Calcium Signaling and Meiotic Exit at Fertilization in Xenopus Egg

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Xenopus Eggs before Fertilization

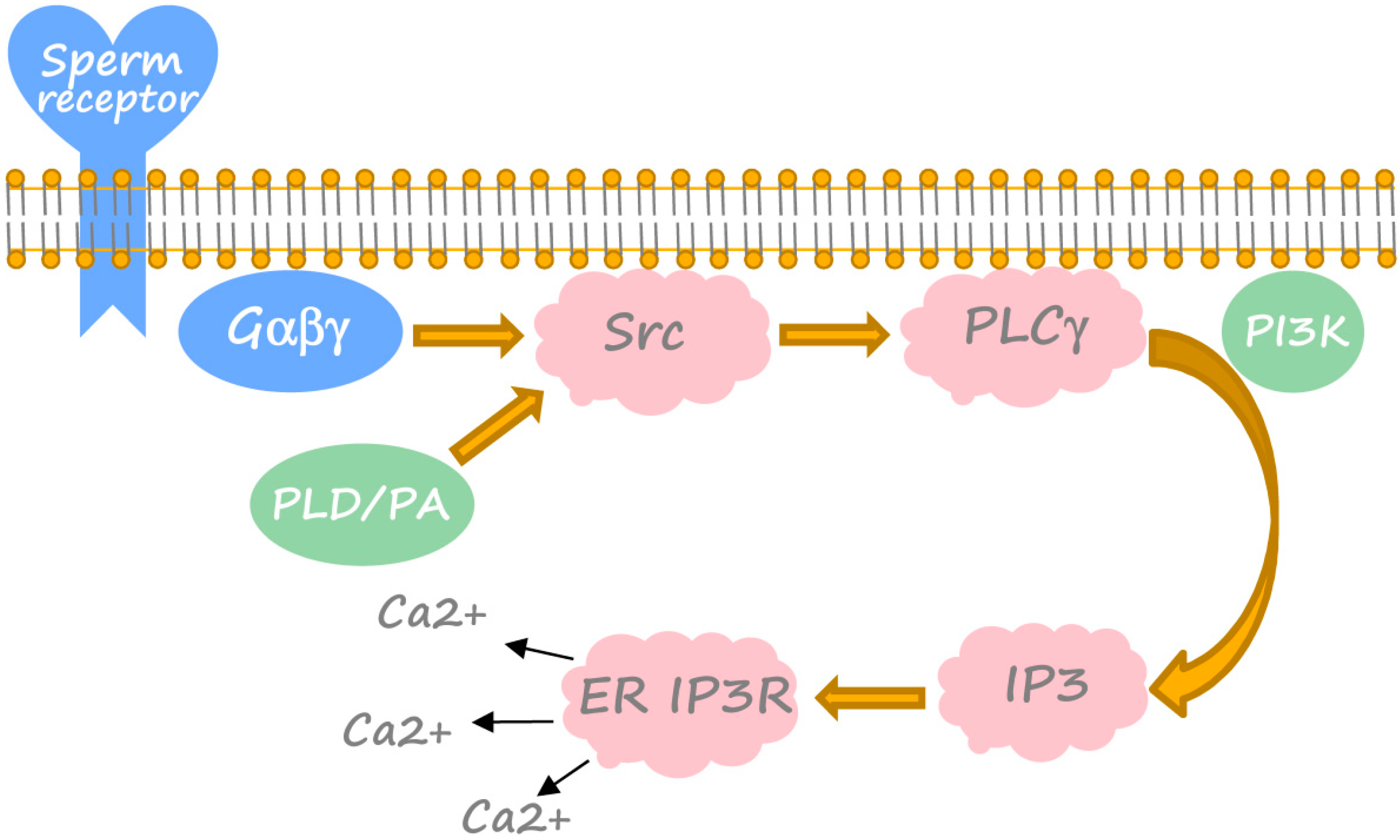
3. Generation of Calcium Transient in Xenopus Egg at Fertilization

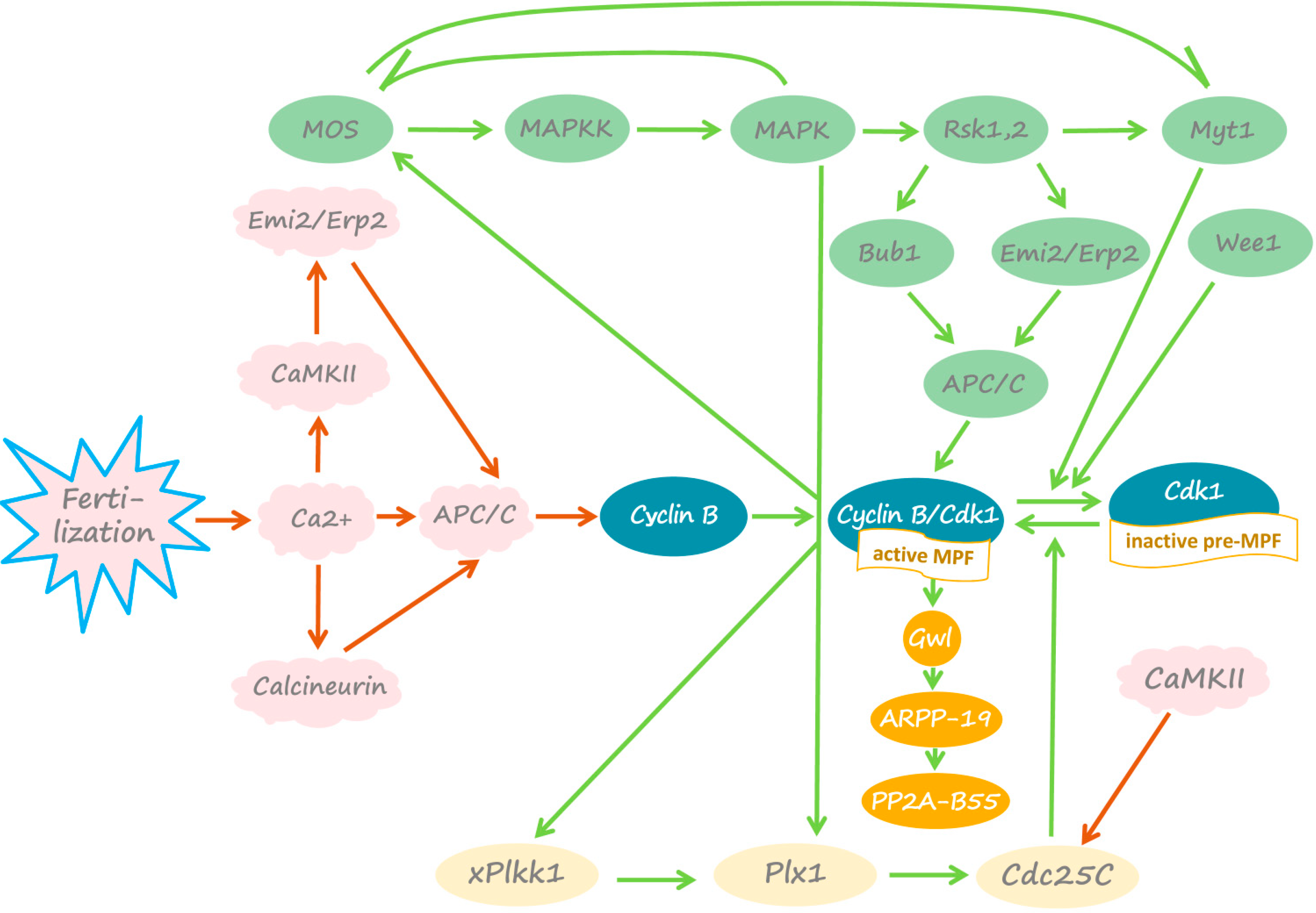
4. Fertilization-Induced Meiotic Exit
5. Other Effects of Calcium in Fertilized Xenopus Egg
6. Conclusions and Perspectives
Acknowledgments
Conflicts of Interest
References
- Masui, Y.; Markert, C.L. Cytoplasmic control of nuclear behavior during meiotic maturation of frog oocytes. J. Exp. Zool. 1971, 177, 129–145. [Google Scholar] [CrossRef]
- Mueller, P.R.; Coleman, T.R.; Kumagai, A.; Dunphy, W.G. Myt1: A membrane-associated inhibitory kinase that phosphorylates Ccd2 on both threonine-14 and tyrosine-15. Science 1995, 270, 86–90. [Google Scholar] [CrossRef]
- Murakami, M.S.; vande Woude, G.F. Analysis of the early embryonic cell cycles of Xenopus; regulation of cell cycle length by Xe-wee1 and Mos. Development 1998, 125, 237–248. [Google Scholar]
- Nakajo, N.; Yoshitome, S.; Iwashita, J.; Iida, M.; Uto, K.; Ueno, S.; Okamoto, K.; Sagata, N. Absence of Wee1 ensures the meiotic cell cycle in Xenopus oocytes. Genes Dev. 2000, 14, 328–338. [Google Scholar]
- Duckworth, B.C.; Weaver, J.S.; Ruderman, J.V. G2 arrest in Xenopus oocytes depends on phosphorylation of cdc25 by protein kinase A. Proc. Natl. Acad. Sci. USA 2002, 99, 16794–16799. [Google Scholar] [CrossRef]
- Sagata, N.; Watanabe, N.; vande Woude, G.F.; Ikawa, Y. The c-mos proto-oncogene product is a cytostatic factor responsible for meiotic arrest in vertebrate eggs. Nature 1989, 342, 512–518. [Google Scholar] [CrossRef]
- Gotoh, Y.; Masuyama, N.; Dell, K.; Shirakabe, K.; Nishida, E. Initiation of Xenopus oocyte maturation by activation of the mitogen-activated protein kinase cascade. J. Biol. Chem. 1995, 270, 25898–25904. [Google Scholar] [CrossRef]
- Gotoh, Y.; Nishida, E. The MAP kinase cascade: Its role in Xenopus oocytes, eggs and embryos. Prog. Cell Cycle Res. 1995, 1, 287–297. [Google Scholar]
- Haccard, O.; Lewellyn, A.; Hartley, R.S.; Erikson, E.; Maller, J.L. Induction of Xenopus oocyte meiotic maturation by MAP kinase. Dev. Biol. 1995, 168, 677–682. [Google Scholar] [CrossRef]
- Castro, A.; Peter, M.; Magnaghi-Jaulin, L.; Vigneron, S.; Galas, S.; Lorca, T.; Labbé, J.C. Cyclin B/cdc2 induces c-Mos stability by direct phosphorylation in Xenopus oocytes. Mol. Biol. Cell. 2001, 12, 2660–2671. [Google Scholar] [CrossRef]
- Howard, E.L.; Charlesworth, A.; Welk, J.; MacNicol, A.M. The mitogen-activated protein kinase signaling pathway stimulates mos mRNA cytoplasmic polyadenylation during Xenopus oocyte maturation. Mol. Cell. Biol. 1999, 19, 1990–1999. [Google Scholar]
- Palmer, A.; Gavin, A.C.; Nebreda, A.R. A link between MAP kinase and p34(cdc2)/cyclin B during oocyte maturation: p90(rsk) phosphorylates and inactivates the p34(cdc2) inhibitory kinase Myt1. EMBO J. 1998, 17, 5037–5047. [Google Scholar] [CrossRef]
- Mueller, P.R.; Coleman, T.R.; Dunphy, W.G. Cell cycle regulation of a Xenopus wee1-like kinase. Mol. Biol. Cell. 1995, 6, 119–134. [Google Scholar] [CrossRef]
- Inoue, D.; Ohe, M.; Kanemori, Y.; Nobui, T.; Sagata, N. A direct link of the Mos–MAPK pathway to Erp1/Emi2 in meiotic arrest of Xenopus laevis eggs. Nature 2007, 446, 1100–1104. [Google Scholar]
- Nishiyama, T.; Ohsumi, K.; Kishimoto, T. Phosphorylation of Erp1 by p90rsk is required for cytostatic factor arrest in Xenopus laevis eggs. Nature 2007, 446, 1096–1099. [Google Scholar] [CrossRef]
