Emodin Ameliorates LPS-Induced Acute Lung Injury, Involving the Inactivation of NF-κB in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
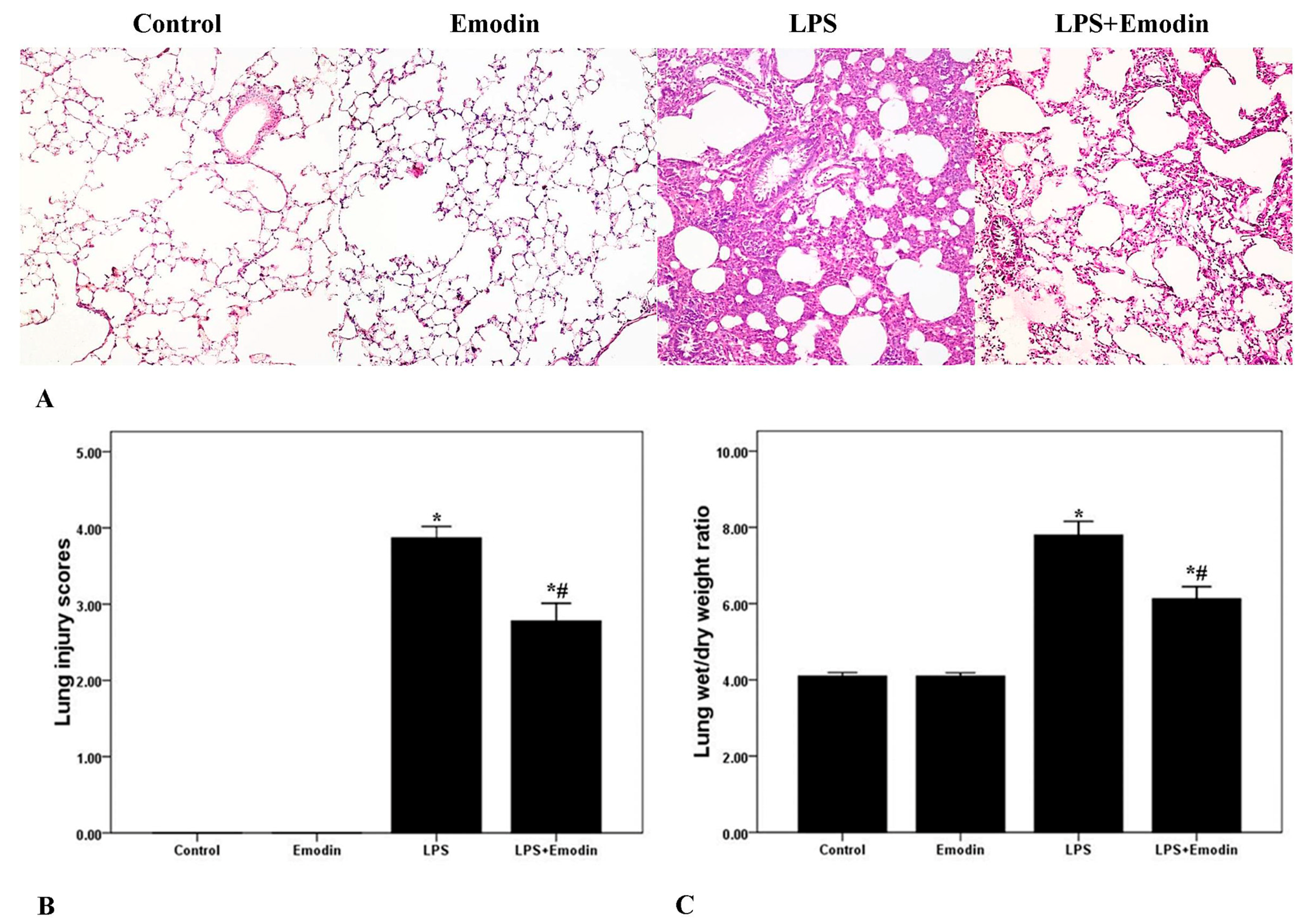
2.1. Emodin Ameliorates Pulmonary Inflammation and Pulmonary Edema in Lipopolysaccharide (LPS)-Induced Acute Lung Injury (ALI)

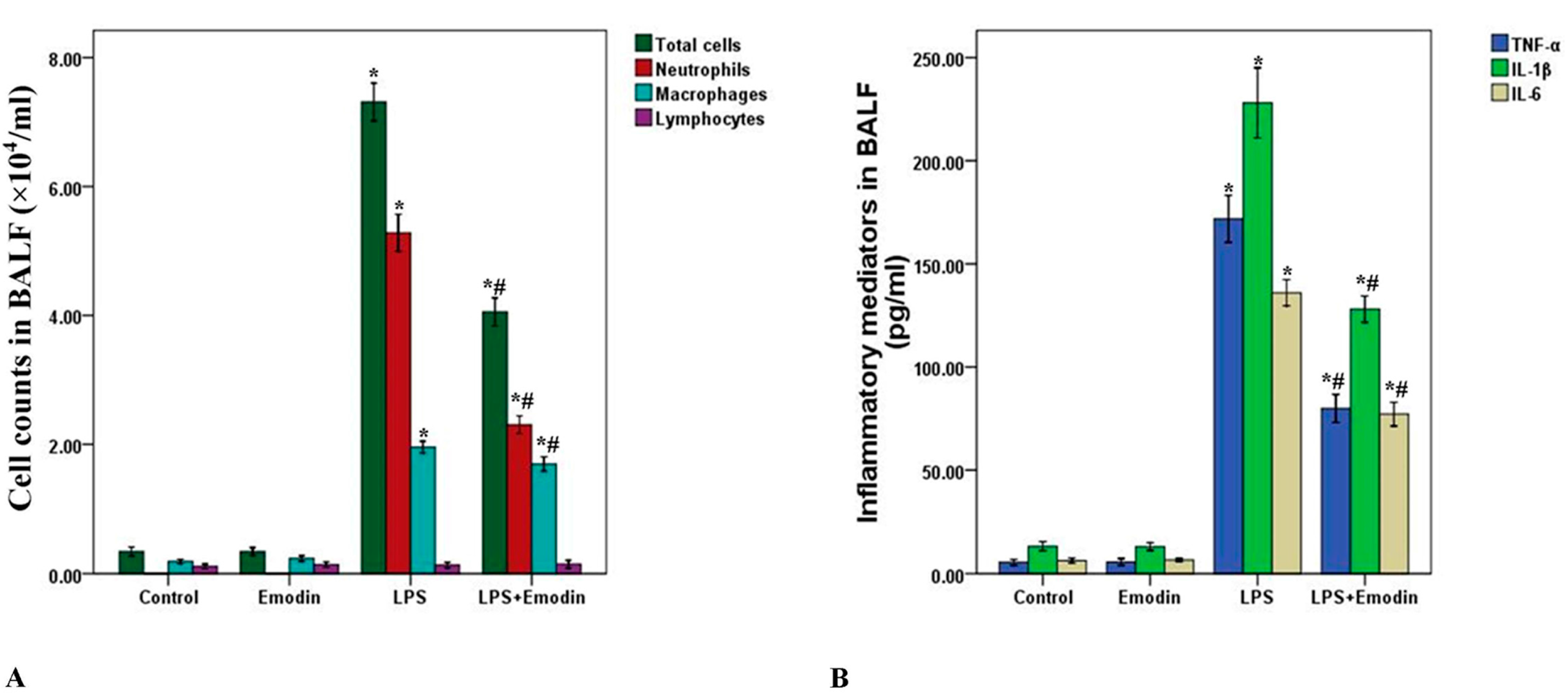
2.2. Emodin Decreases Cell Counts and Inflammatory Mediators in Bronchoalveolar Lavage Fluid (BALF) in LPS-Induced ALI

2.3. Emodin Inhibits Myeloperoxidase (MPO) Activity in LPS-Induced ALI
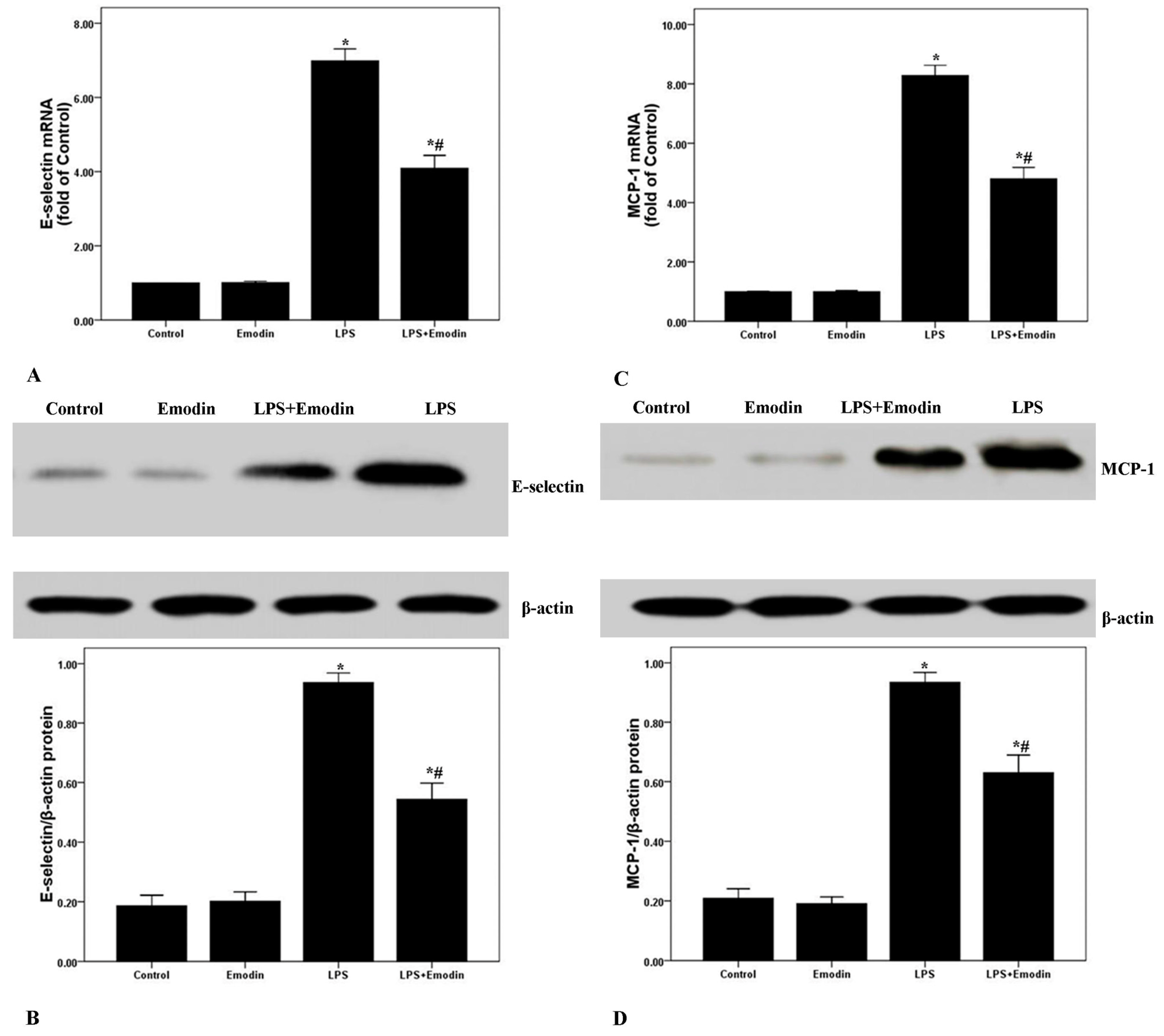
2.4. Emodin Reduces E-Selectin and MCP-1 Expression in Lung Tissues in LPS-Induced ALI


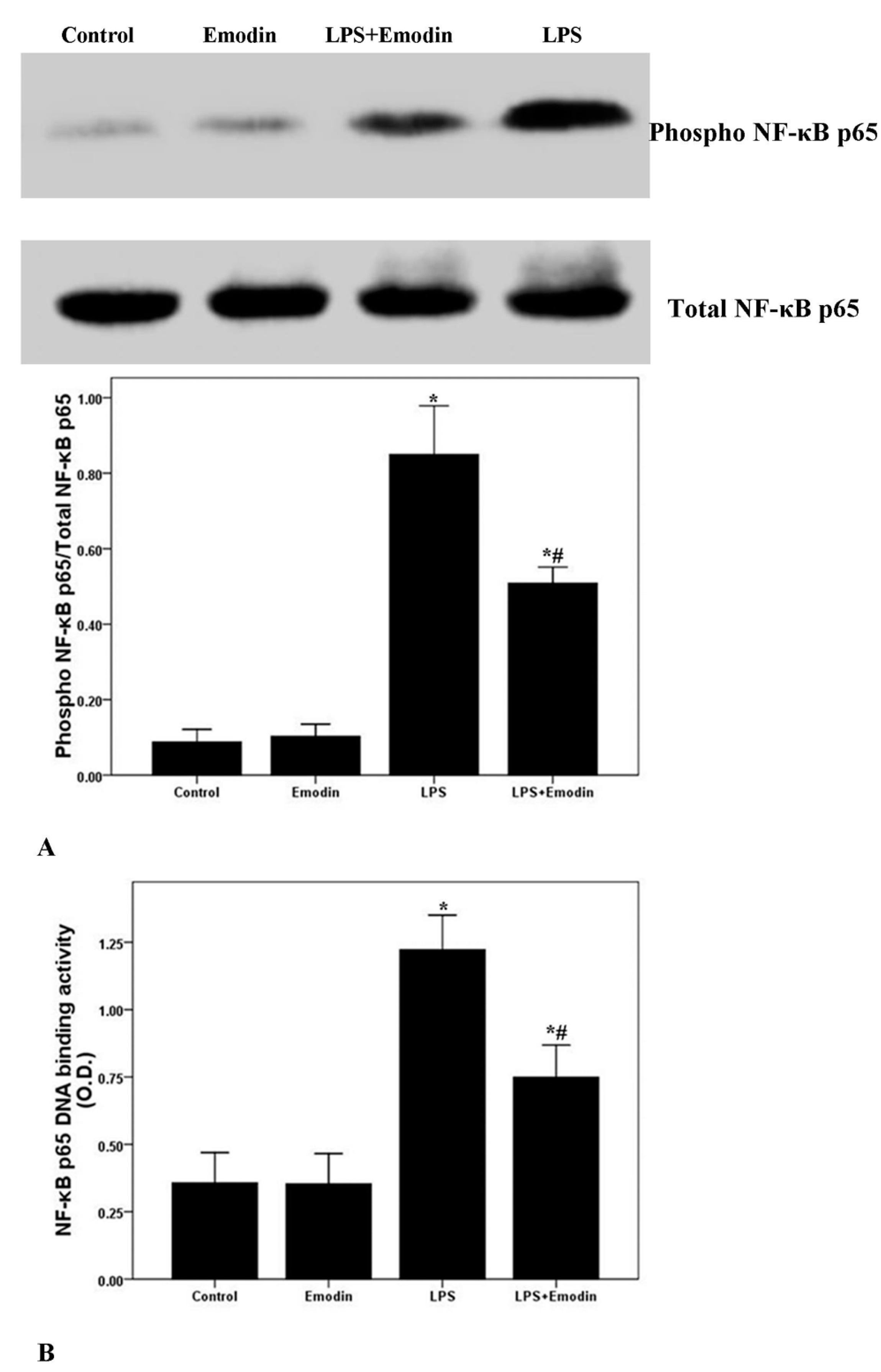
2.5. Emodin Inhibits LPS-Induced NF-κB Activation and DNA Binding Activity in Lung in LPS-Induced ALI

3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Murine Model of LPS-Induced ALI
4.3. Bronchoalveolar Lavage Fluid (BALF) and Cells Counting
4.4. TNF-α, IL-6 and IL-1β in BALF
4.5. Myeloperoxidase (MPO) Activity Assay
4.6. Lung Wet/Dry Weight Ratio
4.7. H&E Staining
4.8. Quantitative PCR
4.9. Western Blotting
4.10. NF-κB p65 DNA Binding Activity Assay
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gotts, J.E.; Matthay, M.A. Treating ARDS: New hope for a tough problem. Lancet Respir. Med. 2014, 2, 84–85. [Google Scholar]
- Ferguson, N.D.; Fan, E.; Camporota, L.; Antonelli, M.; Anzueto, A.; Beale, R.; Brochard, L.; Brower, R.; Esteban, A.; Gattinoni, L.; et al. The Berlin definition of ARDS: An expanded rationale, justification, and supplementary material. Intensive Care Med. 2012, 38, 1573–1582. [Google Scholar]
