Nano-Scale Spatial Assessment of Calcium Distribution in Coccolithophores Using Synchrotron-Based Nano-CT and STXM-NEXAFS

 ,
,
Abstract
:1. Introduction
2. Results and Discussion

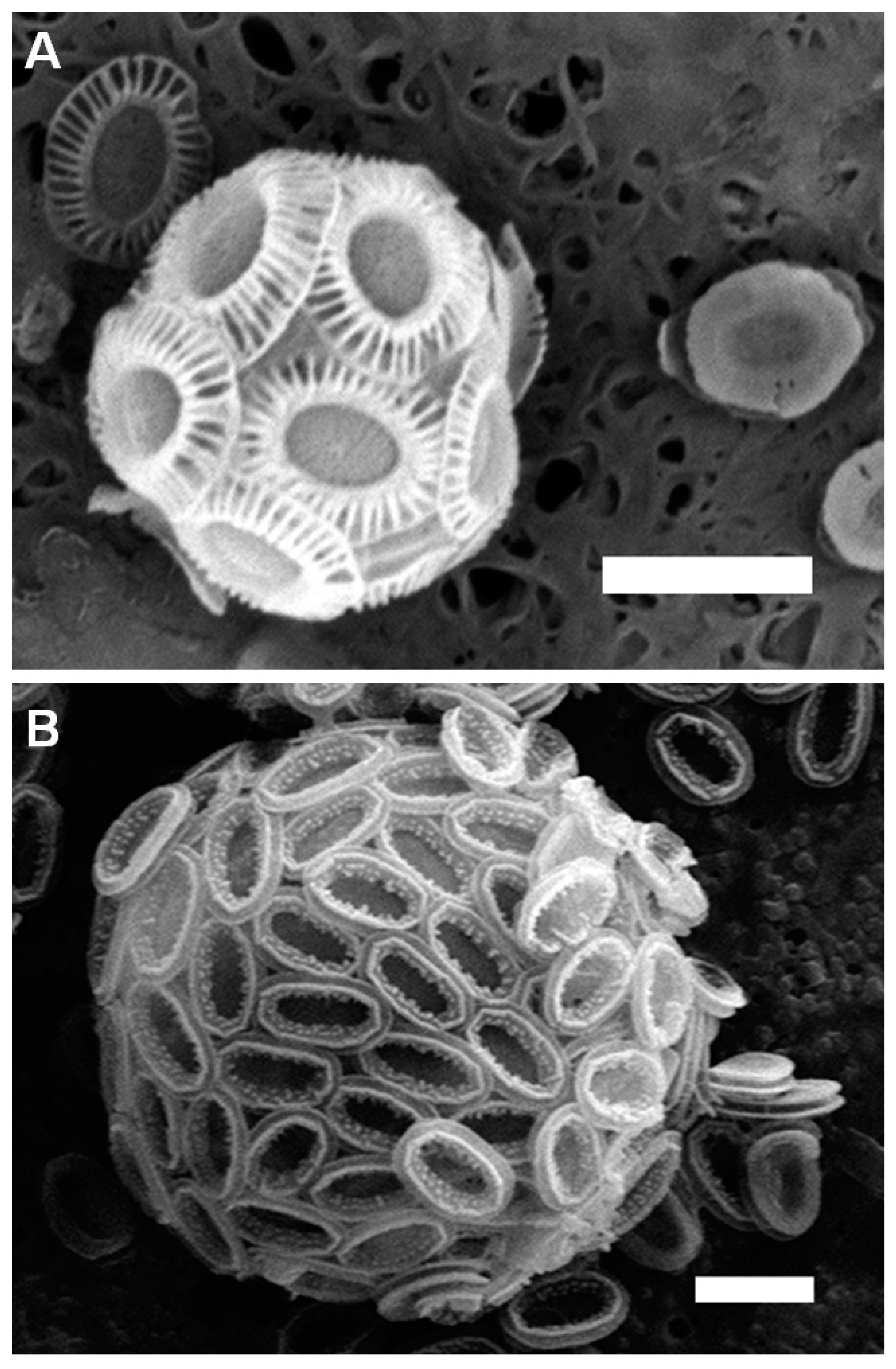
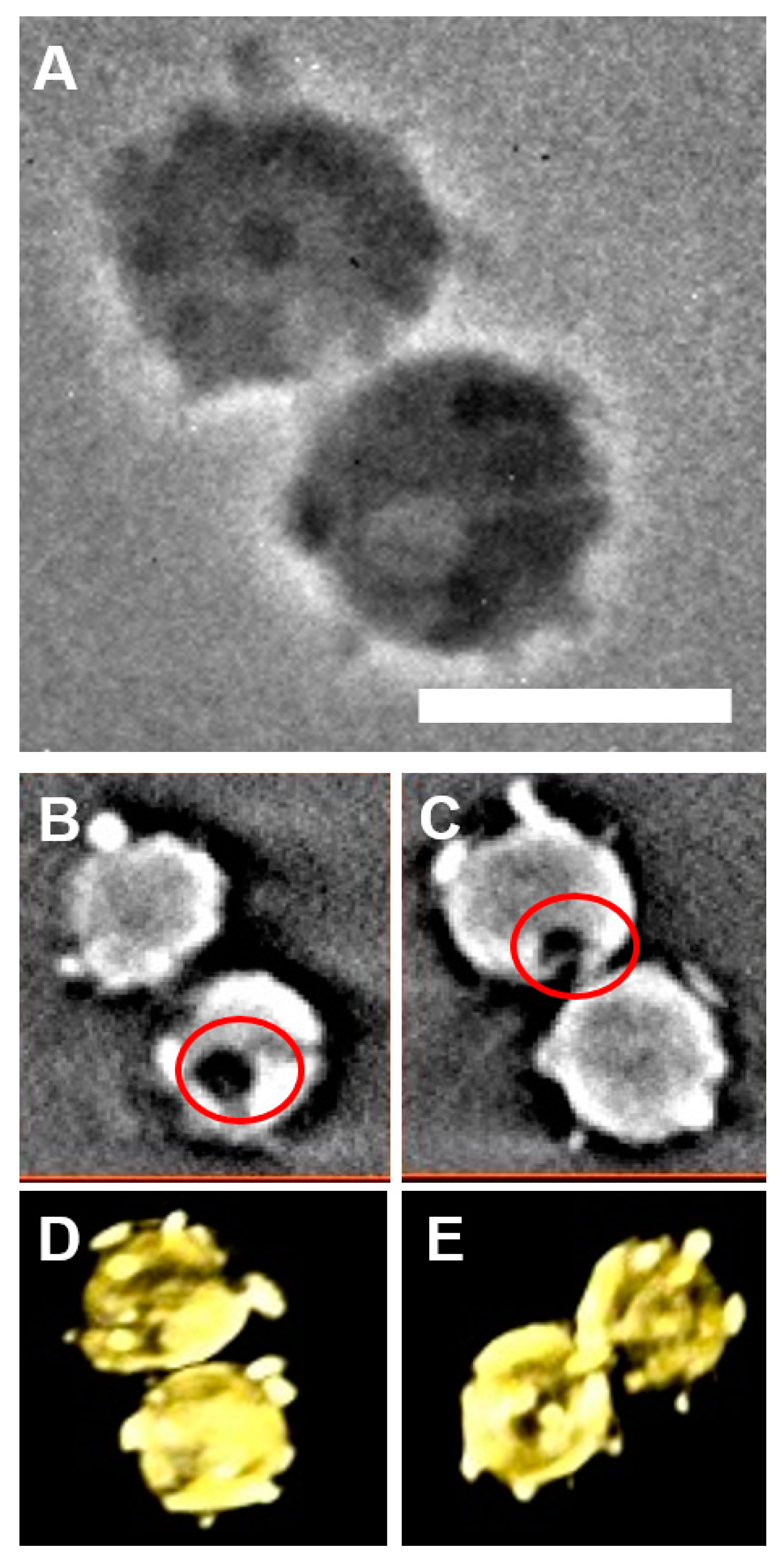
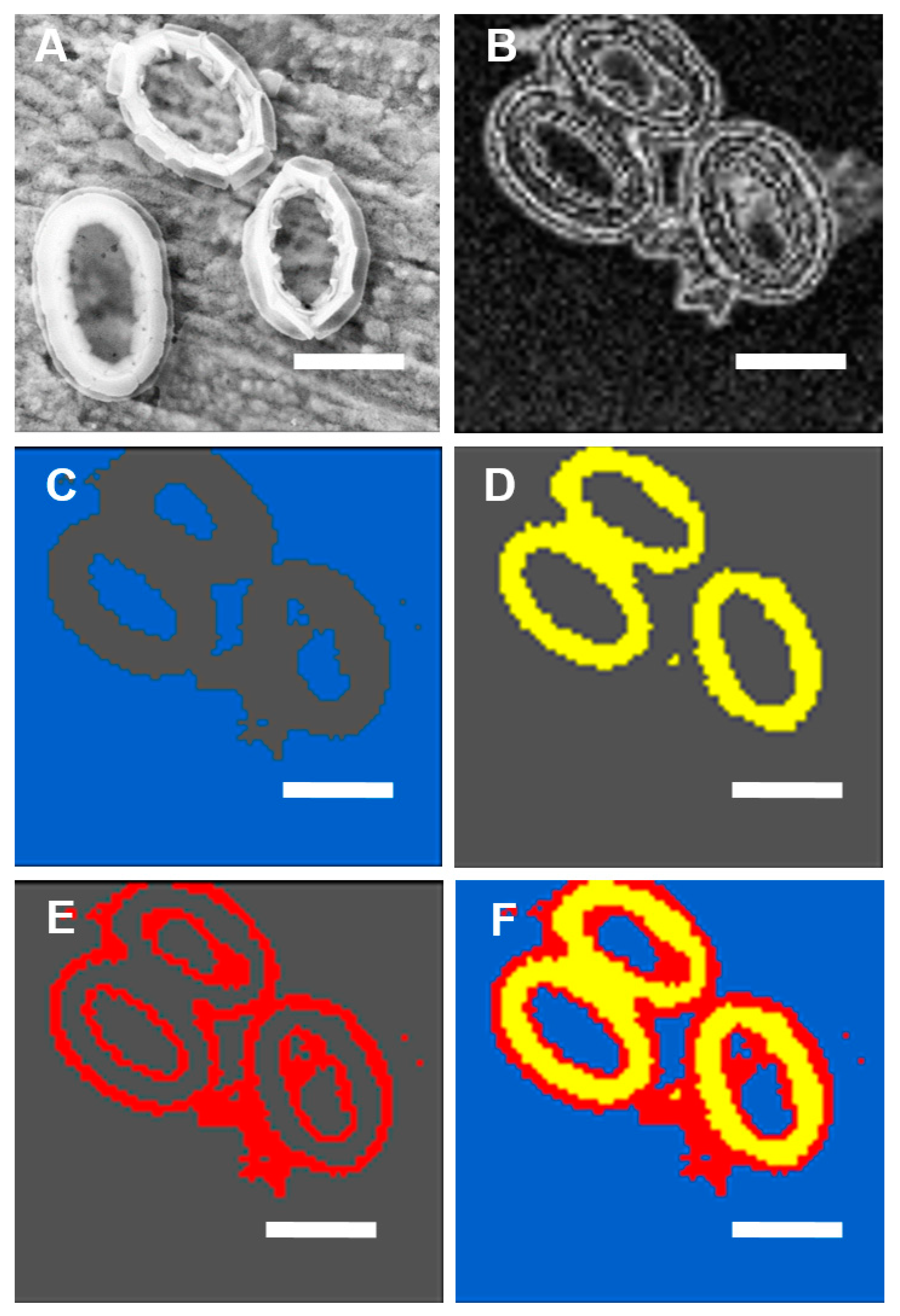
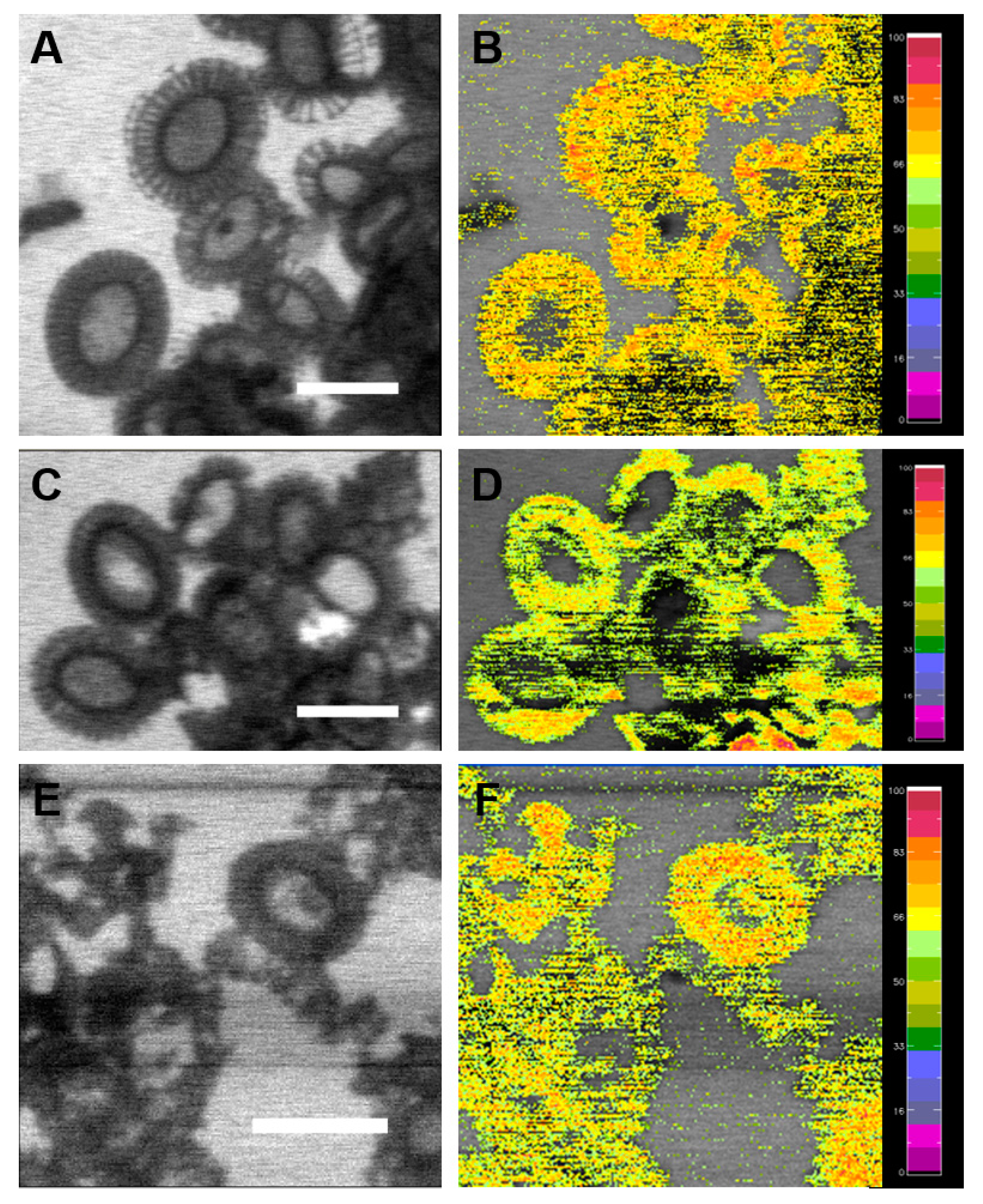
2.1. Spatial Distribution of Coccoliths

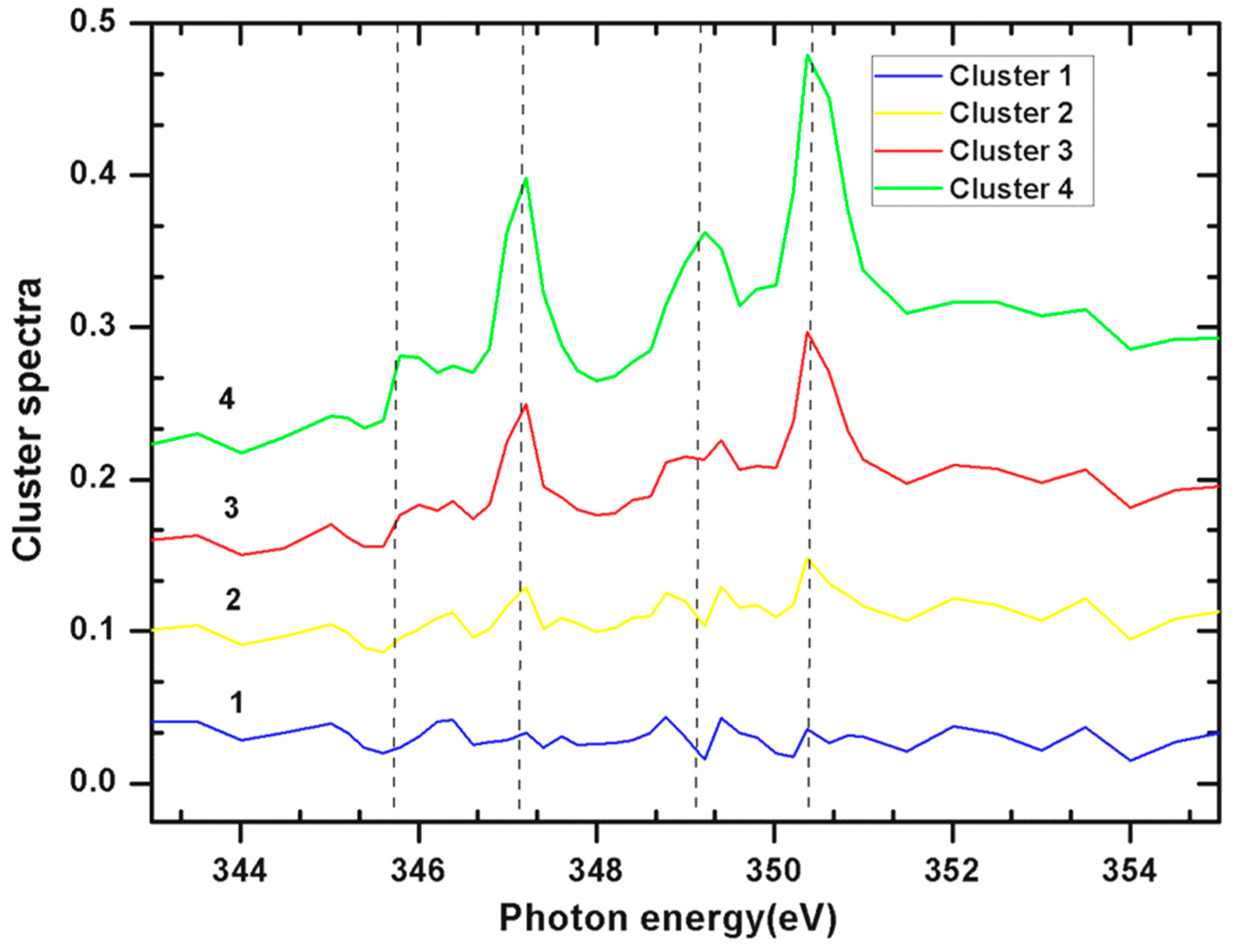
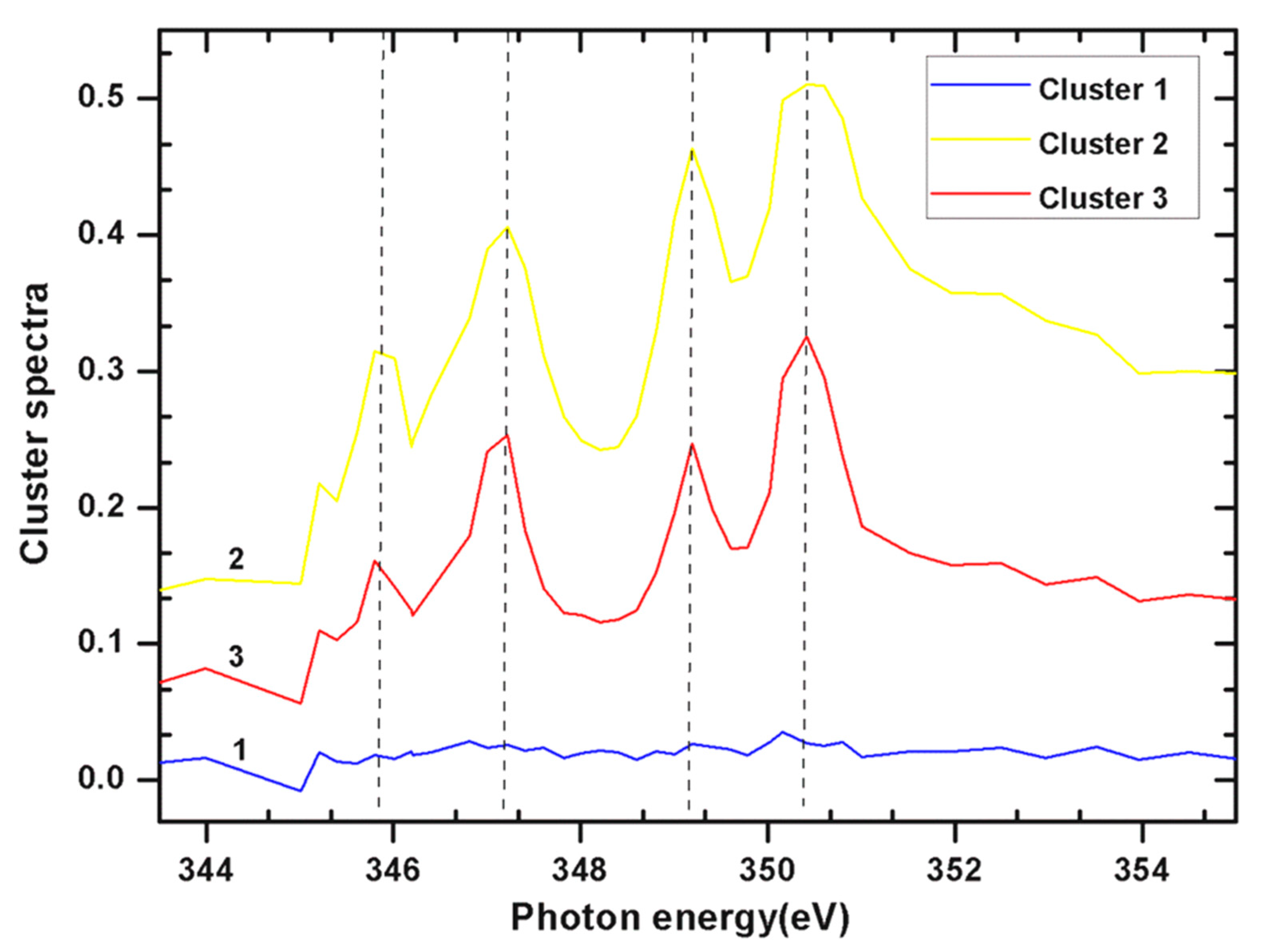
2.2. Ca L Edge of Coccoliths



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster Index | Pixels in Cluster % | Pixels in Cluster % |
|---|---|---|
| Emilianiahuxleyi | Pleurochrysis carterae | |
| Cluster 1 | 33.949 | 70.5536 |
| Cluster 2 | 30.1769 | 15.0029 |
| Cluster 3 | 24.3236 | 14.4435 |
| Cluster 4 | 11.5505 |


2.3. Sr/Ca Substitution

| Original Sr | Ca | Mg | Sr | Sr/Ca | Mg/Ca |
