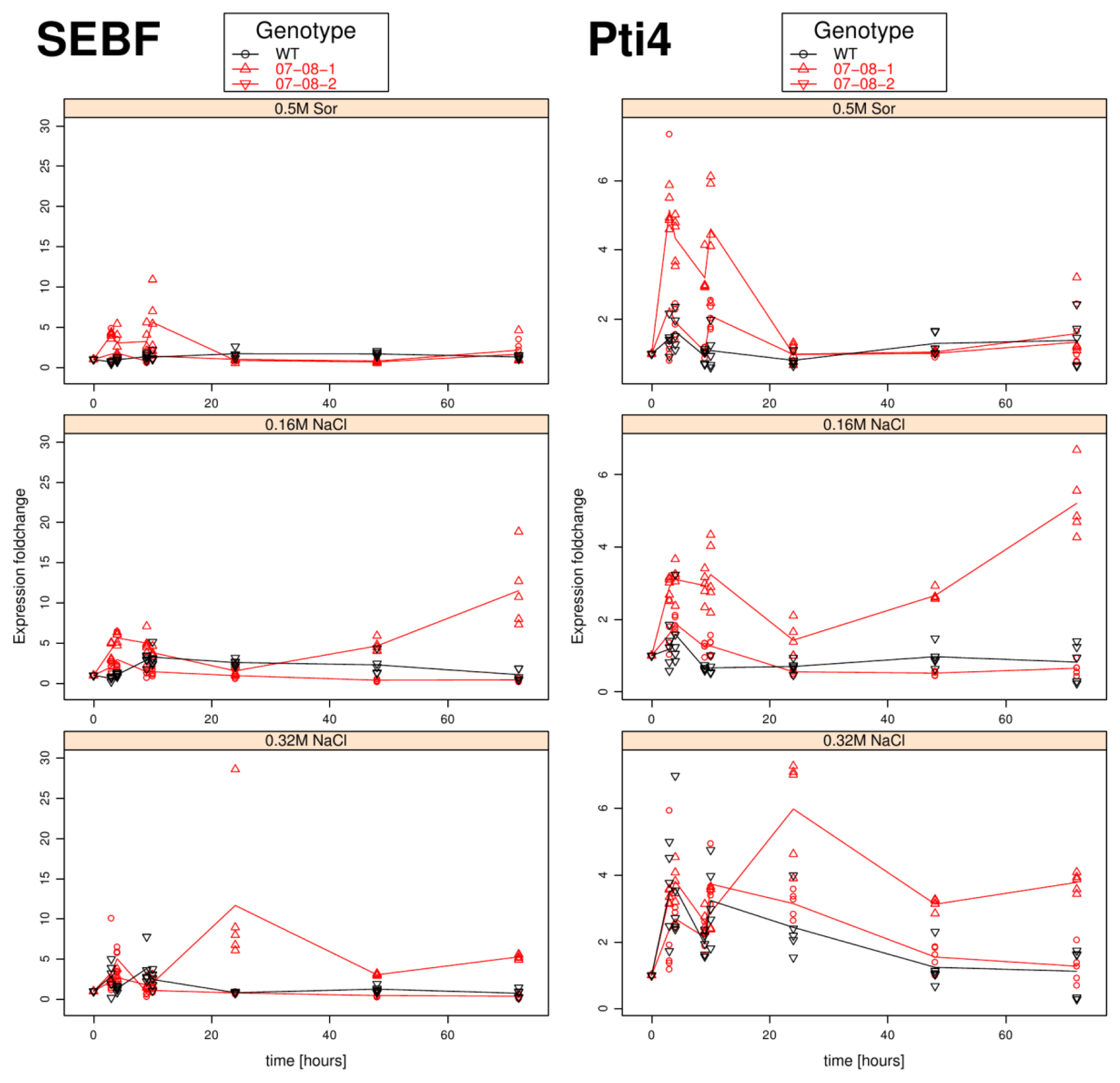
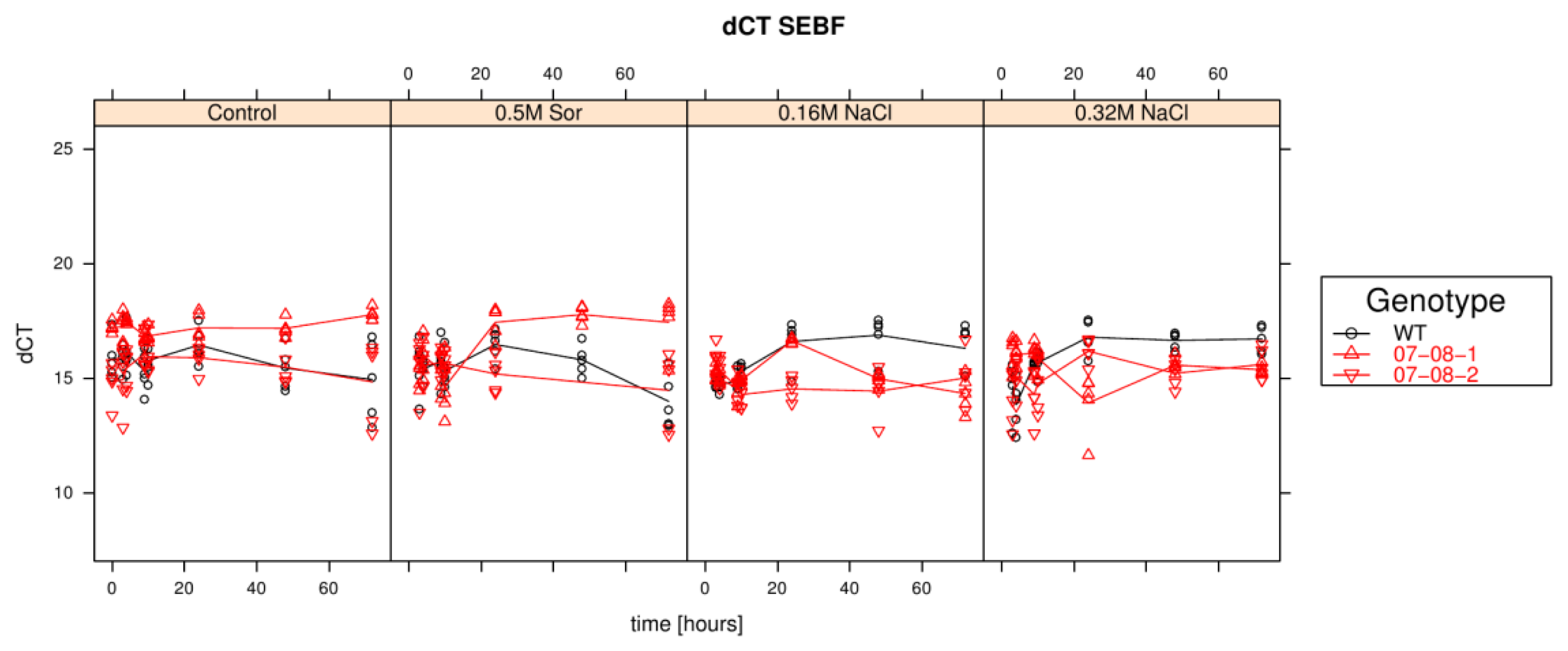
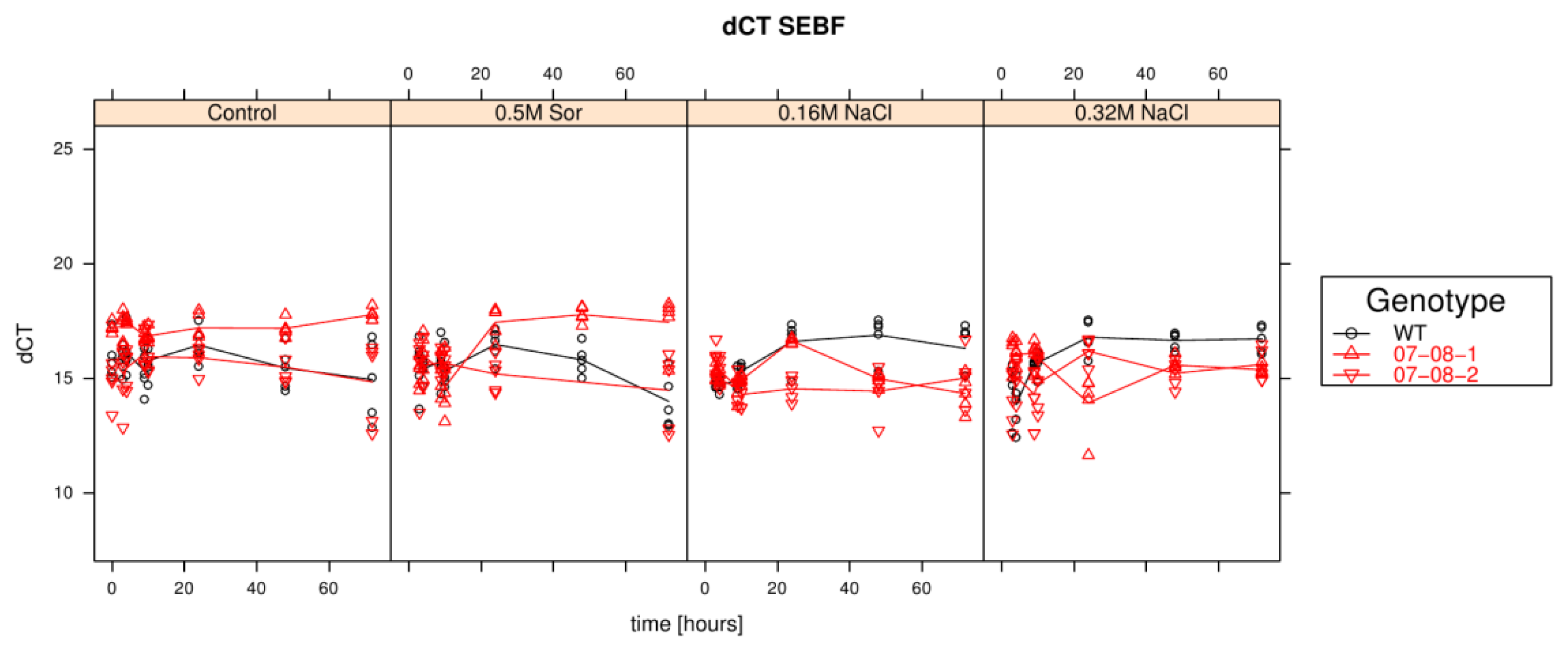
Results of relative sebf expression analysis of cells treated with differently supplemented medium (control, 0.5 M sorbitol, 0.32 M NaCl, 0.16 M NaCl). Given are the expressions levels standardized