- Schwab, M.S.; Roberts, B.T.; Gross, S.D.; Tunquist, B.J.; Taieb, F.E.; Lewellyn, A.L.; Maller, J.L. Bub1 is activated by the protein kinase p90 (Rsk) during Xenopus oocyte maturation. Curr. Biol. 2001, 11, 141–150. [Google Scholar] [CrossRef]
- Tung, J.J.; Padmanabhan, K.; Hansen, D.V.; Richter, J.D.; Jackson, P.K. Translational unmasking of Emi2 directs cytostatic factor arrest in meiosis II. Cell Cycle 2007, 6, 725–731. [Google Scholar] [CrossRef]
- Schmidt, A.; Duncan, P.I.; Rauh, N.R.; Sauer, G.; Fry, A.M.; Nigg, E.A.; Mayer, T.U. Xenopus polo-like kinase Plx1 regulates XErp1, a novel inhibitor of APC/C activity. Genes Dev. 2005, 19, 502–513. [Google Scholar] [CrossRef]
- Tung, J.J.; Hansen, D.V.; Ban, K.H.; Loktev, A.V.; Summers, M.K.; Adler, J.R., 3rd; Jackson, P.K. A role for the anaphase-promoting complex inhibitor Emi2/XErp1, a homolog of early mitotic inhibitor 1, in cytostatic factor arrest of Xenopus eggs. Proc. Natl. Acad. Sci. USA 2005, 102, 4318–4323. [Google Scholar] [CrossRef]
- Shoji, S.; Yoshida, N.; Amanai, M.; Ohgishi, M.; Fukui, T.; Fujimoto, S.; Nakano, Y.; Kajikawa, E.; Perry, A.C. Mammalian Emi2 mediates cytostatic arrest and transduces the signal for meiotic exit via Cdc20. EMBO J. 2006, 25, 834–845. [Google Scholar] [CrossRef]
- Abrieu, A.; Brassac, T.; Galas, S.; Fisher, D.; Labbé, J.C.; Dorée, M. The Polo-like kinase Plx1 is a component of the MPF amplification loop at the G2/M-phase transition of the cell cycle in Xenopus eggs. J. Cell Sci. 1998, 111, 1751–1757. [Google Scholar]
- Gavin, A.C.; Ni Ainle, A.; Chierici, E.; Jones, M.; Nebreda, A.R. A p90(rsk) mutant constitutively interacting with MAP kinase uncouples MAP kinase from p34(cdc2)/cyclin B activation in Xenopus oocytes. Mol. Biol. Cell 1999, 10, 2971–2986. [Google Scholar] [CrossRef]
- Kumagai, A.; Dunphy, W.G. Purification and molecular cloning of Plx1, a Cdc25-regulatory kinase from Xenopus egg extracts. Science 1996, 273, 1377–1380. [Google Scholar]
- Qian, Y.W.; Erikson, E.; Maller, J.L. Purification and cloning of a protein kinase that phosphorylates and activates the polo-like kinase Plx1. Science 1998, 282, 1701–1704. [Google Scholar] [CrossRef]
- De Moor, C.H.; Richter, J.D. The Mos pathway regulates cytoplasmic polyadenylation in Xenopus oocytes. Mol. Cell. Biol. 1997, 17, 6419–6426. [Google Scholar]
- Matten, W.T.; Copeland, T.D.; Ahn, N.G.; Vande Woude, G.F. Positive feedback between MAP kinase and Mos during Xenopus oocyte maturation. Dev. Biol. 1996, 179, 485–492. [Google Scholar] [CrossRef]
- Mochida, S.; Ikeo, S.; Gannon, J.; Hunt, T. Regulated activity of PP2A-B55 delta is crucial for controlling entry into and exit from mitosis in Xenopus egg extracts. EMBO J. 2009, 28, 2777–2785. [Google Scholar] [CrossRef]
- Yu, J.; Zhao, Y.; Li, Z.; Galas, S.; Goldberg, M.L. Greatwall kinase participates in the Cdc2 autoregulatory loop in Xenopus egg extracts. Mol. Cell. 2006, 22, 83–91. [Google Scholar] [CrossRef]
- Castilho, P.V.; Williams, B.C.; Mochida, S.; Zhao, Y.; Goldberg, M.L. The M phase kinase Greatwall (Gwl) promotes inactivation of PP2A/B55delta, a phosphatase directed against CDK phosphosites. Mol. Biol. Cell. 2009, 20, 4777–4789. [Google Scholar] [CrossRef]
- Vigneron, S.; Brioudes, E.; Burgess, A.; Labbé, J.C.; Lorca, T.; Castro, A. Greatwall maintains mitosis through regulation of PP2A. EMBO J. 2009, 28, 2786–2793. [Google Scholar] [CrossRef]