|---|---|---|---|---|---|
| ppm | % | % | % | ||
| 0 | 5.3 | 0.43 | 0.02 | 0.004 | 0.081 |
| 50 | 6.21 | 1.72 | 0.11 | 0.018 | 0.277 |
| 100 | 6.78 | 3.28 | 0.23 | 0.034 | 0.484 |
3. Experimental Section
3.1. Culture Conditions
3.2. Nano-CT
3.3. STXM-NEXAFS
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Taylor, A.R.; Russell, M.A.; Harper, G.M.; Collins, T.F.T.; Brownlee, C. Dynamics of formation and secretion of heterococcoliths by coccolithus pelagicus ssp. Braarudii. Eur. J. Phycol. 2007, 42, 125–136. [Google Scholar] [CrossRef]
- Kayano, K.; Saruwatari, K.; Kogure, T.; Shiraiwa, Y. Effect of coccolith polysaccharides isolated from the coccolithophorid, emiliania huxleyi, on calcite crystal formation in in vitro caco 3 crystallization. Mar. Biotechnol. 2011, 13, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Young, J.R.; Davis, S.A.; Bown, P.R.; Mann, S. Coccolith ultrastructure and biomineralisation. J. Struct. Biol. 1999, 126, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Young, J.R.; Henriksen, K. Biomineralization within vesicles: The calcite of coccoliths. Rev. Mineral. Geochem. 2003, 54, 189–215. [Google Scholar] [CrossRef]
- Young, J.R.; Andruleit, H.; Probert, I. Coccolith function and morphogenesis: Insights from appendage-bearing coccolithophores of the family syracosphaeraceae(haptophyta). J. Phycol. 2009, 45, 213–226. [Google Scholar] [CrossRef]
- Saruwatari, K.; Ozaki, N.; Nagasawa, H.; Kogure, T. Crystallographic alignments in a coccolith (pleurochrysis carterae) revealed by electron back-scattered diffraction (ebsd). Am. Mineral. 2006, 91, 1937–1940. [Google Scholar] [CrossRef]