to their 18S rRNA threshold cycle (ΔCt). Wild type (WT) is encoded by black lines and circles, transgenic cell cultures (07-08-1 and 07-08-2, respectively) by red lines and triangles, while symbols indicate single measurement points and lines corresponding group means. Note that smaller ΔCt values indicate a stronger expression [1].
{kind=link}
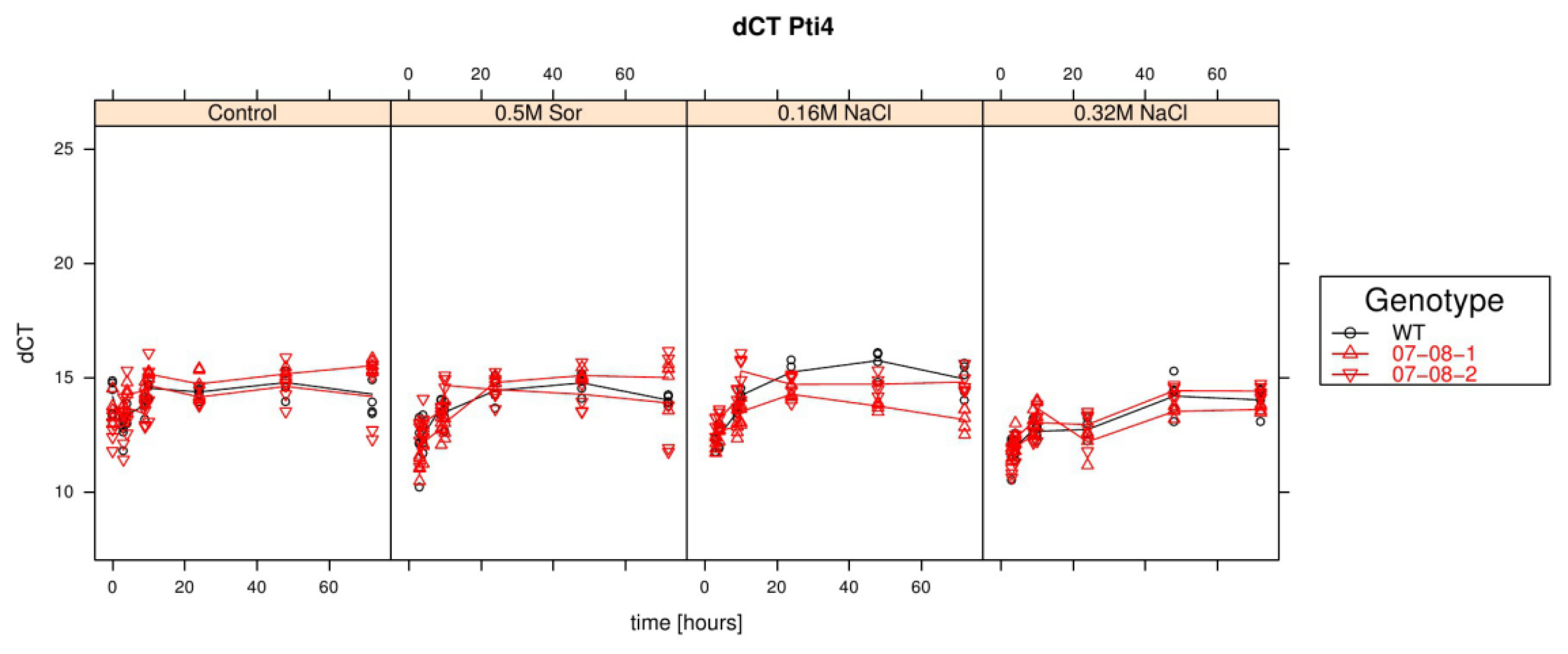
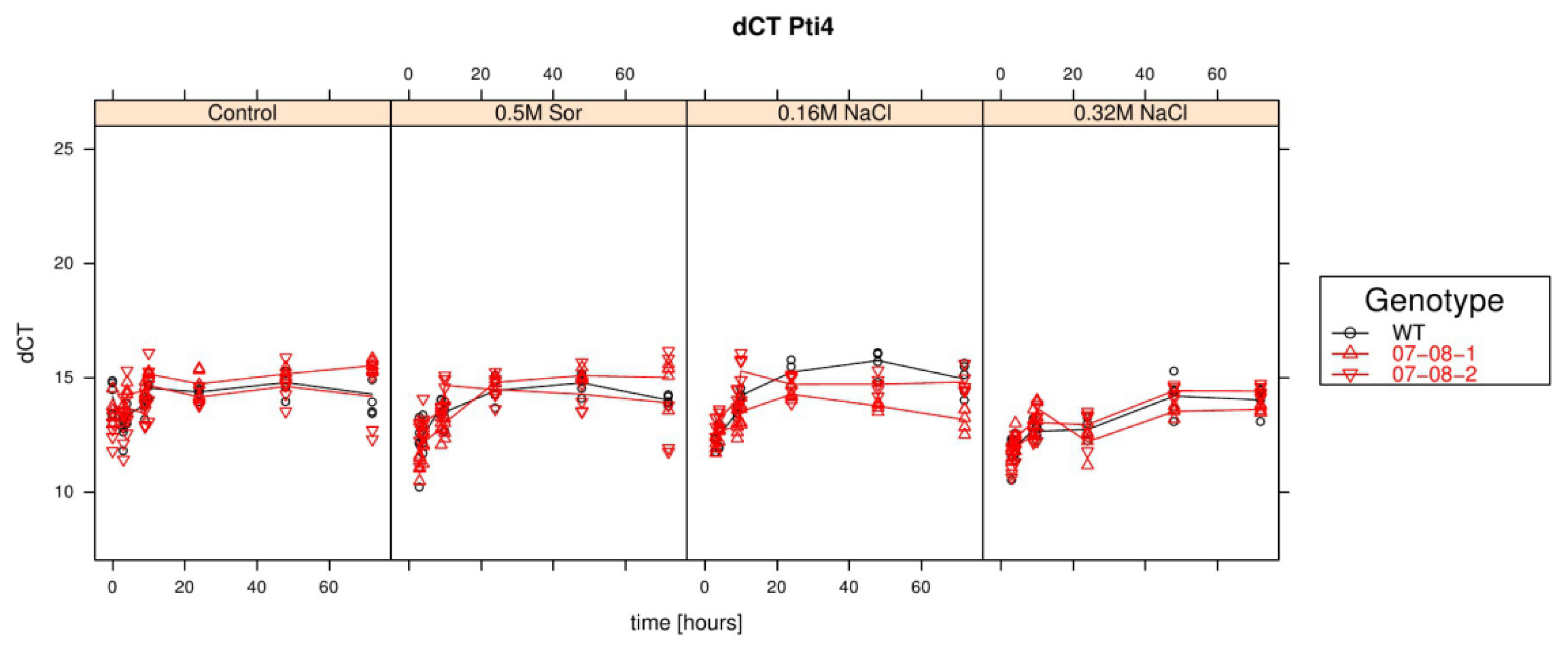
Results of relative pti4 expression analysis of cells treated with differently supplemented medium (control, 0.5 M sorbitol, 0.32 M NaCl, 0.16 M NaCl). Given are the expressions levels standardized to their 18S rRNA threshold cycle (ΔCt). Wild type (WT) is encoded by black lines and circles, transgenic cell cultures (07-08-1 and 07-08-2, respectively) by red lines and triangles, while symbols indicate single measurement points and lines corresponding group means. Note that smaller ΔCt values indicate a stronger expression [47].
