Testosterone Reduces Knee Passive Range of Motion and Expression of Relaxin Receptor Isoforms via 5α-Dihydrotestosterone and Androgen Receptor Binding

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
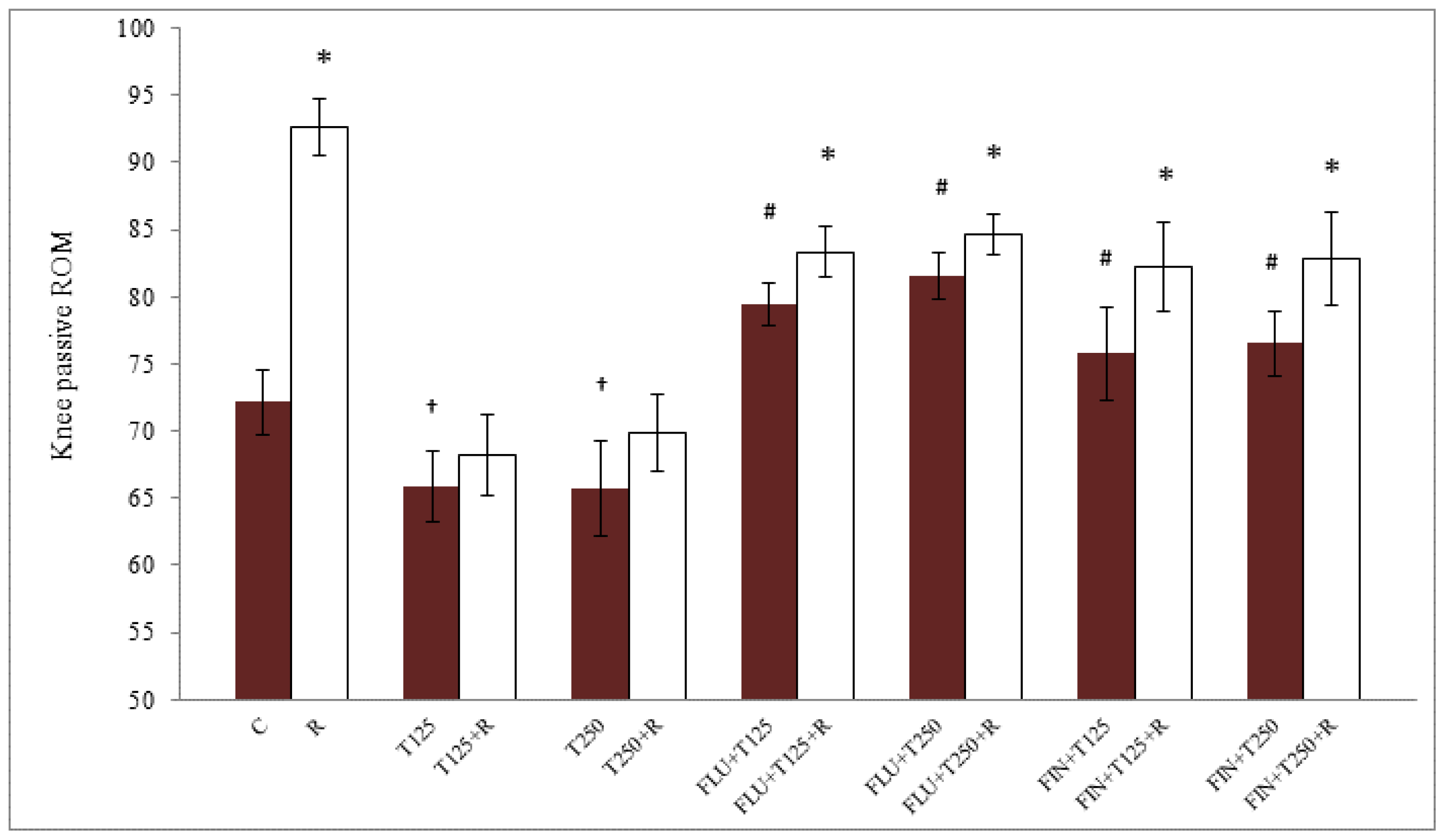
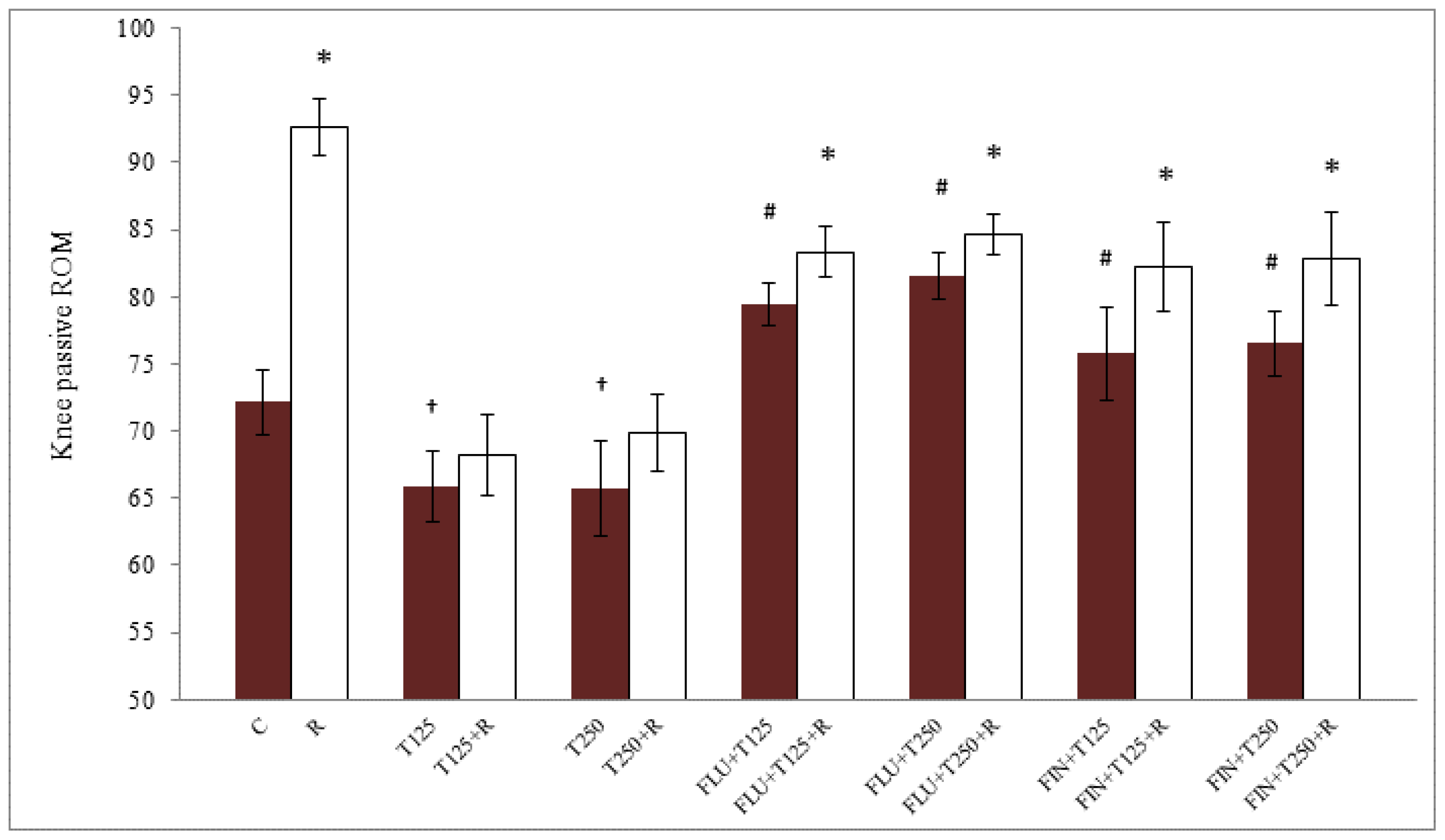
2.1. Passive Knee ROM in Testosterone-Treated Ovariectomised Rats
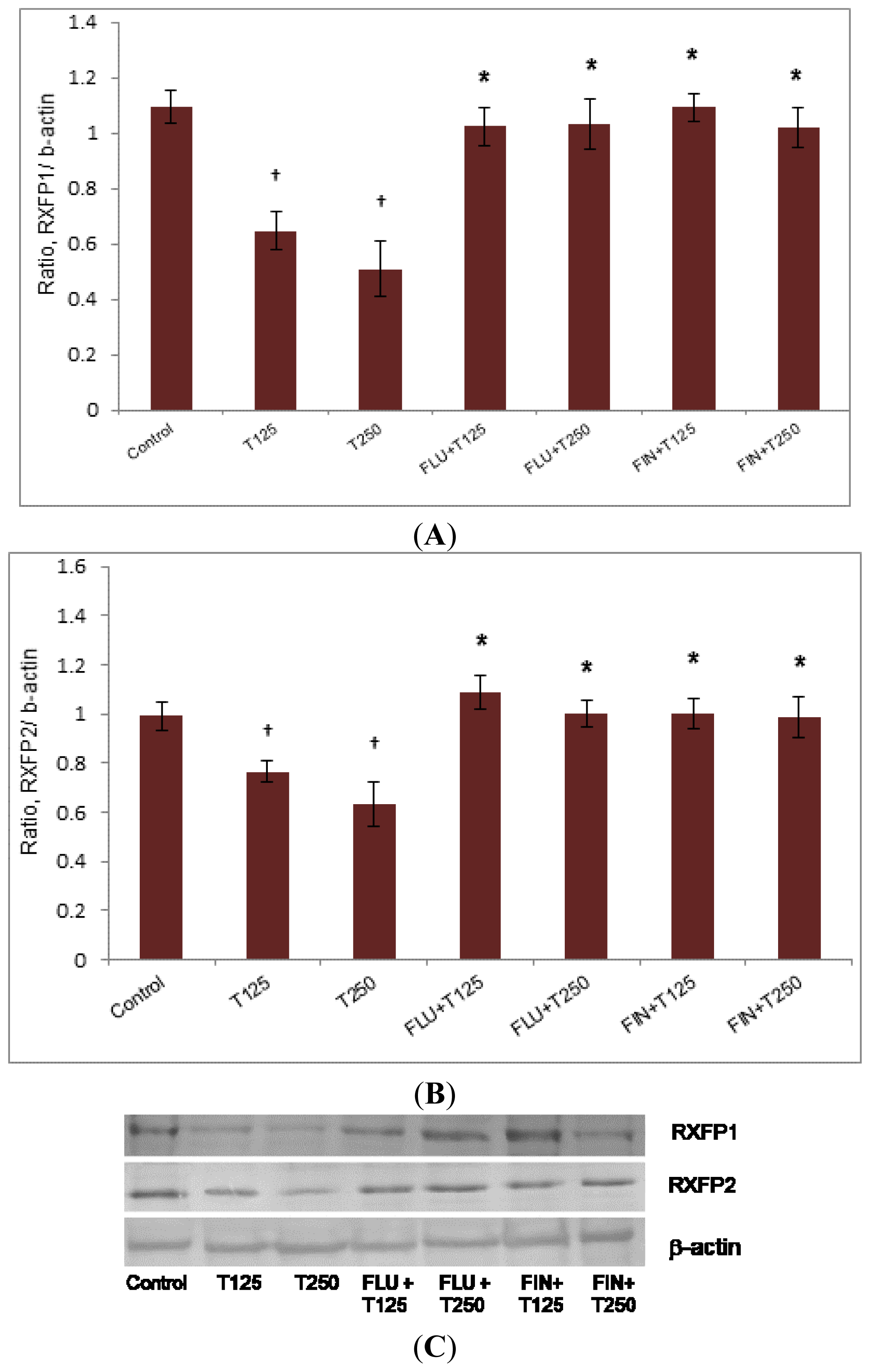
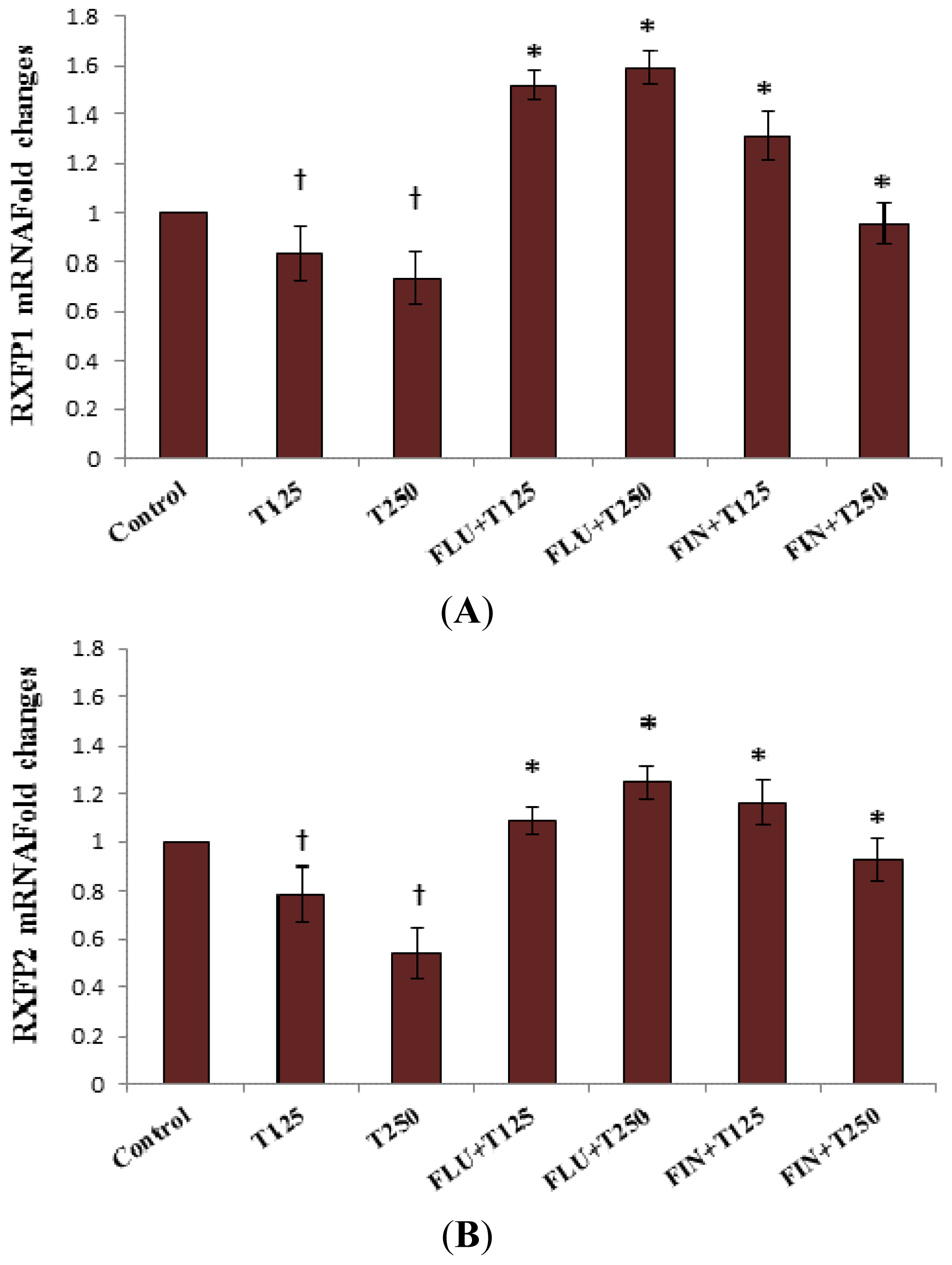
2.2. Rxfp1 and Rxfp2 mRNA Expression in Patellar Tendon
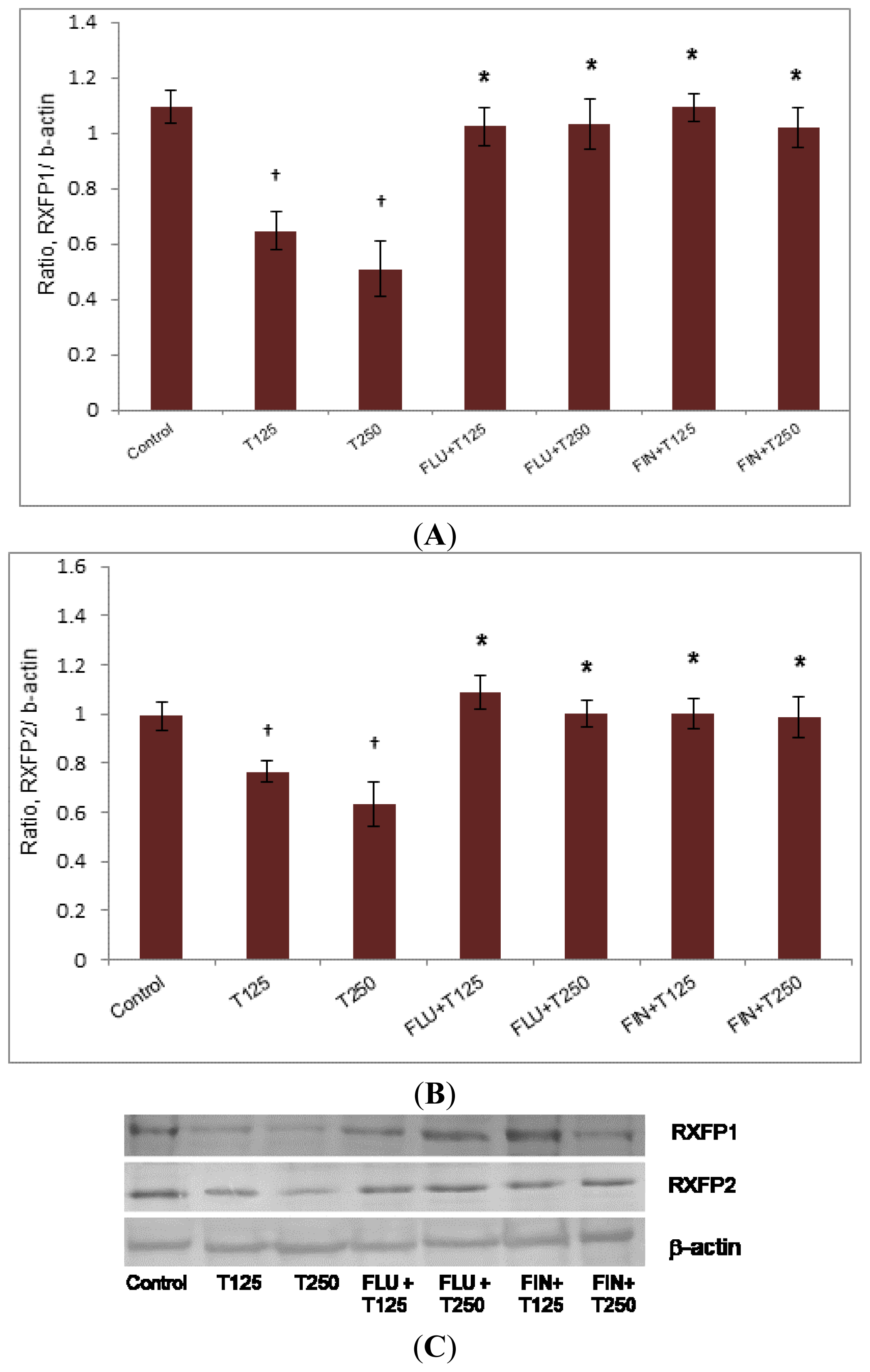
2.3. Rxfp1 and Rxfp2 Protein Expression in Patellar Tendon
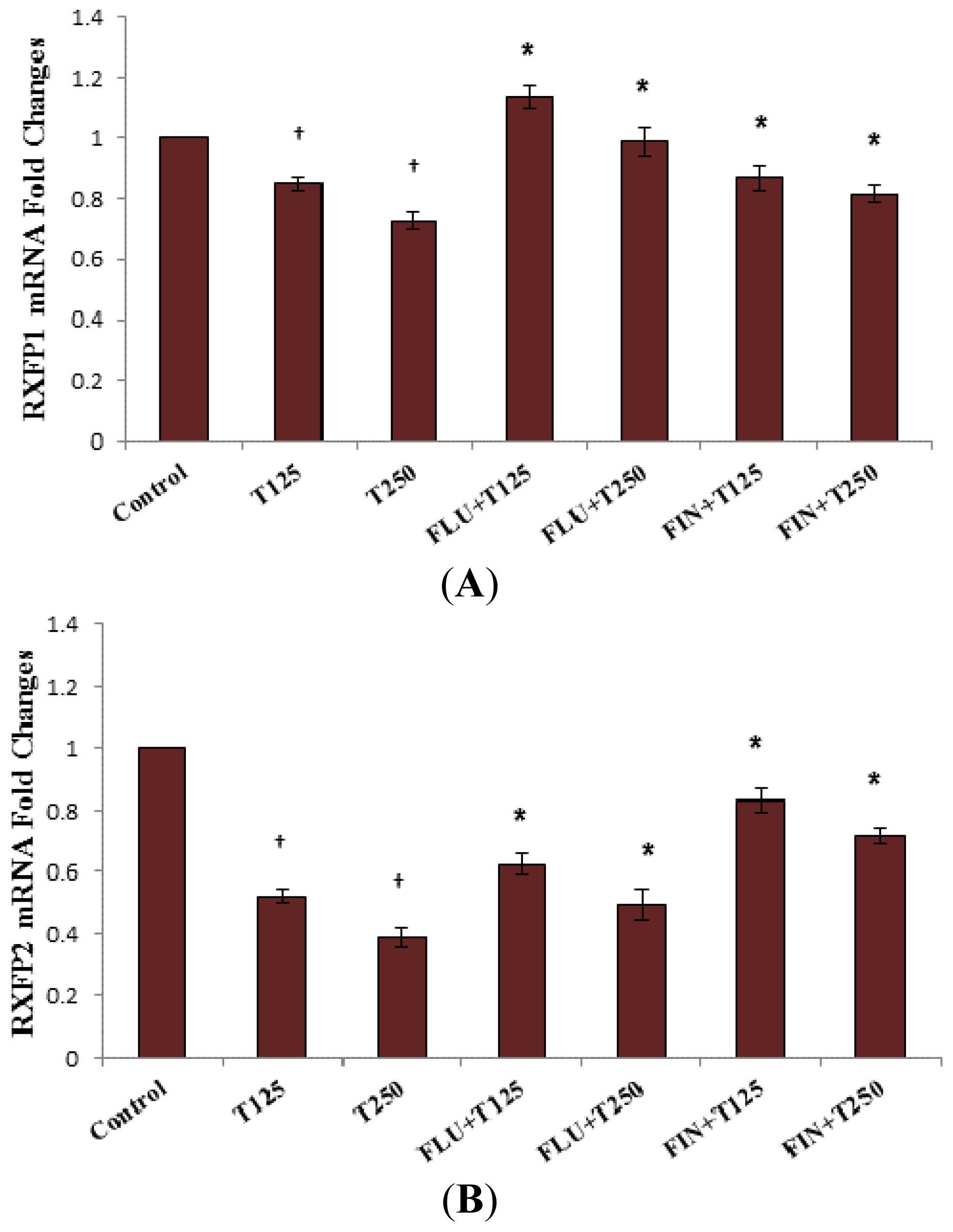
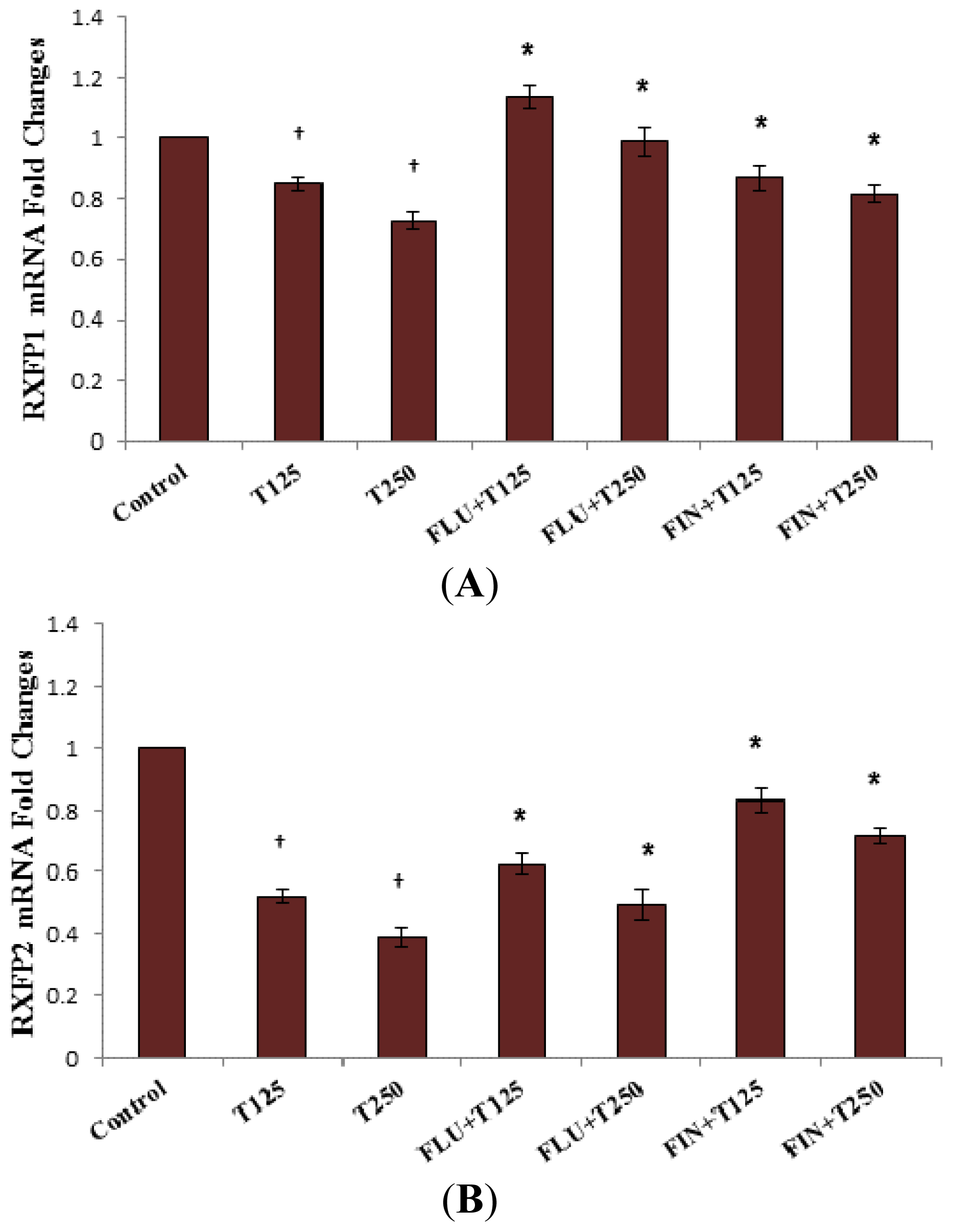
2.4. Rxfp1 and Rxfp2 mRNA Expression in Lateral Collateral Ligament
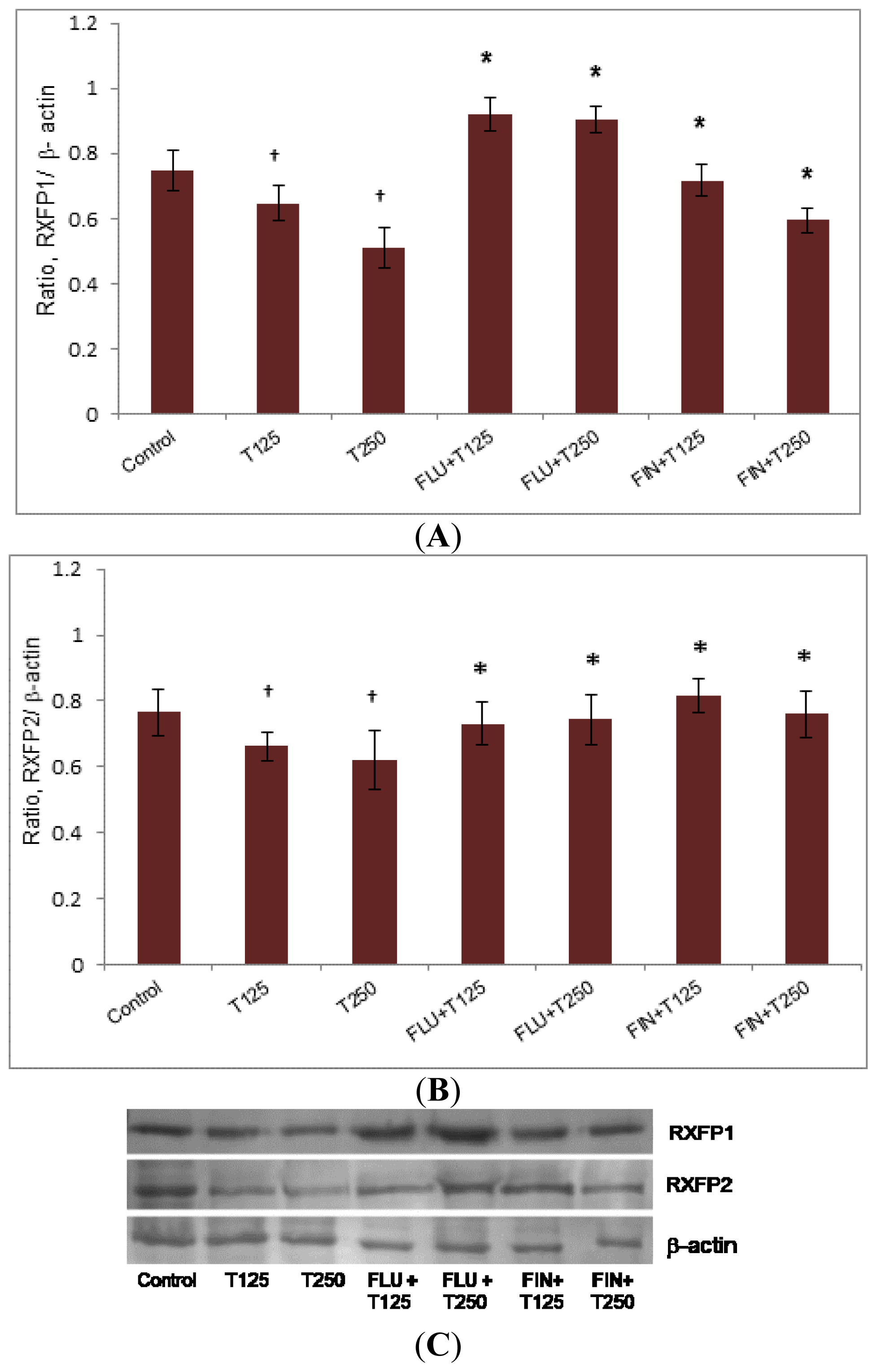
2.5. Rxfp1 and Rxfp2 Protein Expression in Lateral Collateral Ligament
3. Discussion
4. Experimental Sections
4.1. Animal Preparation and Hormone Treatment
4.2. Measurement of Knee ROM Following Sex-Steroid Treatments
4.3. Rxfp1 and Rxfp2 mRNA Expression Analyses by Real Time PCR (qPCR)
4.4. Protein Expression Analysis by Western Blotting
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Schmitzemail, R.J.; Sauret, J.J.; Shultz, S.J. Anterior tibiofemoral intersegmental forces during landing are predicted by passive restraint measures in women. Knee 2013, 20, 493–499. [Google Scholar]