- Mochida, S.; Maslen, S.L.; Skehel, M.; Hunt, T. Greatwall phosphorylates an inhibitor of protein phosphatase 2A that is essential for mitosis. Science 2010, 330, 1670–1673. [Google Scholar] [CrossRef]
- Gharbi-Ayachi, A.; Labbé, J.C.; Burgess, A.; Vigneron, S.; Strub, J.M.; Brioudes, E.; Van-Dorsselaer, A.; Castro, A.; Lorca, T. The substrate of Greatwall kinase, Arpp19, controls mitosis by inhibiting protein phosphatase 2A. Science 2010, 330, 1673–1677. [Google Scholar] [CrossRef]
- Hara, M.; Abe, Y.; Tanaka, T.; Yamamoto, T.; Okumura, E.; Kishimoto, T. Greatwall kinase and cyclin B-Cdk1 are both critical constituents of M-phase-promoting factor. Nat. Commun. 2012, 3, 1059. [Google Scholar] [CrossRef]
- Dupré, A.; Buffin, E.; Roustan, C.; Nairn, A.C.; Jessus, C.; Haccard, O. The phosphorylation of ARPP19 by Greatwall renders the auto-amplification of MPF independently of PKA in Xenopus oocytes. J. Cell Sci. 2013, 126, 3916–3926. [Google Scholar] [CrossRef]
- Whitaker, M. Control of meiotic arrest. Rev. Reprod. 1996, 1, 127–135. [Google Scholar] [CrossRef]
- Stricker, S.A. Comparative biology of calcium signaling during fertilization and egg activation in animals. Dev. Biol. 1999, 211, 157–176. [Google Scholar] [CrossRef]
- Saunders, C.M.; Larman, M.G.; Parrington, J.; Cox, L.J.; Royse, J.; Blayney, L.M.; Swann, K.; Lai, F.A. PLCζ: A sperm-specific trigger of Ca2+ oscillations in eggs and embryo development. Development 2002, 129, 3533–3544. [Google Scholar]
- Swann, K.; Saunders, C.M.; Rogers, N.T.; Lai, F.A. PLCζ: A sperm protein that triggers Ca2+ oscillations and egg activation in mammals. Semin. Cell Dev. Biol. 2006, 17, 264–273. [Google Scholar] [CrossRef]
- Swann, K.; Lai, F.A. PLCζ and the initiation of Ca2+ oscillations in fertilizing mammalian eggs. Cell Calcium 2013, 53, 55–62. [Google Scholar] [CrossRef]
- McGinnis, L.K.; Carroll, D.J.; Kinsey, W.H. Protein tyrosine kinase signaling during oocyte maturation and fertilization. Mol. Reprod. Dev. 2011, 78, 831–845. [Google Scholar]
- Kinsey, W.H. Intersecting roles of protein tyrosine kinase and calcium signaling during fertilization. Cell Calcium 2013, 53, 32–40. [Google Scholar] [CrossRef]
- Busa, W.B.; Nuccitelli, R. An elevated free cytosolic Ca2+ wave follows fertilization in eggs of the frog, Xenopus laevis. J. Cell Biol. 1985, 100, 1325–1329. [Google Scholar] [CrossRef]
- Nuccitelli, R.; Yim, D.L.; Smart, T. The sperm-induced Ca2+ wave following fertilization of the Xenopus egg requires the production of Ins(1,4,5)P3. Dev. Biol. 1993, 158, 200–212. [Google Scholar] [CrossRef]
- Sakakibara, K.; Sato, K.; Yoshino, K.; Oshiro, N.; Hirahara, S.; Mahbub Hasan, A.K.; Iwasaki, T.; Ueda, Y.; Iwao, Y.; Yonezawa, K.; et al. Molecular identification and characterization of Xenopus egg uroplakin III, an egg raft-associated transmembrane protein that is tyrosine-phosphorylated upon fertilization. J. Biol. Chem. 2005, 280, 15029–15037. [Google Scholar] [CrossRef]
- Mahbub Hasan, A.K.; Hashimoto, A.; Maekawa, Y.; Matsumoto, T.; Kushima, S.; Ijiri, T.W.; Fukami, Y.; Sato, K. The egg membrane microdomain-associated uroplakin III-Src system becomes functional during oocyte maturation and is required for bidirectional gamete signaling at fertilization in Xenopus laevis. Development 2014, 141, 1705–1714. [Google Scholar] [CrossRef]
- Kline, D.; Kopf, G.S.; Muncy, L.F.; Jaffe, L.A. Evidence for the involvement of a pertussis toxin-insensitive G-protein in egg activation of the frog, Xenopus laevis. Dev. Biol. 1991, 143, 218–229. [Google Scholar] [CrossRef]