- Lin, H.; Chen, G.; Zhu, S.; Chen, Y.; Chen, D.; Xu, W.; Yu, X.; Shi, J. The interaction of cus and halothiobacillus ht1 biofilm in microscale using synchrotron radiation-based techniques. Int. J. Mol. Sci. 2013, 14, 11113–11124. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Li, W.; Guan, Y.; Song, X.; Xiong, Y.; Liu, G.; Tian, Y. Quantitative 3d imaging of yeast by hard x-ray tomography. Microsc. Res. Tech. 2012, 75, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, Y.; Zhang, X.; Andrews, J.; Pianetta, P.; Guan, Y.; Liu, G.; Xiong, Y.; Wu, Z.; Tian, Y. 3d nanoscale imaging of the yeast, schizosaccharomyces pombe, by full-field transmission x-ray microscopy at 5.4 kev. Anal. Bioanal. Chem. 2010, 397, 2117–2121. [Google Scholar] [CrossRef] [PubMed]
- Obst, M.; Schmid, G. 3D chemical mapping: Application of scanning transmission (soft) X-ray microscopy (stxm) in combination with angle-scan tomography in bio-, geo-, and environmental sciences. In Electron Microscopy; Kuo, J., Ed.; Humana Press: New York, NY, USA, 2014; Volume 1117, pp. 757–781. [Google Scholar]