- Marieb, E.; Hoehn, K. Anatomy & Physiology, 5th ed.; Benjamin Cummings: San Francisco, CA, USA, 2013. [Google Scholar]
- Janousek, A.T.; Jones, D.G.; Clatworthy, M.; Higgins, L.D.; Fu, F.H. Posterior cruciate ligament injuries of the knee joint. Sports Med 1999, 28, 429–441. [Google Scholar]
- Wojtys, E.M.; Huston, L.J.; Schock, H.J.; Boylan, J.P.; Ashton-Miller, J.A. Gender differences in muscular protection of the knee in torsion in size-matched athletes. J. Bone Jt. Surg 2003, 85A, 782–789. [Google Scholar]
- Hsu, W.H.; Fisk, J.A.; Yamamoto, Y.; Debski, R.E.; Woo, S.L. Differences in torsional joint stiffness of the knee between genders: A human cadaveric study. Am. J. Sports Med 2006, 34, 765–570. [Google Scholar]
- Beynnon, B.D.; Bernstein, I.M.; Belisle, A.; Brattbakk, B.; Devanny, P.; Risinger, R.; Durant, D. The effect of estradiol and progesterone on knee and ankle joint laxity. Am. J. Sports Med 2005, 33, 1298–1304. [Google Scholar]
- Dragoo, J.L.; Padrez, K.; Workman, R.; Lindsey, D.P. The effect of relaxin on the female anterior cruciate ligament: Analysis of mechanical properties in an animal model. Knee 2009, 16, 69–72. [Google Scholar]
- Chen, C.Z.; Southall, N.; Xiao, J.; Marugan, J.J.; Ferrer, M.; Hu, X.; Jones, R.E.; Feng, S.; Agoulnik, I.U.; Zheng, W.; et al. Identification of small-molecule agonists of human relaxin family receptor 1 (RXFP1) by using a homogenous cell-based cAMP assay. J. Biomol. Screen 2012, 153, 4655–4665. [Google Scholar]