- Jaffe, L.A. First messengers at fertilization. J. Reprod. Fertil. Suppl. 1990, 42, 107–116. [Google Scholar]
- Runft, L.L.; Watras, J.; Jaffe, L.A. Calcium release at fertilization of Xenopus eggs requires type I IP(3) receptors, but not SH2 domain-mediated activation of PLCgamma or G(q)-mediated activation of PLCβ. Dev. Biol. 1999, 214, 399–411. [Google Scholar] [CrossRef]
- Hepler, J.R.; Kozasa, T.; Smrcka, A.V.; Simon, M.I.; Rhee, S.G.; Sternweis, P.C.; Gilman, A.G. Purification from Sf9 cells and characterization of recombinant Gqα and G11α: Activation of purified phospholipase C isozymes by Gα subunits. J. Biol. Chem. 1993, 268, 14367–14375. [Google Scholar]
- Singer, W.D.; Miller, R.T.; Sternweis, P.C. Purification and characterization of the α subunit of G13. J. Biol. Chem. 1994, 269, 19796–19802. [Google Scholar]
- Kozasa, T.; Gilman, A.G. Purification of recombinant G proteins from Sf9 cells by hexahistidine tagging of associated subunits: Characterization of α12 and inhibition of adenylyl cyclase by αz. J. Biol. Chem. 1995, 270, 1734–1741. [Google Scholar] [CrossRef]
- Ciapa, B.; Chiri, S. Egg activation: Upstream of the fertilization calcium signal. Biol. Cell 2000, 92, 215–233. [Google Scholar] [CrossRef]
- Sato, K.; Tokmakov, A.A.; Fukami, Y. Fertilization signalling and protein-tyrosine kinases. Comp. Biochem. Physiol. B 2000, 126, 129–148. [Google Scholar] [CrossRef]
- Sato, K.; Fukami, Y.; Stith, B.J. Signal transduction pathways leading to Ca2+ release in a vertebrate model system: Lessons from Xenopus eggs. Semin. Cell Dev. Biol. 2006, 17, 285–292. [Google Scholar] [CrossRef]
- Tokmakov, A.A.; Iwasaki, T.; Sato, K.; Fukami, Y. Analysis of signal transduction in cell-free extracts and rafts of Xenopus eggs. Methods 2010, 51, 177–182. [Google Scholar] [CrossRef]
- Sato, K.; Iwao, Y.; Fujimura, T.; Tamaki, I.; Ogawa, K.; Iwasaki, T.; Tokmakov, A.A.; Hatano, O.; Fukami, Y. Evidence for the involvement of a Src-related tyrosine kinase in Xenopus egg activation. Dev. Biol. 1999, 209, 308–320. [Google Scholar] [CrossRef]
- Sato, K.; Tokmakov, A.A.; Iwasaki, T.; Fukami, Y. Tyrosine kinase-dependent activation of phospholipase Cgamma is required for calcium transient in Xenopus egg fertilization. Dev. Biol. 2000, 224, 453–469. [Google Scholar] [CrossRef]
- Tokmakov, A.A.; Sato, K.I.; Iwasaki, T.; Fukami, Y. Src kinase induces calcium release in Xenopus egg extracts via PLCγ and IP3-dependent mechanism. Cell Calcium 2002, 32, 11–20. [Google Scholar] [CrossRef]
- Mammadova, G.; Iwasaki, T.; Tokmakov, A.A.; Fukami, Y.; Sato, K. Evidence that phosphatidylinositol 3-kinase is involved in sperm-induced tyrosine kinase signaling in Xenopus egg fertilization. BMC Dev. Biol. 2009, 9, 68. [Google Scholar]
- Tokmakov, A.; Iwasaki, T.; Itakura, S.; Sato, K.; Shirouzu, M.; Fukami, Y.; Yokoyama, S. Regulation of Src kinase activity during Xenopus oocyte maturation. Dev. Biol. 2005, 278, 289–300. [Google Scholar] [CrossRef]
- Sato, K.; Aoto, M.; Mori, K.; Akasofu, S.; Tokmakov, A.A.; Sahara, S.; Fukami, Y. Purification and characterization of a Src-related p57 protein–tyrosine kinase from Xenopus oocytes. Isolation of an inactive form of the enzyme and its activation and translocation upon fertilization. J. Biol. Chem. 1996, 271, 13250–13257. [Google Scholar] [CrossRef]
