Aberrant Expression of Xist in Aborted Porcine Fetuses Derived from Somatic Cell Nuclear Transfer Embryos



Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
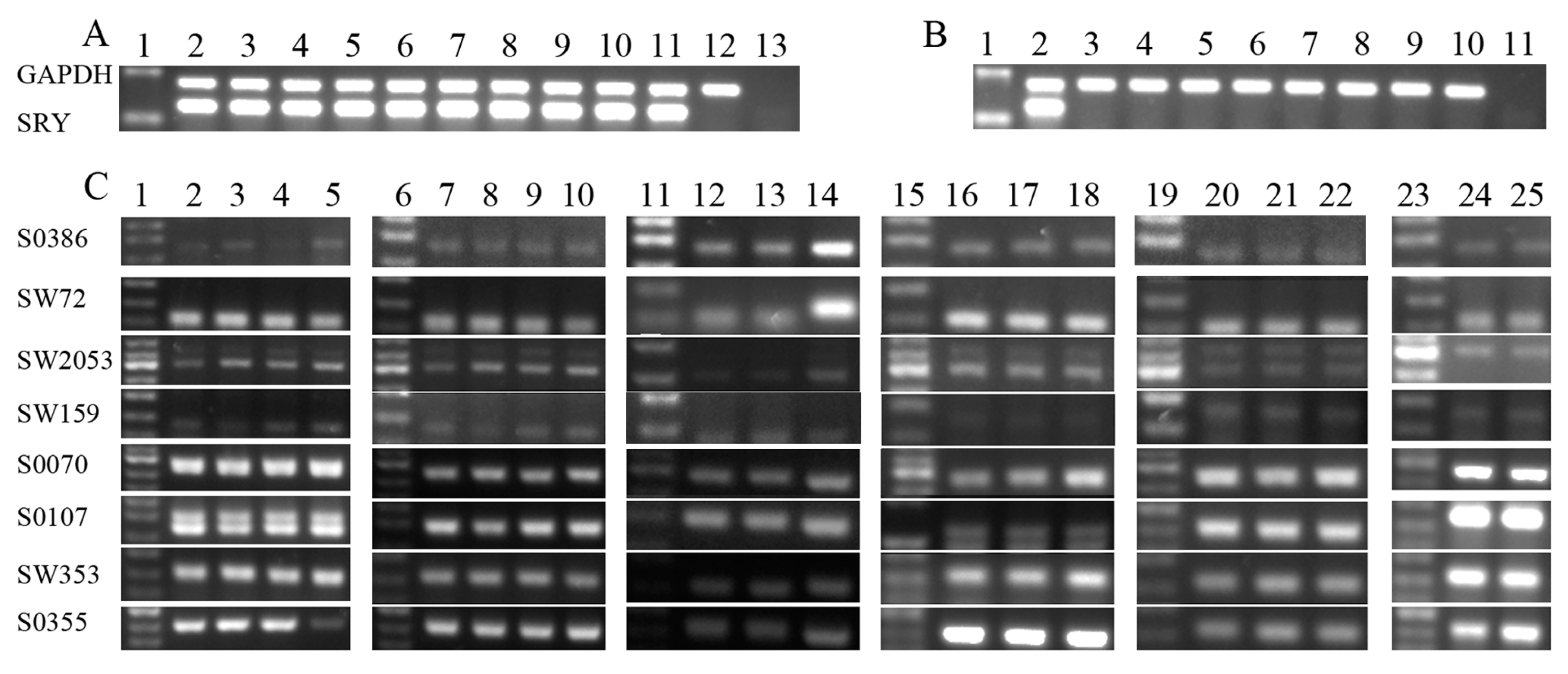
2.1.1. Sample Collection and Sex Identification of Aborted Fetuses

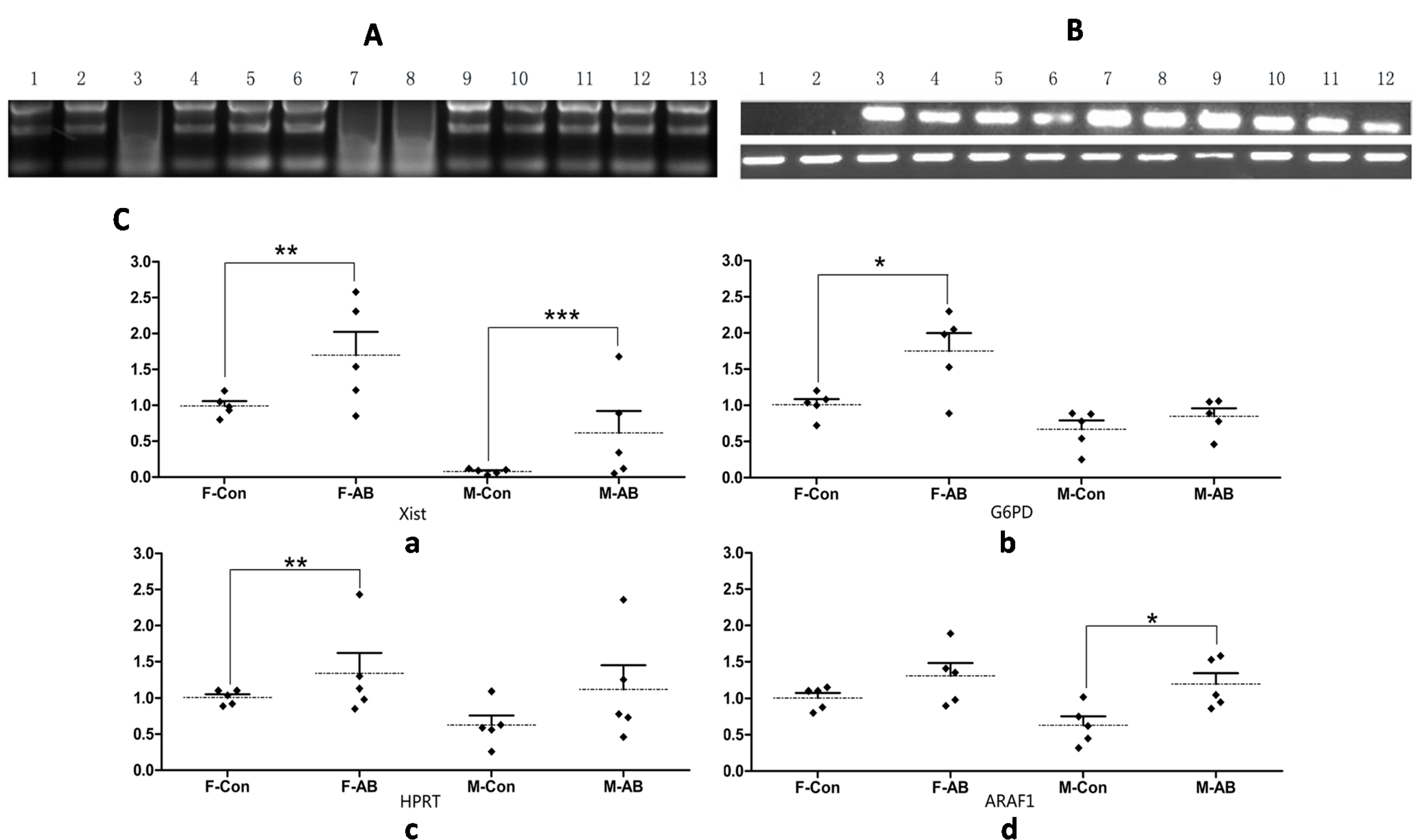
2.1.2. Transcription Patterns of X-Linked Genes in Aborted Fetuses

{kind=link}
{kind=link}
{kind=link}
| Groups | No. of Samples | Gender of the Donor Cell | SRY Amplification * | q-PCR & |
|---|---|---|---|---|
| 1 | 3 | Male | 3 | 2 |
| 2 | 3 | Male | 3 | 3 |
| 3 | 2 | Male | 1 | 0 |
| 4 | 2 | Female | 0 | 2 |
| 5 | 2 | Female | 0 | 2 |
| 6 | 1 | Female | 0 | 1 |

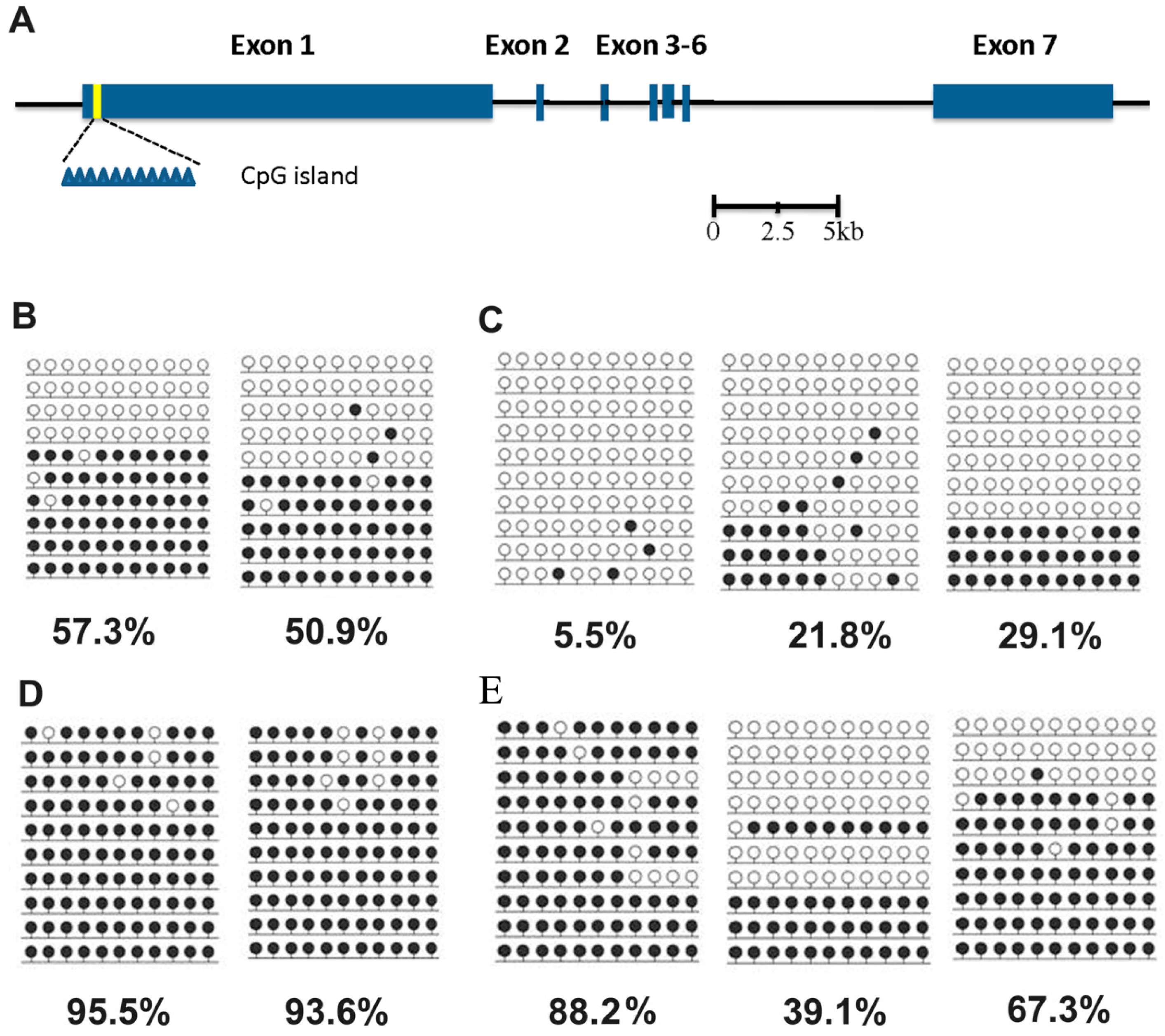
2.1.3. Methylation Status of the Xist Gene in Male and Female Aborted Fetuses

2.2. Discussion
3. Experimental Section
3.1. Ethics Statement
3.2. Samples Collection
3.3. Genotype Identification and Bisulfate-Specific PCR (BSP)
| Gene | Primer Sequence | Size (bp) |
|---|---|---|
| SRY | Sense: GCTTTCATTGTGTGGTCTCGT | 309 |
| Antisense: CTTGGCGACTGTGTATGTGAAG | ||
| GAPDH | Sense: GATGGCCCCTCTGGGAAACTGTG | 404 |
| Antisense: GGACGCCTGCTTCACCACCTTCT | ||
| Xist | Sense: GGATAATATGGTTGATTTTGTTATGTG | 212 |
| Antisense: CCACCACCCTTTCTAATTAAATATATC |
| Locus | Primer Sequence |
|---|---|
| S0386 | Sense: 5-GAACTCCTGGGTCTTATTTTCTA-3 |
| Antisense: 5-GTCAAAAATCTTTTTATCTCCAACAGTAT-3 | |
| SW72 | Sense: 5-ATCAGAACAGTGCGCCGT-3 |
| Antisense: 5-TTTGAAAATGGGGTGTTTCC-3 | |
| SW2053 | Sense: 5-AAGCAAGGTGCCACTGTTG-3 |
| Antisense: 5-CGAACCCGATGTCCTCTGAC-3 | |
| SW159 | Sense: 5-GATTGGGAATTTGGGGTT-3 |
| Antisense: 5-CGTCTTTACTTTTGTTGTTACG-3 | |
| S0070 | Sense: 5-GGCGAGCATTTCATTCACAG-3 |
| Antisense: 5-GAGCAAACAGCATCGTGAGC-3 | |
| S0107 | Sense: 5-CAAGGATGCCTGTAACTGGTGCAG-3 |