- Sherwood, O.D. Relaxin’s physiological roles and other diverse actions. Endocr. Rev 2004, 25, 205–234. [Google Scholar]
- Lee, W.J.; Choi, I.-K.; Lee, J.H.; Lee, J.-S.; Kim, Y.O.; Rah, D.K.; Yun, C.-O. Relaxin-expressing adenovirus decreases collagen synthesis and up-regulates matrix metalloproteinase expression in keloid fibroblasts: in vitro experiments. Plast. Reconstr. Surg 2012, 130, 407e–417e. [Google Scholar]
- Scott, D.J.; Rosengren, K.J.; Bathgate, R.A.D. The different ligand-binding modes of relaxin family peptide receptors RXFP1 and RXFP2. Mol. Endocrinol 2012, 26, 1896–1906. [Google Scholar]
- Scott, D.J.; Layfield, S.; Riesewijk, A.; Morita, H.; Tregear, G.W.; Bathgate, R.A.D. Characterization of the mouse and rat relaxin receptors. Ann. N. Y. Acad. Sci 2005, 1041, 8–12. [Google Scholar]
- Bathgate, R.A.; Ivell, R.; Sanborn, B.M.; Sherwood, O.D.; Summers, R.J. Receptors for relaxin family peptides. Ann. N. Y. Acad. Sci 2005, 1041, 61–76. [Google Scholar]
- Kumagai, J.; Hsu, S.Y.; Matsumi, H.; Roh, J.S.; Fu, P.; Wade, J.D.; Bathgate, R.A.D.; Hsueh, A.J.W. INSL3/Leydig insulin-like peptide activates the LGR8 receptor important in testis descent. J. Biol. Chem 2002, 277, 31283–31286. [Google Scholar]
- Forst, J.; Forst, C.; Forst, R.; Heller, K.-D. Pathogenetic relevance of the pregnancy hormone relaxin to inborn hip instability. Arch. Orthop. Trauma Surg 1997, 116, 209–212. [Google Scholar]