{kind=link}
{kind=link}
{kind=link}
Conflict of Interest
References
- Gaspar, T.; Franck, T.; Bisbis, B.; Kevers, C.; Jouve, L.; Hausman, J.F.; Dommes, J. Concepts in plant stress physiology. Application to plant tissue cultures. Plant Growth Regul 2002, 37, 263–285. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol 2008, 59, 651–681. [Google Scholar]
- Vaas, L.A.I.; Marheine, M.; Seufert, S.; Schumacher, H.-M.; Kiesecker, H.; Heine-Dobbernack, E. Impact of pr-10a overexpression on the cryopreservation success of Solanum tuberosum suspension cultures. Plant Cell Rep 2012, 31, 1061–1071. [Google Scholar]
- Kav, N.N.V.; Srivastava, S.; Goonewardene, L.; Blade, S.F. Proteome-level changes in the roots of Pisum sativum in response to salinity. Ann. Appl. Biol 2004, 145, 217–230. [Google Scholar]
- Jain, S.; Srivastava, S.; Sarin, N.B.; Kav, N.N.V. Proteomics reveals elevated levels of PR10 proteins in saline-tolerant peanut (Arachis hypogaea) calli. Plant Physiol. Biochem 2006, 44, 253–259. [Google Scholar]
- Jellouli, N.; Ben, J.H.; Skouri, H.; Ghorbel, A.; Gourgouri, A.; Mliki, A. Proteomic analysis of Tunisian grapevine cultivar Razegui under salt stress. J. Plant Physiol 2008, 165, 471–481. [Google Scholar]
- Jellouli, N.; Jouira, B.H.; Daldoul, S.; Chenennaoui, S.; Ghorbel, A.; Asma, A.S.; Gargouri, A. Proteomic and transcriptomic analysis of grapevine PR10 expression during salt stress and functional characterization in yeast. Plant Mol. Biol. Report 2010, 28, 1–8. [Google Scholar]
- Marineau, C.; Matton, D.P.; Brisson, N. Differential accumulation of potato tuber mRNA’s during the hypersensitive response induced by arachidonic acid elicitor. Plant Mol. Biol 1987, 9, 335–342. [Google Scholar]
- Matton, P.; Brisson, N. Cloning, Expression and Sequence Conservation of pathogenesis related gene transcripts of potato. Mol. Plant-Microbe Interact 1989, 2, 326–331. [Google Scholar]
- Constabel, C.P.; Brisson, N. Stigma- and vascular-specific expression of the pr-10a gene of potato: A novel pattern of expression of a pathogenesis-related gene. Mol. Plant-Microbe Interact 1995, 8, 104–1138. [Google Scholar]
- Utriainen, M.; Kokko, H.; Auriola, S.; Sarrazin, O.; Kärenlampi, S. PR-10 protein is induced by copper stress in roots and leaves of a Cu/Zn tolerant clone of birch, Betula pendula. Plant Cell Environ 1998, 21, 821–828. [Google Scholar]
- Hashimoto, M.; Kisseleva, L.; Sawa, S.; Furukawa, T.; Komatsu, S.; Koshiba, T. A novel rice PR10 protein, RSOsPR10, specifically induced in roots by biotic and abiotic stresses, possibly via the Jasmonic Acid signaling pathway. Plant Cell Physiol 2004, 45, 550–559. [Google Scholar]
- Mur, L.A.J.; Sturgess, F.J.; Farrell, G.G.; Draper, J. The AoPR10 promoter and certain endogenous PR10 genes respond to oxidative signals in Arabidopsis. Mol. Plant Pathol 2004, 5, 435–451. [Google Scholar]
- Ukaji, N.; Kuwabara, C.; Takezawa, D.; Arakawa, K.; Fujikawa, S. Accumulation of pathogenesis-related (PR) 10/Bet v 1 protein homologues in mulberry (Morus bombycis Koidz.) during winter. Plant Cell Environ 2004, 27, 1112–1121. [Google Scholar]
- Crowell, D.N.; John, M.E.; Russel, D.; Amasona, R. Characterization of a stress-induced, developmentally regulated gene family from soybean. Plant Mol. Biol 1992, 18, 459–466. [Google Scholar]
- Walter, M.H.; Liu, J.W.; Wünn, J.; Hess, D. Bean ribonuclease-like pathogenesis-related protein gene (Ypr10) display complex patterns of developmental, dark-induced and exogenous-stimulus-dependent expression. Eur. J. Biochem 1996, 239, 281–293. [Google Scholar]
- Liu, J.J.; Ekramoddoullah, A.K.M. The family 10 of plant pathogenesis-related proteins: Their structure, regulation, and function in response to biotic and abiotic stresses. Physiol. Mol. Plant Pathol 2006, 68, 3–13. [Google Scholar]
- Srivastava, S.; Fristensky, B.; Kav, N.N.V. Constitutive expression of a PR10 protein enhances the germination of Brassica napus under saline conditions. Plant Cell Physiol 2004, 45, 1320–1324. [Google Scholar]
