GPC1 Regulated by miR-96-5p, Rather than miR-182-5p, in Inhibition of Pancreatic Carcinoma Cell Proliferation

Abstract
:1. Introduction
2. Results and Discussion
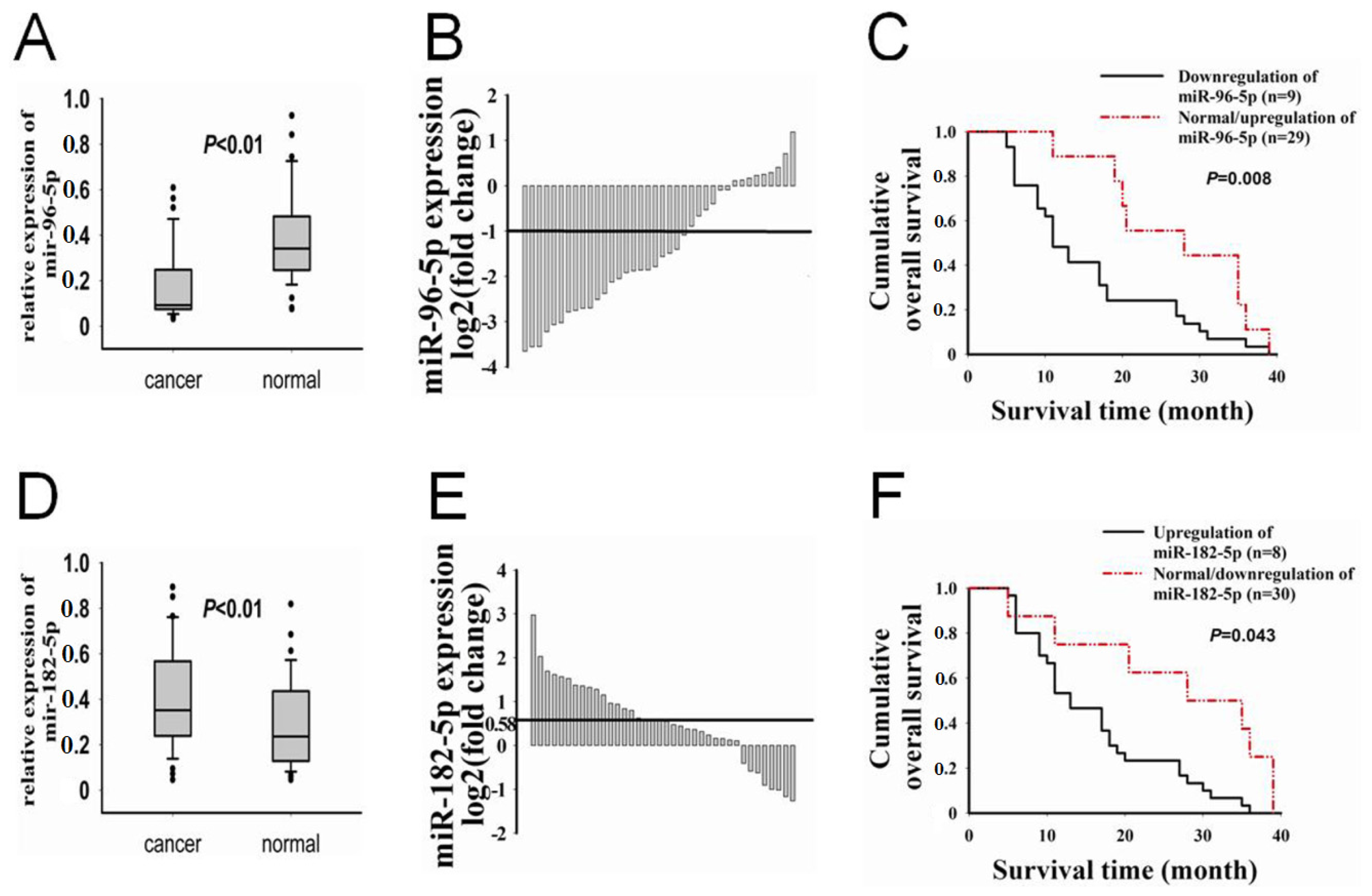