- Raghavendran, K.; Napolitano, L.M. ALI and ARDS: Challenges and advances. Crit. Care Clin. 2011, 27, 429–437. [Google Scholar]
- Frohlich, S.; Murphy, N.; Boylan, J.F. ARDS: Progress unlikely with non-biological definition. Br. J. Anaesth. 2013, 111, 696–699. [Google Scholar]
- Boyle, A.J.; Mac, S.R.; McAuley, D.F. Pharmacological treatments in ARDS; a state-of-the-art update. BMC Med. 2013, 11, 166. [Google Scholar]
- Janz, D.R.; Ware, L.B. Biomarkers of ALI/ARDS: Pathogenesis, discovery, and relevance to clinical trials. Semin. Respir. Crit. Care Med. 2013, 34, 537–548. [Google Scholar]
- Antonelli, M.; Bonten, M.; Chastre, J.; Citerio, G.; Conti, G.; Curtis, J.R.; de Backer, D.; Hedenstierna, G.; Joannidis, M.; Macrae, D.; et al. Year in review in Intensive Care Medicine 2011: III. ARDS and ECMO, weaning, mechanical ventilation, noninvasive ventilation, pediatrics and miscellanea. Intensive Care Med. 2012, 38, 542–556. [Google Scholar]
- Wise, M.P.; Hart, N.; Frost, P.J. ARDS outcomes: A marker of critical care quality in the UK? Thorax 2011, 66, 359. [Google Scholar]
- Zhu, Y.G.; Qu, J.M.; Zhang, J.; Jiang, H.N.; Xu, J.F. Novel interventional approaches for ALI/ARDS: Cell-based gene therapy. Mediat. Inflamm. 2011, 2011, 560194. [Google Scholar]
- Fernandez-Bustamante, A.; Repine, J.E. Chronic inflammatory diseases and the acute respiratory distress syndrome (ARDS). Curr. Pharm. Des. 2014, 20, 1400–1408. [Google Scholar]