- Saugstad, L.F. Persistent pelvic pain and pelvic joint instability. Eur. J. Obstet. Gynecol. Reprod. Biol 1991, 41, 197–201. [Google Scholar]
- Steinetz, B.G.; Williams, A.J.; Lust, G.S.; Schwabe, C.; Büllesbach, E.E.; Goldsmith, L.T. Transmission of relaxin and estrogens to suckling canine pups via milk and possible association with hip joint laxity. Am. J. Vet. Res 2008, 69, 59–67. [Google Scholar]
- Dragoo, J.L.; Castillo, T.N.; Braun, H.J.; Ridley, B.A.; Kennedy, A.C.; Golish, S.R. Prospective correlation between serum relaxin concentration and anterior cruciate ligament tears among elite collegiate female athletes. Am. J. Sports Med 2011, 39, 2175–2180. [Google Scholar]
- He, H.; Yang, F.; Liu, X.; Zeng, X.; Hu, Q.; Zhu, Q.; Tu, B. Sex hormone ratio changes in men and postmenopausal women with coronary artery disease. Menopause 2007, 14, 385–390. [Google Scholar]
- Barrett, K.E.; Susan, M.S.; Boitano, S.; Brooks, H. Ganong Review of Medical Physiology; McGraw-Hill Medical: Maidenhead, UK, 2009; p. 726. [Google Scholar]
- Ishikawa, T.; Harada, T.; Kubota, T.; Aso, T. Testosterone inhibits matrix metalloproteinase-1 production in human endometrial stromal cells in vitro. Reproduction 2007, 133, 1233–1239. [Google Scholar]
- Kouloumenta, V.; Hatziefthimiou, A.; Paraskeva, E.; Gourgoulianis, K. Molyvdas PANon-genomic effect of testosterone on airway smooth muscle. Br. J. Pharmacol 2006, 149, 1083–1091. [Google Scholar]