| Antisense: 5-TCCTTAAGGCCTCGTAGGATCTGT-5 | |
| SW353 | Sense: 5-CACCCCATGCCTGAATACTG-3 |
| Antisense: 5-ATGTGAAGACTCATGCTTGGG-3 | |
| S0355 | Sense: 5-TCTGGCTCCTACACTCCTTCTTGATG-3 |
| Antisense: 5-GTTTGGGTGGGTGCTGAAAAATAGGA-3 |
| Gene | Primer Sequence | Size (bp) |
|---|---|---|
| Xist outside | Sense: GTGTGTATTTTTTGATAAATTTTGT | 330 |
| Antisense: CTATACTAACTAACTAAATAAAAAC | ||
| Xist inside | Sense: GGATAATATGGTTGATTTTGTTATGTG | 212 |
| Antisense: CACCACCCTTTCTAATTAAATATATC |
3.4. Gene Expression Analysis
| Gene | Primer Sequence | Size (bp) |
|---|---|---|
| Xist | Sense: GAAGAGATGCTCCAGGCCAAT | 87 |
| Antisense: AGGTGTTGCTGGCTGATGCT | ||
| G6PD | Sense: CCTCCTGCAGATGCTGTGTCT | 112 |
| Antisense: CGCCTGCACCTCTGAGATG | ||
| HPRT1 | Sense: CGTCTTGCTCGAGATGTGATG | 98 |
| Antisense: TCCAGCAGGTCAGCAAAGAA | ||
| ARAF1 | Sense: CGGGATGGCATGAGTGTCTAC | 108 |
| Antisense: GACTGTCTTTCGCCCCTTGA | ||
| GAPDH | Sense: ATTCCACGGCACAGTCAAGG | 120 |
| Antisense: ACATACTCAGCACCAGCATCG |
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lai, L.; Kolber-Simonds, D.; Park, K.W.; Cheong, H.T.; Greenstein, J.L.; Im, G.S.; Samuel, M.; Bonk, A.; Rieke, A.; Day, B.N.; et al. Production of alpha-1,3-galactosyltransferase knockout pigs by nuclear transfer cloning. Science 2002, 295, 1089–1092. [Google Scholar]
- Suzuki, S.; Iwamoto, M.; Saito, Y.; Fuchimoto, D.; Sembon, S.; Suzuki, M.; Mikawa, S.; Hashimoto, M.; Aoki, Y.; Najima, Y.; et al. Il2rg Gene-targeted severe combined immunodeficiency pigs. Cell Stem Cell 2012, 10, 753–758. [Google Scholar]
- Park, K.W.; Cheong, H.T.; Lai, L.X.; Im, G.S.; Kuhholzer, B.; Bonk, A.; Samuel, M.; Rieke, A.; Day, B.N.; Murphy, C.N.; et al. Production of nuclear transfer-derived swine that express the enhanced green fluorescent protein. Anim. Biotechnol. 2001, 12, 173–181. [Google Scholar]