- Sato, K.; Tokmakov, A.A.; He, C.L.; Kurokawa, M.; Iwasaki, T.; Shirouzu, M.; Fissore, R.A.; Yokoyama, S.; Fukami, Y. Reconstitution of Src-dependent phospholipase Cγ phosphorylation and transient calcium release by using membrane rafts and cell-free extracts from Xenopus eggs. J. Biol. Chem. 2003, 278, 38413–38420. [Google Scholar] [CrossRef]
- Mehlmann, L.M; Carpenter, G.; Rhee, S.G.; Jaffe, L.A. SH2 domain-mediated activation of phospholipase Cgamma is not required to initiate Ca2+ release at fertilization of mouse eggs. Dev. Biol. 1998, 203, 221–232. [Google Scholar] [CrossRef]
- Stith, B.J.; Goalstone, M.; Silva, S.; Jaynes, C. Inositol 1,4,5-trisphosphate mass changes from fertilization through first cleavage in Xenopus laevis. Mol. Biol. Cell 199, 4, 435–443. [Google Scholar]
- Dupont, G.; Goldbeter, A. Properties of intracellular Ca2+ waves generated by a model based on Ca2+-induced Ca2+ release. Biophys. J. 1994, 67, 2191–2204. [Google Scholar] [CrossRef]
- Wagner, J.; Li, Y.X.; Pearson, J.; Keizer, J. Simulation of the fertilization Ca2+ wave in Xenopus laevis eggs. Biophys. J. 1998, 75, 2088–2097. [Google Scholar] [CrossRef]
- Wagner, J.; Fall, C.P.; Hong, F.; Sims, C.E.; Allbritton, N.L.; Fontanilla, R.A.; Moraru, I.I.; Loew, L.M.; Nuccitelli, R. A wave of IP3 production accompanies the fertilization Ca2+ wave in the egg of the frog, Xenopus laevis: Theoretical and experimental support. Cell Calcium 2004, 35, 433–447. [Google Scholar] [CrossRef]
- Machaca, K.; Haun, S. Store-operated calcium entry inactivates at the germinal vesicle breakdown stage of Xenopus meiosis. J. Biol. Chem. 2000, 275, 38710–38715. [Google Scholar] [CrossRef]
- El-Jouni, W.; Jang, B.; Haun, S.; Machaca, K. Calcium signaling differentiation during Xenopus oocyte maturation. Dev. Biol. 2005, 288, 514–525. [Google Scholar] [CrossRef]
- Yu, F.; Sun, L.; Machaca, K. Orai1 internalization and STIM1 clustering inhibition modulate SOCE inactivation during meiosis. Proc. Natl. Acad. Sci. USA 2009, 106, 17401–17406. [Google Scholar] [CrossRef]
- Sun, L.; Yu, F.; Ullah, A.; Hubrack, S.; Daalis, A.; Jung, P.; Machaca, K. Endoplasmic reticulum remodeling tunes IP3-dependent Ca2+ release sensitivity. PLoS One 2011, 6, e27928. [Google Scholar]
- Ullah, A.; Jung, P.; Ullah, G.; Machaca, K. The role of IP3 receptor channel clustering in Ca2+ wave propagation during oocyte maturation. Prog. Mol. Biol. Transl. Sci. 2014, 123, 83–101. [Google Scholar]
- Stith, B.J.; Woronoff, K.; Espinoza, R.; Smart, T. sn-1,2-Diacylglycerol and choline increase after fertilization in Xenopus laevis. Mol. Biol. Cell 1997, 8, 755–765. [Google Scholar] [CrossRef]
- Bates, R.C.; Fees, C.P.; Holland, W.L.; Winger, C.C.; Batbayar, K.; Ancar, R.; Bergren, T.; Petcoff, D.; Stith, B.J. Activation of Src and release of intracellular calcium by phosphatidic acid during Xenopus laevis fertilization. Dev. Biol. 2014, 386, 165–180. [Google Scholar] [CrossRef]
- Watanabe, N.; Hunt, T.; Ikawa, Y.; Sagata, N. Independent inactivation of MPF and cytostatic factor (Mos) upon fertilization of Xenopus eggs. Nature 1991, 352, 247–248. [Google Scholar] [CrossRef]
- Lorca, T.; Cruzalegui, F.H.; Fesquet, D.; Cavadore, J.C.; Méry, J.; Means, A.; Dorée, M. Calmodulin-dependent protein kinase II mediates inactivation of MPF and CSF upon fertilization of Xenopus eggs. Nature 1993, 366, 270–273. [Google Scholar] [CrossRef]
- Hansen, D.V.; Tung, J.J.; Jackson, P.K. CaMKII and polo-like kinase 1 sequentially phosphorylate the cytostatic factor Emi2/XErp1 to trigger its destruction and meiotic exit. Proc. Natl. Acad. Sci. USA 2006, 103, 608–613. [Google Scholar] [CrossRef]