- Obst, M.; Wang, J.; Hitchcock, A.P. Soft X-ray spectro-tomography study of cyanobacterial biomineral nucleation. Geobiology 2009, 7, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Obst, M.; Dynes, J.J.; Lawrence, J.R.; Swerhone, G.D.W.; Benzerara, K.; Karunakaran, C.; Kaznatcheev, K.; Tyliszczak, T.; Hitchcock, A.P. Precipitation of amorphous caco3 (aragonite-like) by cyanobacteria: A stxm study of the influence of eps on the nucleation process. Geochim. Cosmochim. Acta 2009, 73, 4180–4198. [Google Scholar] [CrossRef]
- Lerotic, M.; Jacobsen, C.; Gillow, J.; Francis, A.; Wirick, S.; Vogt, S.; Maser, J. Cluster analysis in soft X-ray spectromicroscopy: Finding the patterns in complex specimens. J. Electron. Spectrosc. Relat. Phenom. 2005, 144, 1137–1143. [Google Scholar] [CrossRef]
- Naftel, S.; Sham, T.; Yiu, Y.; Yates, B. Calcium l-edge xanes study of some calcium compounds. J. Synchrot. Radiat. 2001, 8, 255–257. [Google Scholar] [CrossRef]
- Fleet, M.E.; Liu, X. Calcium l2, 3-edge xanes of carbonates, carbonate apatite, and oldhamite (cas). Am. Mineral. 2009, 94, 1235–1241. [Google Scholar] [CrossRef]
- Xu, K.; Gao, K. Reduced calcification decreases photoprotective capability in the coccolithophorid emiliania huxleyi. Plant Cell Physiol. 2012, 53, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Li, W.J.; Wang, N.; Chen, J.; Liu, G.; Pan, Z.Y.; Guan, Y.; Yang, Y.H.; Wu, W.Q.; Tian, J.P.; Wei, S.Q.; et al. Quantitative study of interior nanostructure in hollow zinc oxide particles on the basis of nondestructive x-ray nanotomography. Appl. Phys. Lett. 2009, 95, 053108. [Google Scholar] [CrossRef]