- Mao, J.; Tessanne, K.; Whitworth, K.M.; Spate, L.D.; Walters, E.M.; Samuel, M.S.; Murphy, C.N.; Tracy, L.; Zhao, J.; Prather, R.S. Effects of combined treatment of MG132 and scriptaid on early and term development of porcine somatic cell nuclear transfer embryos. Cell. Reprogram. 2012, 14, 385–389. [Google Scholar]
- Huang, Y.; Ouyang, H.; Yu, H.; Lai, L.; Pang, D.; Li, Z. Efficiency of porcine somatic cell nuclear transfer—A retrospective study of factors related to embryo recipient and embryos transferred. Biol. Open 2013, 2, 1223–1228. [Google Scholar]
- Wells, K.D. More with less Xist. Proc. Natl. Acad. Sci. USA 2012, 109, 349–350. [Google Scholar]
- Park, C.H.; Jeong, Y.H.; Jeong, Y.I.; Lee, S.Y.; Jeong, Y.W.; Shin, T.; Kim, N.H.; Jeung, E.B.; Hyun, S.H.; Lee, C.K.; et al. X-Linked gene transcription patterns in female and male in vivo, in vitro and cloned porcine individual blastocysts. PLoS One 2012, 7, e51398. [Google Scholar]
- Jiang, L.; Lai, L.; Samuel, M.; Prather, R.S.; Yang, X.; Tian, X.C. Expression of X-linked genes in deceased neonates and surviving cloned female piglets. Mol. Reprod. Dev. 2008, 75, 265–273. [Google Scholar]
- Zuccotti, M.; Boiani, M.; Ponce, R.; Guizzardi, S.; Scandroglio, R.; Garagna, S.; Redi, C.A. Mouse Xist expression begins at zygotic genome activation and is timed by a zygotic clock. Mol. Reprod. Dev. 2002, 61, 14–20. [Google Scholar]
- Nolen, L.D.; Gao, S.; Han, Z.; Mann, M.R.; Gie Chung, Y.; Otte, A.P.; Bartolomei, M.S.; Latham, K.E. X chromosome reactivation and regulation in cloned embryos. Dev. Biol. 2005, 279, 525–540. [Google Scholar]
- Oikawa, M.; Inoue, K.; Shiura, H.; Matoba, S.; Kamimura, S.; Hirose, M.; Mekada, K.; Yoshiki, A.; Tanaka, S.; Abe, K.; et al. Understanding the X chromosome inactivation cycle in mice. Epigenetics 2014, 9, 204–211. [Google Scholar]