- Rauh, N.R.; Schmidt, A.; Bormann, J.; Nigg, E.A.; Mayer, T.U. Calcium triggers exit from meiosis II by targeting the APC/C inhibitor XErp1 for degradation. Nature 2005, 437, 1048–1052. [Google Scholar] [CrossRef]
- Schmidt, A.; Rauh, N.R.; Nigg, E.A.; Mayer, T.U. Cytostatic factor: An activity that puts the cell cycle on hold. J. Cell Sci. 2006, 119, 1213–1218. [Google Scholar] [CrossRef]
- Wu, J.Q.; Kornbluth, S. Across the meiotic divide-CSF activity in the post-Emi2/XErp1 era. J. Cell Sci. 2008, 121, 3509–3514. [Google Scholar] [CrossRef]
- Liu, J.; Maller, J.L. Calcium elevation at fertilization coordinates phosphorylation of XErp1/Emi2 by Plx1 and CaMK II to release metaphase arrest by cytostatic factor. Curr. Biol. 2005, 15, 1458–1468. [Google Scholar] [CrossRef]
- Nishiyama, A.; Tachibana, K.; Igarashi, Y.; Yasuda, H.; Tanahashi, N.; Tanaka, K.; Ohsumi, K.; Kishimoto, T. A nonproteolytic function of the proteasome is required for the dissociation of Cdc2 and cyclin B at the end of M phase. Genes Dev. 2000, 14, 2344–2357. [Google Scholar] [CrossRef]
- Hutchins, J.R.; Dikovskaya, D.; Clarke, P.R. Regulation of Cdc2/cyclin B activation in Xenopus egg extracts via inhibitory phosphorylation of Cdc25C phosphatase by Ca2+/calmodulin-dependent protein [corrected] kinase II. Mol. Biol. Cell. 2003, 14, 4003–4014. [Google Scholar] [CrossRef]
- Mochida, S.; Hunt, T. Calcineurin is required to release Xenopus egg extracts from meiotic M phase. Nature 2007, 449, 336–340. [Google Scholar] [CrossRef]
- Nishiyama, T.; Yoshizaki, N.; Kishimoto, T.; Ohsumi, K. Transient activation of calcineurin is essential to initiate embryonic development in Xenopus laevis. Nature 2007, 449, 341–345. [Google Scholar] [CrossRef]
- Chung, E.; Chen, R.H. Phosphorylation of Cdc20 is required for its inhibition by the spindle checkpoint. Nat. Cell Biol. 2003, 5, 748–753. [Google Scholar] [CrossRef]
- Hunt, T. On the regulation of protein phosphatase 2A and its role in controlling entry into and exit from mitosis. Adv. Biol. Regul. 2013, 53, 173–178. [Google Scholar] [CrossRef]
- Nishizawa, M.; Furuno, N.; Okazaki, K.; Tanaka, H.; Ogawa, Y.; Sagata, N. Degradation of Mos by the N-terminal proline (Pro2)-dependent ubiquitin pathway on fertilization of Xenopus eggs: Possible significance of natural selection for Pro2 in Mos. EMBO J. 1993, 12, 4021–4027. [Google Scholar]
- Paillard, L.; Omilli, F.; Legagneux, V.; Bassez, T.; Maniey, D.; Osborne, H.B. EDEN and EDEN-BP, a cis element and an associated factor that mediate sequence-specific mRNA deadenylation in Xenopus embryos. EMBO J. 1998, 17, 278–287. [Google Scholar] [CrossRef]
- Ueno, S.; Sagata, N. Requirement for both EDEN and AUUUA motifs in translational arrest of Mos mRNA upon fertilization of Xenopus eggs. Dev. Biol. 2002, 250, 156–167. [Google Scholar] [CrossRef]
- Watanabe, N.; Vande Woude, G.F.; Ikawa, Y.; Sagata, N. Specific proteolysis of the c-mos proto-oncogene product by calpain on fertilization of Xenopus eggs. Nature 1989, 342, 505–511. [Google Scholar] [CrossRef]
- Lorca, T.; Galas, S.; Fesquet, D.; Devault, A.; Cavadore, J.C.; Dorée, M. Degradation of the proto-oncogene product p39mos is not necessary for cyclin proteolysis and exit from meiotic metaphase: Requirement for a Ca2+-calmodulin dependent event. EMBO J. 1991, 10, 2087–2093. [Google Scholar]