- Xue, C.; Wang, Y.; Guo, Z.; Wu, Y.; Zhen, X.; Chen, M.; Chen, J.; Xue, S.; Peng, Z.; Lu, Q.; et al. High-performance soft X-ray spectromicroscopy beamline at ssrf. Rev. Sci. Instrum. 2010, 81, 103502. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xu, Z.; Tai, R.; Zhen, X.; Wang, Y.; Guo, Z.; Yan, R.; Chang, R.; Wang, B.; Li, M. Ratio-contrast imaging of dual-energy absorption for element mapping with a scanning transmission X-ray microscope. J. Synchrot. Radiat. 2010, 17, 804–809. [Google Scholar] [CrossRef]
- Lerotic, M.; Jacobsen, C.; Schäfer, T.; Vogt, S. Cluster analysis of soft X-ray spectromicroscopy data. Ultramicroscopy 2004, 100, 35–57. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, S.; Yao, Y.; Zou, X.; Fan, S.; Zhou, Q.; Dai, Q.; Dong, F.; Liu, M.; Nie, X.; Tan, D.; et al. Nano-Scale Spatial Assessment of Calcium Distribution in Coccolithophores Using Synchrotron-Based Nano-CT and STXM-NEXAFS. Int. J. Mol. Sci. 2014, 15, 23604-23615. https://doi.org/10.3390/ijms151223604
Sun S, Yao Y, Zou X, Fan S, Zhou Q, Dai Q, Dong F, Liu M, Nie X, Tan D, et al. Nano-Scale Spatial Assessment of Calcium Distribution in Coccolithophores Using Synchrotron-Based Nano-CT and STXM-NEXAFS. International Journal of Molecular Sciences. 2014; 15(12):23604-23615. https://doi.org/10.3390/ijms151223604
Chicago/Turabian StyleSun, Shiyong, Yanchen Yao, Xiang Zou, Shenglan Fan, Qing Zhou, Qunwei Dai, Faqin Dong, Mingxue Liu, Xiaoqin Nie, Daoyong Tan, and et al. 2014. "Nano-Scale Spatial Assessment of Calcium Distribution in Coccolithophores Using Synchrotron-Based Nano-CT and STXM-NEXAFS" International Journal of Molecular Sciences 15, no. 12: 23604-23615. https://doi.org/10.3390/ijms151223604