- El-Banna, A.; Hajirezaei, M.R.; Wissing, J.; Ali, Z.; Vaas, L.; Heine-Dobbernack, E.; Jacobsen, H.-J.; Schumacher, H.-M.; Kiesecker, H. Over-expression of PR-10a leads to increased salt and osmotic tolerance in potato cell cultures. J. Biotechnol 2010, 150, 277–287. [Google Scholar]
- Constabel, C.P.; Bertrand, C.; Brisson, N. Transgenic potato plants overexpressing the pathogenesis-related STH-2 gene show unaltered susceptibility to Phytophthora infestans and potato virus X. Plant Mol. Biol 1993, 22, 775–782. [Google Scholar]
- Colditz, F.; Niehaus, K.; Kajinski, F. Silencing of PR-10-like proteins in Medicago truncatula results in an antagonistic induction of other PR proteins an in an increased tolerance upon infection with the oomycete Aphanomyces euteiches. Planta 2007, 226, 57–71. [Google Scholar]
- Gonzalez-Lamothe, R.; Boyle, P.; Dulude, A.; Roy, V.; Lezin-Doumbou, C.; Kaur, G.S.; Bouarab, K.; Despres, C.; Brisson, N. The transcriptional activator Pti4 is required for the recruitment of a repressosome nucleated by repressor SEBF at the potato PR-10a gene. Plant Cell 2008, 20, 3136–3147. [Google Scholar]
- Després, C.; Subramaniam, R.; Matton, D.P.; Brisson, N. The activation of the potato PR-10a gene requires the phosphorylation of the nuclear factor PBF-1. Plant Cell 1995, 7, 589–598. [Google Scholar]
- Desveaux, D.; Després, C.; Joyeux, A.; Subramaniam, R.; Brisson, N. PBF-2 is a novel single-stranded DNA binding factor implicated in PR-10a gene activation in potato. Plant Cell 2000, 12, 1477–1489. [Google Scholar]
- Boyle, B.; Brisson, N. Repression of the defense gene PR-10a by the single-stranded DNA binding protein SEBF. Plant Cell 2001, 13, 2525–2537. [Google Scholar]
- Bantignies, B.; Seguin, J.; Muzac, I.; Dedaldechamp, F.; Gulick, P.; Ibrahim, R. Direct evidence for ribonucleolytic activity of a PR-10-like protein from white lupin roots. Plant Mol. Biol 2000, 42, 871–881. [Google Scholar]
- Zubini, P.; Zambelli, B.; Musiani, F.; Ciurli, S.; Bertolini, P.; Baraldi, E. The RNA hydrolysis and the cytokinin binding activities of PR-10 proteins are differently performed by two isoforms of the Pru p1 peach major allergen and are possibly functionally related. Plant Physiol 2009, 150, 1235–1247. [Google Scholar]
- Fujimoto, Y.; Nagata, R.; Fukasawa, H.; Yano, K.; Azuma, M.; Iida, A.; Sugimoto, S.; Shudo, K.; Hashimoto, Y. Purification and cDNA cloning of cytokininspecific binding protein from mung bean (Vigna radiata). Eur. J. Biochem 1998, 258, 794–802. [Google Scholar]
- Gonneau, M.; Pagant, S.; Brun, F.; Laloue, M. Photoaffinity labelling with the cytokinin agonist acido-CPPU of a 34 kDa peptide of the intracellular pathogenesis-related protein family in the moss Physcomitrella patens. Plant Mol. Biol 2001, 46, 539–548. [Google Scholar]
- Mogensen, J.E.; Wimmer, R.; Larsen, J.N.; Spangfort, M.D. The major birch allergen, Bet v 1, shows affinity for a broad spectrum of physiological ligands. J. Biol. Chem 2002, 277, 684–692. [Google Scholar]
- Markovic-Housely, Z.; Degano, M.; Lamba, D.; von Roepenack-Lahaye, E.; Clemens, S.; Susani, M.; Ferreira, F.; Scheiner, O.; Breiteneder, H. Crystal structure of a hypoallergenic isoforms of the major birch pollen allergen Bet v 1 and its likely biological function as a plant steroid carrier. J. Mol. Biol 2003, 325, 123–133. [Google Scholar]
- Genoud, T.; Metraux, J.P. Crosstalk in plant cell signaling: Structure and function of the genetic network. Trends Plant Sci 1999, 4, 503–507. [Google Scholar]
- Xiong, L.; Yang, Y. Disease resistance and abiotic stress tolerance in rice are inversely modulated by an abscisic acid-inducible mitogen-activated protein kinase. Plant Cell 2003, 15, 745–759. [Google Scholar]
- Van Loon, L.C.; van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol 1999, 55, 85–97. [Google Scholar]
- Patterson, M.K. Measurements of growth and viability of cells in culture. Methods Enzymol 1979, 58, 141–152. [Google Scholar]
- Bochner, B.R.; Savageau, M.A. Generalized indicator plate for genetic, metabolic, and taxonomic studies with microorganisms. Appl. Environ. Microbiol 1977, 33, 434–444. [Google Scholar]