- Inoue, K.; Kohda, T.; Sugimoto, M.; Sado, T.; Ogonuki, N.; Matoba, S.; Shiura, H.; Ikeda, R.; Mochida, K.; Fujii, T.; et al. Impeding Xist expression from the active X chromosome improves mouse somatic cell nuclear transfer. Science 2010, 330, 496–499. [Google Scholar]
- Matoba, S.; Inoue, K.; Kohda, T.; Sugimoto, M.; Mizutani, E.; Ogonuki, N.; Nakamura, T.; Abe, K.; Nakano, T.; Ishino, F.; et al. RNAi-mediated knockdown of Xist can rescue the impaired postimplantation development of cloned mouse embryos. Proc. Natl. Acad. Sci. USA 2011, 108, 20621–20626. [Google Scholar]
- Peippo, J.; Farazmand, A.; Kurkilahti, M.; Markkula, M.; Basrur, P.K.; King, W.A. Sex-chromosome linked gene expression in in vitro produced bovine embryos. Mol. Hum. Reprod. 2002, 8, 923–929. [Google Scholar]
- Brown, C.J.; Lafreniere, R.G.; Powers, V.E.; Sebastio, G.; Ballabio, A.; Pettigrew, A.L.; Ledbetter, D.H.; Levy, E.; Craig, I.W.; Willard, H.F. Localization of the X inactivation centre on the human X chromosome in Xq13. Nature 1991, 349, 82–84. [Google Scholar]
- Lee, J.E.; Beck, T.W.; Brennscheidt, U.; DeGennaro, L.J.; Rapp, U.R. The complete sequence and promoter activity of the human A-raf-1 gene (ARAF1). Genomics 1994, 20, 43–55. [Google Scholar]
- Yasue, H.; Adams, L.; Ozawa, A.; Hanazono, M.; Li, N.; Lin, Z.H.; Kusumoto, H. Assignment of ARAF1 to porcine chromosome Xp11.2-p13 by fluorescence in situ hybridization. Mamm. Genome 1997, 8, 457–458. [Google Scholar]
- Ogura, A.; Inoue, K.; Ogonuki, N.; Lee, J.; Kohda, T.; Ishino, F. Phenotypic effects of somatic cell cloning in the mouse. Cloning Stem Cells 2002, 4, 397–405. [Google Scholar]
- Meissner, A.; Jaenisch, R. Mammalian nuclear transfer. Dev. Dyn. 2006, 235, 2460–2469. [Google Scholar]