- Levasseur, M.; Dumollard, R.; Chambon, J.P.; Hebras, C.; Sinclair, M.; Whitaker, M.; McDougall, A. Release from meiotic arrest in ascidian eggs requires the activity of two phosphatases but not CaMKII. Development 2013, 140, 4583–4593. [Google Scholar] [CrossRef]
- Madgwick, S.; Levasseur, M.; Jones, K.T. Calmodulin-dependent protein kinase II, and not protein kinase C, is sufficient for triggering cell-cycle resumption in mammalian eggs. J. Cell Sci. 2005, 118, 3849–3859. [Google Scholar] [CrossRef]
- Suzuki, T.; Suzuki, E.; Yoshida, N.; Kubo, A.; Li, H.; Okuda, E.; Amanai, M.; Perry, A.C. Mouse Emi2 as a distinctive regulatory hub in second meiotic metaphase. Development 2010, 137, 3281–3291. [Google Scholar] [CrossRef]
- Kline, D. Calcium-dependent events at fertilization of the frog egg: Injection of a calcium buffer blocks ion channel opening, exocytosis, and formation of pronuclei. Dev. Biol. 1988, 126, 346–361. [Google Scholar] [CrossRef]
- Grandin, N.; Charbonneau, M. The increase in intracellular pH associated with Xenopus egg activation is a Ca2+-dependent wave. J. Cell Sci. 1992, 101, 55–67. [Google Scholar]
- Bement, W.M.; Capco, D.G. Protein kinase C acts downstream of calcium at entry into the first mitotic interphase of Xenopus laevis. Cell Regul. 1990, 1, 315–326. [Google Scholar]
- Stith, B.J.; Espinoza, R.; Roberts, D.; Smart, T. Sperm increase inositol 1,4,5-trisphosphate mass in Xenopus laevis eggs preinjected with calcium buffers or heparin. Dev. Biol. 1994, 165, 206–215. [Google Scholar] [CrossRef]
- Holloway, S.L.; Glotzer, M.; King, R.W.; Murray, A.W. Anaphase is initiated by proteolysis rather than by the inactivation of maturation-promoting factor. Cell 1993, 73, 1393–1402. [Google Scholar] [CrossRef]
- Morin, N.; Abrieu, A.; Lorca, T.; Martin, F.; Dorée, M. The proteolysis-dependent metaphase to anaphase transition: Calcium/calmodulin-dependent protein kinase II mediates onset of anaphase in extracts prepared from unfertilized Xenopus eggs. EMBO J. 1994, 13, 4343–4352. [Google Scholar]
- Larabell, C.A.; Rowning, B.A.; Moon, R.T. A PKC wave follows the calcium wave after activation of Xenopus eggs. Differentiation 2004, 72, 41–47. [Google Scholar] [CrossRef]
- Bement, W.M.; Capco, D.G. Activators of protein kinase C trigger cortical granule exocytosis, cortical contraction, and cleavage furrow formation in Xenopus laevis oocytes and eggs. J. Cell Biol. 1989, 108, 885–892. [Google Scholar] [CrossRef]
- Collas, P.; Thompson, L.; Fields, A.P.; Poccia, D.L.; Courvalin, J.C. Protein kinase C-mediated interphase lamin B phosphorylation and solubilization. J. Biol. Chem. 1997, 272, 21274–21280. [Google Scholar] [CrossRef]
- Exton, J.H. Phosphatidylcholine breakdown and signal transduction. Biochim. Biophys. Acta 1994, 1212, 26–42. [Google Scholar] [CrossRef]
- Barish, M.E. A transient calcium-dependent chloride current in the immature Xenopus oocyte. J. Physiol. 1983, 342, 309–325. [Google Scholar]
- Glahn, D.; Nuccitelli, R. Voltage-clamp study of the activation currents and fast block to polyspermy in the egg of Xenopus laevis. Dev. Growth Differ. 2003, 45, 187–197. [Google Scholar] [CrossRef]
- Grandin, N.; Charbonneau, M. Intracellular pH and intracellular free calcium responses to protein kinase C activators and inhibitors in Xenopus eggs. Development 1991, 112, 461–470. [Google Scholar]
- Rankin, S.; Kirschner, M.W. The surface contraction waves of Xenopus eggs reflect the metachronous cell-cycle state of the cytoplasm. Curr. Biol. 1997, 7, 451–454. [Google Scholar] [CrossRef]