- Shultz, S.J.; Sander, T.C.; Kirk, S.E.; Perrin, D.H. Sex differences in knee joint laxity change across the female menstrual cycle. J. Sports Med. Phys. Fit 2005, 45, 594–603. [Google Scholar]
- Rozzi, S.L.; Lephart, S.M.; Gear, W.S.; Fu, F.H. Knee joint laxity and neuromuscular characteristics of male and female soccer and basketball players. Am. J. Sports Med 1999, 27, 312–319. [Google Scholar]
- Lovering, R.M.; Romani, W.A. Effect of testosterone on the female anterior cruciate ligament. Am. J. Physiol.-Regul. Integr. Compar. Physiol 2005, 289, R15–R22. [Google Scholar]
- Shultz, S.J.; Kirk, S.E.; Johnson, M.L.; Sander, T.C.; Perrin, D.H. Relationship between sex hormones and anterior knee laxity across the menstrual cycle. Med. Sci. Sports Exerc 2004, 36, 1165–1174. [Google Scholar]
- Speroff, L.; Glass, R.H.; Kase, N.G. Hormone Biosynthesis, Metabolism and Mechanism of Action, Clinical Gynecologic Endocrinology and Infertility; Williams & Wilkins: Baltimore, MD, USA, 1989. [Google Scholar]
- Rao, A.J.; Kotagi, S.G. Serum testosterone levels during the menstrual cycle and early pregnancy in the bonnet monkey (Macaca radiata). Endocrinol. Jpn 1982, 29, 271–275. [Google Scholar]
- O’Leary, C.B.; Lehman, C.; Koltun, K.; Smith-Ryan, A.; Hackney, A.C. Response of testosterone to prolonged aerobic exercise during different phases of the menstrual cycle. Eur. J. Appl. Physiol 2013, 113, 2419–2424. [Google Scholar]