- Ware, L.B. Autopsy in ARDS: Insights into natural history. Lancet Respir. Med. 2013, 1, 352–354. [Google Scholar]
- Thompson, B.T.; Matthay, M.A. The Berlin definition of ARDS versus pathological evidence of diffuse alveolar damage. Am. J. Respir. Crit. Care Med. 2013, 187, 675–677. [Google Scholar]
- De Luca, D.; Piastra, M.; Tosi, F.; Pulitano, S.; Mancino, A.; Genovese, O.; Pietrini, D.; Conti, G. Pharmacological therapies for pediatric and neonatal ALI/ARDS: An evidence-based review. Curr. Drug Targets 2012, 13, 906–916. [Google Scholar]
- Lin, X.; Dean, D.A. Gene therapy for ALI/ARDS. Crit. Care Clin. 2011, 27, 705–718. [Google Scholar]
- Zhu, T.; Wang, D.X.; Zhang, W.; Liao, X.Q.; Guan, X.; Bo, H.; Sun, J.Y.; Huang, N.W.; He, J.; Zhang, Y.K.; et al. Andrographolide protects against LPS-induced acute lung injury by inactivation of NF-κB. PLoS One 2013, 8, e56407. [Google Scholar]
- Zhu, T.; Zhang, W.; Wang, D.X. Insulin up-regulates epithelial sodium channel in LPS-induced acute lung injury model in rats by SGK1 activation. Injury 2012, 43, 1277–1283. [Google Scholar]
- Xiao, M.; Zhu, T.; Wang, T.; Wen, F.Q. Hydrogen-rich saline reduces airway remodeling via inactivation of NF-κB in a murine model of asthma. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 1033–1043. [Google Scholar]
- Zhu, T.; Zhang, W.; Xiao, M.; Chen, H.; Jin, H. Protective role of andrographolide in bleomycin-induced pulmonary fibrosis in mice. Int. J. Mol. Sci. 2013, 14, 23581–23596. [Google Scholar]