- Pérez-Mongiovi, D.; Chang, P.; Houliston, E. A propagated wave of MPF activation accompanies surface contraction waves at first mitosis in Xenopus. J. Cell Sci. 1998, 111, 385–393. [Google Scholar]
- Pérez-Mongiovi, D.; Beckhelling, C.; Chang, P.; Ford, C.C.; Houliston, E. Nuclei and microtubule asters stimulate maturation/M phase promoting factor (MPF) activation in Xenopus eggs and egg cytoplasmic extracts. J. Cell Biol. 2000, 150, 963–974. [Google Scholar]
- Marrari, Y.; Clarke, E.J.; Rouvière, C.; Houliston, E. Analysis of microtubule movement on isolated Xenopus egg cortices provides evidence that the cortical rotation involves dynein as well as Kinesin Related Proteins and is regulated by local microtubule polymerisation. Dev. Biol. 2003, 257, 55–70. [Google Scholar] [CrossRef]
- Shiina, N.; Moriguchi, T.; Ohta, K.; Gotoh, Y.; Nishida, E. Regulation of a major microtubule-associated protein by MPF and MAP kinase. EMBO J. 1992, 11, 3977–3984. [Google Scholar]
- Andersen, S.S.; Buendia, B.; Domínguez, J.E.; Sawyer, A.; Karsenti, E. Effect on microtubule dynamics of XMAP230, a microtubule-associated protein present in Xenopus laevis eggs and dividing cells. J. Cell Biol. 1994, 127, 1289–1299. [Google Scholar] [CrossRef]
- Vasquez, R.J.; Gard, D.L.; Cassimeris, L. Phosphorylation by CDK1 regulates XMAP215 function in vitro. Cell Motil. Cytoskelet. 1999, 43, 310–321. [Google Scholar] [CrossRef]
- Lorca, T.; Castro, A.; Martinez, A.M.; Vigneron, S.; Morin, N.; Sigrist, S.; Lehner, C.; Dorée, M.; Labbé, J.C. Fizzy is required for activation of the APC/cyclosome in Xenopus egg extracts. EMBO J. 1998, 17, 3565–3575. [Google Scholar] [CrossRef]
- Peter, M.; Castro, A.; Lorca, T.; Le Peuch, C.; Magnaghi-Jaulin, L.; Dorée, M.; Labbé, J.C. The APC is dispensable for first meiotic anaphase in Xenopus oocytes. Nat. Cell Biol. 2001, 3, 83–87. [Google Scholar] [CrossRef]
- Ohta, Y.; Ohba, T.; Miyamoto, E. Ca2+/calmodulin-dependent protein kinase II: Localization in the interphase nucleus and the mitotic apparatus of mammalian cells. Proc. Natl. Acad. Sci. USA 1990, 87, 5341–5345. [Google Scholar] [CrossRef]
- Zou, H.; McGarry, T.J.; Bernal, T.; Kirschner, M.W. Identification of a vertebrate sister-chromatid separation inhibitor involved in transformation and tumorigenesis. Science 1999, 285, 418–422. [Google Scholar] [CrossRef]
- Stemmann, O.; Zou, H.; Gerber, S.A.; Gygi, S.P.; Kirschner, M.W. Dual inhibition of sister chromatid separation at metaphase. Cell 2001, 107, 715–726. [Google Scholar] [CrossRef]
- Waizenegger, I.C.; Hauf, S.; Meinke, A.; Peters, J.M. Two distinct pathways remove mammalian cohesin from chromosome arms in prophase and from centromeres in anaphase. Cell 2000, 103, 399–410. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokmakov, A.A.; Stefanov, V.E.; Iwasaki, T.; Sato, K.-I.; Fukami, Y. Calcium Signaling and Meiotic Exit at Fertilization in Xenopus Egg. Int. J. Mol. Sci. 2014, 15, 18659-18676. https://doi.org/10.3390/ijms151018659
Tokmakov AA, Stefanov VE, Iwasaki T, Sato K-I, Fukami Y. Calcium Signaling and Meiotic Exit at Fertilization in Xenopus Egg. International Journal of Molecular Sciences. 2014; 15(10):18659-18676. https://doi.org/10.3390/ijms151018659
Chicago/Turabian StyleTokmakov, Alexander A., Vasily E. Stefanov, Tetsushi Iwasaki, Ken-Ichi Sato, and Yasuo Fukami. 2014. "Calcium Signaling and Meiotic Exit at Fertilization in Xenopus Egg" International Journal of Molecular Sciences 15, no. 10: 18659-18676. https://doi.org/10.3390/ijms151018659