- Srinivasan, N.; Micheal Aruldhas, M.; Govindarajulu, P. Sex steroid induced changes in collagen of the prostate and seminal vesicle of rats. J. Androl 1986, 7, 55–58. [Google Scholar]
- Asano, K.; Maruyama, S.; Usui, T.; Fujimoto, N. Regulation of estrogen receptor alpha and alpha expression by testosterone in the rat prostate gland. Endocr. J 2003, 50, 281–287. [Google Scholar]
- Dragoo, J.L.; Lee, R.S.; Benhaim, P.; Finerman, G.A.; Hame, S.L. Relaxin receptors in the human female anterior cruciate ligament. Am. J. Sports Med 2003, 31, 577–584. [Google Scholar]
- Bagnell, C.A.; Zhang, Q.; Downey, B.; Ainsworth, L. Sources and biological actions of relaxin in pigs. J. Reprod. Fertil. Suppl 1993, 48, 127–138. [Google Scholar]
- Fytili, P.; Giannatou, E.; Papanikolaou, V.; Stripeli, F.; Karachalios, T.; Malizos, K.; Tsezou, A. Association of repeat polymorphisms in the estrogen receptors α β and androgen receptor genes with knee osteoarthritis. Clin. Genet 2005, 68, 268–277. [Google Scholar]
- Frye, S.V.; Bramson, H.N.; Hermann, D.J.; Lee, F.W.; Sinhababu, A.K.; Tian, G. Discovery and development of GG745 a potent inhibitor of both isozymes of 5 alpha-reductase. Pharm. Biotechnol 1998, 11, 393–422. [Google Scholar]
- Cárdenas, H.; Pope, W.F. Estrogen receptors in the uterus and ovarian follicles of gilts treated with dihydrotestosterone. Domest. Anim. Endocrinol 2005, 29, 523–533. [Google Scholar]