- Van Thuan, N.; Kishigam, S.; Wakayama, T. How to improve the success rate of mouse cloning technology. J. Reprod. Dev. 2010, 56, 20–30. [Google Scholar]
- Bang, J.I.; Lee, H.S.; Deb, G.K.; Ha, A.N.; Kwon, Y.S.; Cho, S.K.; Kim, B.W.; Cho, K.W.; Kong, I.K. Proteomic identification of abnormally expressed proteins in early-stage placenta derived from cloned cat embryos. Theriogenology 2013, 79, 358–366. [Google Scholar]
- Lyon, M.F. Gene action in the X-chromosome of the mouse (Mus musculus L.). Nature 1961, 190, 372–373. [Google Scholar]
- Lessing, D.; Anguera, M.C.; Lee, J.T. X chromosome inactivation and epigenetic responses to cellular reprogramming. Annu. Rev. Genomics Hum. Genet. 2013, 14, 85–110. [Google Scholar]
- Kalantry, S.; Purushothaman, S.; Bowen, R.B.; Starmer, J.; Magnuson, T. Evidence of Xist RNA-independent initiation of mouse imprinted X-chromosome inactivation. Nature 2009, 460, 647–651. [Google Scholar]
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H.S. Viable offspring derived from fetal and adult mammalian cells. Cloning Stem Cells 2007, 9, 3–7. [Google Scholar]
- Jeon, B.G.; Rho, G.J.; Betts, D.H.; Petrik, J.J.; Favetta, L.A.; King, W.A. Low levels of X-inactive specific transcript in somatic cell nuclear transfer embryos derived from female bovine freemartin donor Cells. Sex. Dev. 2012, 6, 151–159. [Google Scholar]
- Wei, Y.C.; Huan, Y.J.; Shi, Y.Q.; Liu, Z.F.; Bou, G.; Luo, Y.B.; Zhang, L.; Yang, C.R.; Kong, Q.R.; Tian, J.T.;et al. Unfaithful maintenance of methylation imprints due to loss of maternal nuclear dnmt1 during somatic cell nuclear transfer. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Li, E.; Bestor, T.H.; Jaenisch, R. Targeted mutation of the DNA methyltransferase gene results in embryonic lethality. Cell 1992, 69, 915–926. [Google Scholar]