- Wang, X.; Zhang, L.; Duan, W.; Liu, B.; Gong, P.; Ding, Y.; Wu, X. Anti-inflammatory effects of triptolide by inhibiting the NF-κB signalling pathway in LPS-induced acute lung injury in a murine model. Mol. Med. Rep. 2014, 10, 447–452. [Google Scholar]
- Jin, L.Y.; Li, C.F.; Zhu, G.F.; Wu, C.T.; Wang, J.; Yan, S.F. Effect of siRNA against NF-κB on sepsisinduced acute lung injury in a mouse model. Mol. Med. Rep. 2014, 10, 631–637. [Google Scholar]
- Killeen, M.J.; Linder, M.; Pontoniere, P.; Crea, R. NF-κB signaling and chronic inflammatory diseases: Exploring the potential of natural products to drive new therapeutic opportunities. Drug Discov. Today 2014, 19, 373–378. [Google Scholar]
- Zhang, X.; Zhang, R.; Lv, P.; Yang, J.; Deng, Y.; Xu, J.; Zhu, R.; Zhang, D.; Yang, Y. Emodin up-regulates glucose metabolism, decreases lipolysis, and attenuates inflammation in vitro. J. Diabetes 2014. [Google Scholar] [CrossRef]
- Yang, Z.; Zhou, E.; Wei, D.; Li, D.; Wei, Z.; Zhang, W.; Zhang, X. Emodin inhibits LPS-induced inflammatory response by activating PPAR-γ in mouse mammary epithelial cells. Int. Immunopharmacol. 2014, 21, 354–360. [Google Scholar]
- Ni, Q.; Zhang, W.; Sun, K.; Yin, C.; An, J.; Shang, D. Effects of emodin on peritoneal macrophage intercellular adhesion molecule-3 in a rat model of severe acute pancreatitis/systemic inflammatory response syndrome. Biomed. Rep. 2014, 2, 63–68. [Google Scholar]
- Yang, J.; Zeng, Z.; Wu, T.; Yang, Z.; Liu, B.; Lan, T. Emodin attenuates high glucose-induced TGF-β1 and fibronectin expression in mesangial cells through inhibition of NF-κB pathway. Exp. Cell Res. 2013, 319, 3182–3189. [Google Scholar]