- Hamdi, M.M.; Mutungi, G. Dihydrotestosterone activates the MAPK pathway and modulates maximum isometric force through the EGF receptor in isolated intact mouse skeletal muscle fibres. J. Physiol 2010, 588, 511–525. [Google Scholar]
- Bhasin, S.; Storer, T.W.; Berman, N.; Callegari, C.; Clevenger, B.; Phillips, J.; Bunnell, T.J.; Tricker, R.; Shirazi, A.; Casaburi, R. The effects of supraphysiologic doses of testosterone on muscle size and strength in normal men. N. Engl. J. Med 1996, 335, 1–7. [Google Scholar]
- Wolf, J.M.; Williams, A.E.; Delaronde, S.; Leger, R.; Clifton, K.B.; King, K.B. Relationship of serum relaxin to generalized and trapezial-metacarpal joint laxity. J. Hand Surg 2013, 38, 721–728. [Google Scholar]
- Porkka-Heiskanen, T.; Laakso, M.L.; Stenberg, D. The effect of testosterone on serum gonadotropins of castrated rats kept under different lighting conditions. Biol. Reprod 1992, 46, 1127–1129. [Google Scholar]
- Silversides, D.W.; Price, C.A.; Cooke, G.M. Effects of short-term exposure to hydroxyflutamide in utero on the development of the reproductive tract in male mice. Can. J. Physiol. Pharmacol 1995, 73, 1582–1588. [Google Scholar]
- Zhang, Z.; Yang, R.; Cai, W.; Bai, Y.; Sokabe, M.; Chen, L. Treatment with progesterone after focal cerebral ischemia suppresses proliferation of progenitor cells but enhances survival of newborn neurons in adult male mice. Neuropharmacology 2010, 58, 930–939. [Google Scholar]