- Bochner, B.R.; Gadzinski, P.; Panomitros, E. Phenotype MicroArrays for high throughput phenotypic testing and assay of gene function. Genome Res 2001, 11, 1246–1255. [Google Scholar]
- Bochner, B.R. Global phenotypic characterization of bacteria. FEMS Microbiol. Rev 2009, 33, 191–205. [Google Scholar]
- Nachlas, M.M.; Margulies, S.I.; Seligman, A.M. Sites of electron transfer to tetrazolium salts in the succinoxidase system. J. Biol. Chem 1960, 235, 2739–2743. [Google Scholar]
- Duncan, D.R.; Widholm, J.M. Measurement of Viability Suitable for Plant Tissue Cultures. In Methods in Molecular Biology—Plant Cell and Tissue Culture, 2nd ed.; Polland, J.W., Walker, J.M., Eds.; The Humana Press: Totowa, NJ, USA, 1990; Volume 6, pp. 29–37. [Google Scholar]
- Zapata, J.M.; Salinas, C.; Calderon, A.A.; Munoz, R.; Barcelo, A.R. Reduction of 2,3,5-triphenyltetrazolium chloride by the KCN-insensitive, salicylhydroxamic acid-sensitive alternative respiratory pathway of mitochondria from cultured grapevine cells. Plant Cell Rep 1991, 10, 579–582. [Google Scholar]
- Kurzbaum, E.; Kirzhner, F.; Armon, R. A simple method for dehydrogenase activity visualization of intact plant roots grown in soilless cultures using tetrazolium violet. Plant Root 2010, 4, 12–16. [Google Scholar]
- Ruf, M.; Brunner, J. Vitality of tree fine roots: reevaluation of the tetrazolium test. Tree Physiol 2003, 23, 257–263. [Google Scholar]
- Duncan, D.R.; Widholm, J.M. Osmotic induced stimulation of the reduction of the viability dye 2,3,5-triphenyltetrazolium chloride by maize roots and callus cultures. J. Plant Physiol 2004, 161, 397–403. [Google Scholar]
- Hamer, L.; Adachi, K.; Montenegro-Chamorro, M.V.; Tanzer, M.M.; Mahanty, S.K.; Lo, C.; Tarpey, R.W.; Skalchunes, A.R.; Heiniger, R.W.; Frank, S.A.; et al. Gene discovery and gene function assignment in filamentous fungi. Proc. Natl. Acad. Sci. USA 2001, 98, 5110–5115. [Google Scholar]
- Bochner, B.R.; Siri, M.; Huang, R.H.; Noble, S.; Lei, X.H.; Clemons, P.A.; Wagner, B.K. Assay of the multiple energy-producing pathways of mammalian cells. PLoS One 2011, 6, e18147. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 2001, 29, e45. [Google Scholar]
- Lutts, S.; Almansouri, M.; Kinet, J.M. Salinity and water stress have contrasting effects on the relationship between growth and cell viability during and after stress exposure in durum wheat callus. Plant Sci 2004, 167, 9–18. [Google Scholar]
- Gopal, J.; Iwamain, K. In vitro screening of potato against water-stress mediated through sorbitol and polyethylene glycol. Plant Cell Rep 2007, 26, 693–700. [Google Scholar]
- Hohl, M.; Schopfer, P. Water relations of growing maize coleoptiles. Comparison between mannitol and polyethylene glycol 6000 as external osmotica for adjusting turgor pressure. Plant Physiol 1991, 95, 716–722. [Google Scholar]
- Gu, Y.Q.; Yang, C.; Thara, V.K.; Zhou, J.; Martin, G.B. Pti4 is induced by ethylene and salicylic acid, and its product is phosphorylated by the Pto kinase. Plant Cell 2000, 12, 771–786. [Google Scholar]
- Mysore, K.S.; Crasta, O.R.; Tuori, R.P.; Folkers, O.; Swirsky, P.B.; Martin, G.B. Comprehensive transcript profiling of Pto and Prf-mediated host defence responses to infection by Pseudomonas syringae pv. tomato. Plant J 2002, 32, 299–315. [Google Scholar]
- Wu, K.; Tian, L.; Hollingworth, J.; Brown, D.C.; Miki, B. Functional analysis of tomato Pti4 in Arabidopsis. Plant Physiol 2002, 128, 30–37. [Google Scholar]
- Matton, D.P.; Prescott, O.; Bertrand, C.; Camirand, A.; Brisson, N. ldentification of cis-acting elements involved in the regulation of the pathogenesis-related gene STH-2 in potato. Plant Mol. Biol 1993, 22, 279–291. [Google Scholar]
- Langridge, W.H.; Fitzgerald, K.J.; Koncz, C.; Schell, J.; Szalay, A.A. Dual promoter of Agrobacterium tumefaciens mannopine synthase genes is regulated by plant growth hormones. Proc. Natl. Acad. Sci. USA 1989, 86, 3219–3223. [Google Scholar]