- Li, D.; Zhang, N.; Cao, Y.; Zhang, W.; Su, G.; Sun, Y.; Liu, Z.; Li, F.; Liang, D.; Liu, B.; et al. Emodin ameliorates lipopolysaccharide-induced mastitis in mice by inhibiting activation of NF-κB and MAPKs signal pathways. Eur. J. Pharmacol. 2013, 705, 79–85. [Google Scholar]
- Wu, L.; Cai, B.; Zheng, S.; Liu, X.; Cai, H.; Li, H. Effect of emodin on endoplasmic reticulum stress in rats with severe acute pancreatitis. Inflammation 2013, 36, 1020–1029. [Google Scholar]
- Chen, X.H.; Sun, R.S.; Hu, J.M.; Mo, Z.Y.; Yang, Z.F.; Jin, G.Y.; Guan, W.D.; Zhong, N.S. Inhibitory effect of emodin on bleomycin-induced pulmonary fibrosis in mice. Clin. Exp. Pharmacol. Physiol. 2009, 36, 146–153. [Google Scholar]
- Liao, Z.; Dong, J.; Wu, W.; Yang, T.; Wang, T.; Guo, L.; Chen, L.; Xu, D.; Wen, F. Resolvin D1 attenuates inflammation in lipopolysaccharide-induced acute lung injury through a process involving the PPARγ/NF-κB pathway. Respir. Res. 2012, 13, 110. [Google Scholar]
- Wang, X.W.; Wang, J.X. Pattern recognition receptors acting in innate immune system of shrimp against pathogen infections. Fish Shellfish Immunol. 2013, 34, 981–989. [Google Scholar]
- Van der Vaart, M.; Spaink, H.P.; Meijer, A.H. Pathogen recognition and activation of the innate immune response in zebrafish. Adv. Hematol. 2012, 2012, 159807. [Google Scholar]
- Rohmann, K.; Tschernig, T.; Pabst, R.; Goldmann, T.; Dromann, D. Innate immunity in the human lung: Pathogen recognition and lung disease. Cell Tissue Res. 2011, 343, 167–174. [Google Scholar]