- Flecknell, P. Laboratory Animal Anaesthesia, 3rd ed.; Academic Press: Waltham, MA, USA, 2009. [Google Scholar]


© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dehghan, F.; Muniandy, S.; Yusof, A.; Salleh, N. Testosterone Reduces Knee Passive Range of Motion and Expression of Relaxin Receptor Isoforms via 5α-Dihydrotestosterone and Androgen Receptor Binding. Int. J. Mol. Sci. 2014, 15, 4619-4634. https://doi.org/10.3390/ijms15034619
Dehghan F, Muniandy S, Yusof A, Salleh N. Testosterone Reduces Knee Passive Range of Motion and Expression of Relaxin Receptor Isoforms via 5α-Dihydrotestosterone and Androgen Receptor Binding. International Journal of Molecular Sciences. 2014; 15(3):4619-4634. https://doi.org/10.3390/ijms15034619
Chicago/Turabian StyleDehghan, Firouzeh, Sekaran Muniandy, Ashril Yusof, and Naguib Salleh. 2014. "Testosterone Reduces Knee Passive Range of Motion and Expression of Relaxin Receptor Isoforms via 5α-Dihydrotestosterone and Androgen Receptor Binding" International Journal of Molecular Sciences 15, no. 3: 4619-4634. https://doi.org/10.3390/ijms15034619
APA StyleDehghan, F., Muniandy, S., Yusof, A., & Salleh, N. (2014). Testosterone Reduces Knee Passive Range of Motion and Expression of Relaxin Receptor Isoforms via 5α-Dihydrotestosterone and Androgen Receptor Binding. International Journal of Molecular Sciences, 15(3), 4619-4634. https://doi.org/10.3390/ijms15034619