- Fox, P.C.; Vasil, V.; Vasil, I.K.; Gurley, W.B. Multiple ocs-like elements required for efficient transcription of the mannopine synthase gene of T-DNA in maize protoplasts. Plant Mol. Biol 1992, 20, 219–233. [Google Scholar]
- Ali, Z.; Schumacher, H.M.; Heine-Dobbernack, E.; El-Banna, A.; Hafeez, F.Y.; Jacobsen, H.-J.; Kiesecker, H. Dicistronic binary vector system—A versatile tool for gene expression studies in cell cultures and plants. J. Biotechnol 2010, 145, 9–16. [Google Scholar]
- Gu, Y.Q.; Wildermuth, M.C.; Chakravarthy, S.; Loh, Y.T.; Yang, C.; He, X.; Han, Y.; Martin, G.B. Tomato transcription factors Pti4, Pti5 and Pti6 activate defence responses when expressed in Arabidopsis. Plant Cell 2002, 14, 817–831. [Google Scholar]
- Ulanov, A.; Widholm, J.M. Metabolic profiling to determine the cause of the increased triphenyltetrazolium chloride reduction in mannitol-treated maize callus. J. Plant Physiol 2010, 167, 1423–1431. [Google Scholar]
- Nilsen, E.; Orcutt, D.M. The Physiology of Plants under Stress—Abiotic Factors; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- Jacoby, R.P.; Taylor, N.L.; Millar, A.H. The role of mitochondrial respiration in salinity tolerance. Trends Plant Sci 2011, 16, 10. [Google Scholar]
- Millar, A.H.; Whelan, J.; Soole, K.L.; Day, D.A. Organization and regulation of mitochondrial respiration in plants. Annu. Rev. Plant Biol 2011, 62, 79–104. [Google Scholar]
- Galle, A.; Florez-Sarasal, I.; Thameur, A.; de Peape, R.; Flexas, J.; Ribas-Carbo, M. Effects of drought stress and subsequent rewatering on photosynthetic and respiratory pathways in Nicotiana sylvestris wild type and the mitochondrial complex I-deficient CMSII mutant. J. Exp. Bot 2010, 61, 765–775. [Google Scholar]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res 1968, 50, 151–158. [Google Scholar]
- Nicot, N.; Hausman, J.F.; Hoffmann, L.; Evers, D. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot 2005, 56, 2907–2914. [Google Scholar]
- The R Development Core Team. R: A language and environment for statistical computing. Available online: http://www.lsw.uni-heidelberg.de/users/christlieb/teaching/UKStaSS10/R-refman.pdf (on accessed 27 May 2013).
- Sarkar, D. Lattice: Multivariate Data Visualization with R, 1st ed.; Springer: New York, NY, USA, 2008. [Google Scholar]
- Schaarschmidt, F.; Vaas, L.A.I. Analysis of trials with complex treatment structure using multiple contrast tests. HortScience 2009, 44, 188–195. [Google Scholar]
- Chapin, F.S. Integrated responses of plants to stress. A centralized system of physiological responses. BioScience 1991, 40, 29–31. [Google Scholar]
- Brisbin, I.L.; Collins, C.T.; White, G.C.; McCallum, D.A. A new paradigm for the analysis and interpretation of growth data: the shape of things to come. Auk 1987, 104, 434–444. [Google Scholar]
- Vaas, L.A.I.; Sikorski, J.; Michael, V.; Göker, M.; Klenk, H.-P. Visualization and curve-parameter estimation strategies for efficient exploration of phenotype microarray kinetics. PLoS One 2012, 7, e34846. [Google Scholar]
- Tools for analyzing OmniLog (R) Phenotype Microarray data. Available online: http://cran.r-project.org/web/packages/opm/index.html (on accessed 19 July 2013).


© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Vaas, L.A.I.; Marheine, M.; Sikorski, J.; Göker, M.; Schumacher, H.-M. Impacts of pr-10a Overexpression at the Molecular and the Phenotypic Level. Int. J. Mol. Sci. 2013, 14, 15141-15166. https://doi.org/10.3390/ijms140715141
Vaas LAI, Marheine M, Sikorski J, Göker M, Schumacher H-M. Impacts of pr-10a Overexpression at the Molecular and the Phenotypic Level. International Journal of Molecular Sciences. 2013; 14(7):15141-15166. https://doi.org/10.3390/ijms140715141
Chicago/Turabian StyleVaas, Lea A. I., Maja Marheine, Johannes Sikorski, Markus Göker, and Heinz-Martin Schumacher. 2013. "Impacts of pr-10a Overexpression at the Molecular and the Phenotypic Level" International Journal of Molecular Sciences 14, no. 7: 15141-15166. https://doi.org/10.3390/ijms140715141