- Kim, E.; Tolhurst, A.T.; Cho, S. Deregulation of inflammatory response in the diabetic condition is associated with increased ischemic brain injury. J. Neuroinflamm. 2014, 11, 83. [Google Scholar]
- Lv, W.; Lv, C.; Yu, S.; Yang, Y.; Kong, H.; Xie, J.; Sun, H.; Andersson, R.; Xu, D.; Chen, B.; et al. Lipoxin A4 attenuation of endothelial inflammation response mimicking pancreatitis-induced lung injury. Exp. Biol. Med. 2013, 238, 1388–1395. [Google Scholar]
- Xie, K.; Yu, Y.; Huang, Y.; Zheng, L.; Li, J.; Chen, H.; Han, H.; Hou, L.; Gong, G.; Wang, G.; et al. Molecular hydrogen ameliorates lipopolysaccharide-induced acute lung injury in mice through reducing inflammation and apoptosis. Shock 2012, 37, 548–555. [Google Scholar]
- Ueda, A.; Okuda, K.; Ohno, S.; Shirai, A.; Igarashi, T.; Matsunaga, K.; Fukushima, J.; Kawamoto, S.; Ishigatsubo, Y.; Okubo, T.; et al. NF-κB and Sp1 regulate transcription of the human monocyte chemoattractant protein-1 gene. J. Immunol. 1994, 153, 2052–2063. [Google Scholar]
- Fakhrudin, N.; Waltenberger, B.; Cabaravdic, M.; Atanasov, A.G.; Malainer, C.; Schachner, D.; Heiss, E.H.; Liu, R.; Noha, S.M.; Grzywacz, A.M.; et al. Identification of plumericin as a potent new inhibitor of the NF-κB pathway with anti-inflammatory activity in vitro and in vivo. Br. J. Pharmacol. 2014, 171, 1676–1686. [Google Scholar]
- Chen, L.; Zhao, Q.; Wang, X.L.; You, R.; Zhang, Y.H.; Ji, H.; Lai, Y.S. ZLJ-6, a novel COX/5-LOX inhibitor, attenuates TNF-α-induced endothelial E-selectin, ICAM-1 and VCAM-1 expression and monocyte-endothelial interactions via a COX/5-LOX-independent mechanism. Vasc. Pharmacol. 2011, 55, 135–142. [Google Scholar]