- Coleman-Derr, D.; Zilberman, D. DNA methylation, H2A.Z, and the regulation of constitutive expression. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 147–154. [Google Scholar]
- Lai, L.X.; Prather, R.S. Production of cloned pigs by using somatic cells as donors. Cloning Stem Cells 2003, 5, 233–241. [Google Scholar]
- Xu, W.; Li, Z.; Yu, B.; He, X.; Shi, J.; Zhou, R.; Liu, D.; Wu, Z. Effects of DNMT1 and HDAC inhibitors on gene-specific methylation reprogramming during porcine somatic cell nuclear transfer. PLoS One 2013, 8, e64705. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, L.; Wang, A.; Yao, C.; Huang, Y.; Duan, F.; Lv, Q.; Wang, D.; Ouyang, H.; Li, Z.; Lai, L. Aberrant Expression of Xist in Aborted Porcine Fetuses Derived from Somatic Cell Nuclear Transfer Embryos. Int. J. Mol. Sci. 2014, 15, 21631-21643. https://doi.org/10.3390/ijms151221631
Yuan L, Wang A, Yao C, Huang Y, Duan F, Lv Q, Wang D, Ouyang H, Li Z, Lai L. Aberrant Expression of Xist in Aborted Porcine Fetuses Derived from Somatic Cell Nuclear Transfer Embryos. International Journal of Molecular Sciences. 2014; 15(12):21631-21643. https://doi.org/10.3390/ijms151221631
Chicago/Turabian StyleYuan, Lin, Anfeng Wang, Chaogang Yao, Yongye Huang, Feifei Duan, Qinyan Lv, Dongxu Wang, Hongsheng Ouyang, Zhanjun Li, and Liangxue Lai. 2014. "Aberrant Expression of Xist in Aborted Porcine Fetuses Derived from Somatic Cell Nuclear Transfer Embryos" International Journal of Molecular Sciences 15, no. 12: 21631-21643. https://doi.org/10.3390/ijms151221631
APA StyleYuan, L., Wang, A., Yao, C., Huang, Y., Duan, F., Lv, Q., Wang, D., Ouyang, H., Li, Z., & Lai, L. (2014). Aberrant Expression of Xist in Aborted Porcine Fetuses Derived from Somatic Cell Nuclear Transfer Embryos. International Journal of Molecular Sciences, 15(12), 21631-21643. https://doi.org/10.3390/ijms151221631