- Song, H.; Bergstrasser, C.; Rafat, N.; Hoger, S.; Schmidt, M.; Endres, N.; Goebeler, M.; Hillebrands, J.L.; Brigelius-Flohe, R.; Banning, A.; et al. The carbon monoxide releasing molecule (CORM-3) inhibits expression of vascular cell adhesion molecule-1 and E-selectin independently of haem oxygenase-1 expression. Br. J. Pharmacol. 2009, 157, 769–780. [Google Scholar]
- Zhu, T.; Zhang, W.; Wang, D.X.; Huang, N.W.; Bo, H.; Deng, W.; Deng, J. Rosuvastatin attenuates mucus secretion in a murine model of chronic asthma by inhibiting the γ-aminobutyric acid type A receptor. Chin. Med. J. 2012, 125, 1457–1464. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, M.; Zhu, T.; Zhang, W.; Wang, T.; Shen, Y.-C.; Wan, Q.-F.; Wen, F.-Q. Emodin Ameliorates LPS-Induced Acute Lung Injury, Involving the Inactivation of NF-κB in Mice. Int. J. Mol. Sci. 2014, 15, 19355-19368. https://doi.org/10.3390/ijms151119355
Xiao M, Zhu T, Zhang W, Wang T, Shen Y-C, Wan Q-F, Wen F-Q. Emodin Ameliorates LPS-Induced Acute Lung Injury, Involving the Inactivation of NF-κB in Mice. International Journal of Molecular Sciences. 2014; 15(11):19355-19368. https://doi.org/10.3390/ijms151119355
Chicago/Turabian StyleXiao, Min, Tao Zhu, Wei Zhang, Tao Wang, Yong-Chun Shen, Qiong-Fang Wan, and Fu-Qiang Wen. 2014. "Emodin Ameliorates LPS-Induced Acute Lung Injury, Involving the Inactivation of NF-κB in Mice" International Journal of Molecular Sciences 15, no. 11: 19355-19368. https://doi.org/10.3390/ijms151119355