2.1. The Correlations between Expression of miR-96-5p, -182-5p and GPC1 Expression, Clinicopathological Characteristics of PC
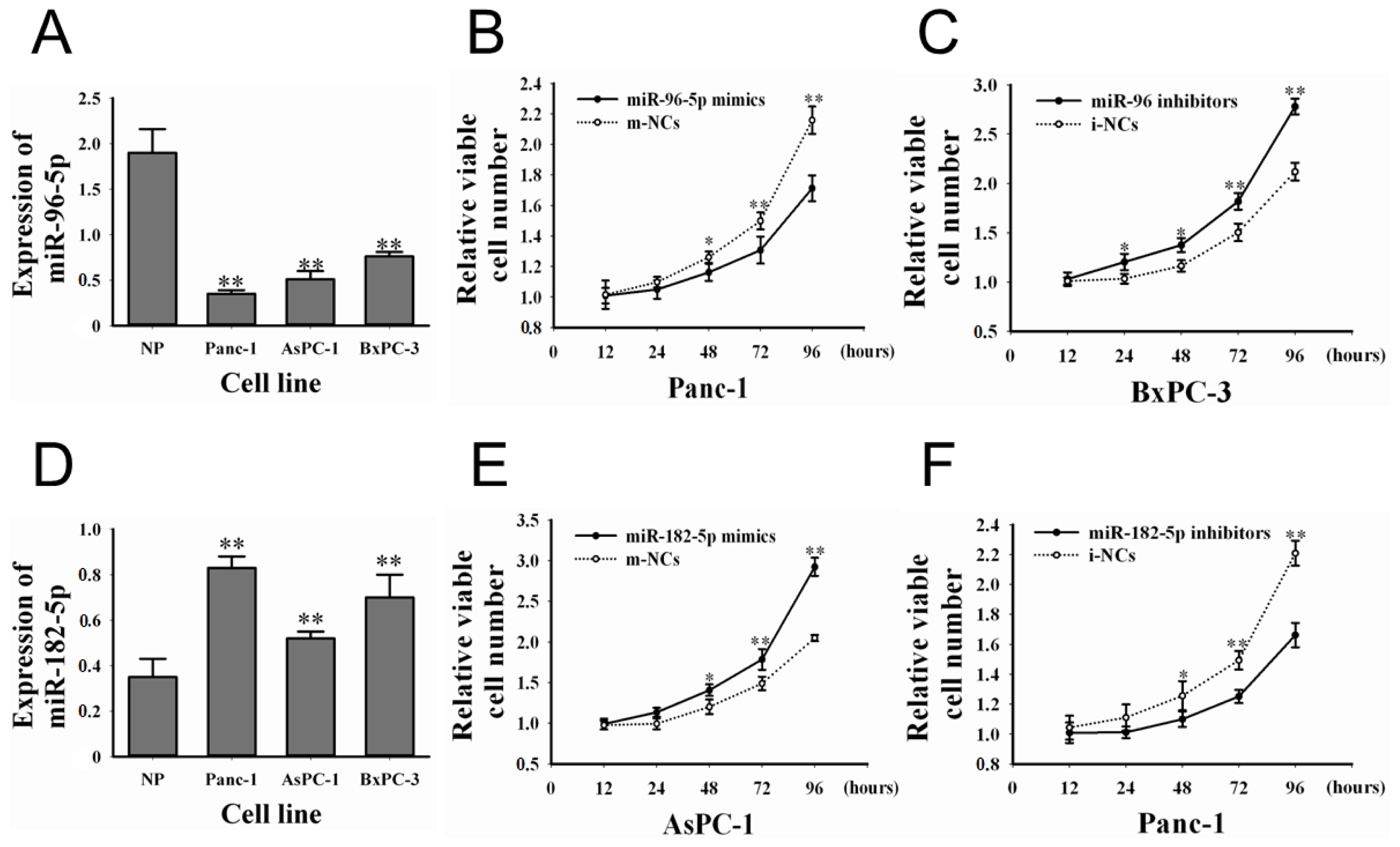
2.2. The Expression of miR-96-5p and -182-5p in Cell Lines and Their Effects on PC Cell Proliferation
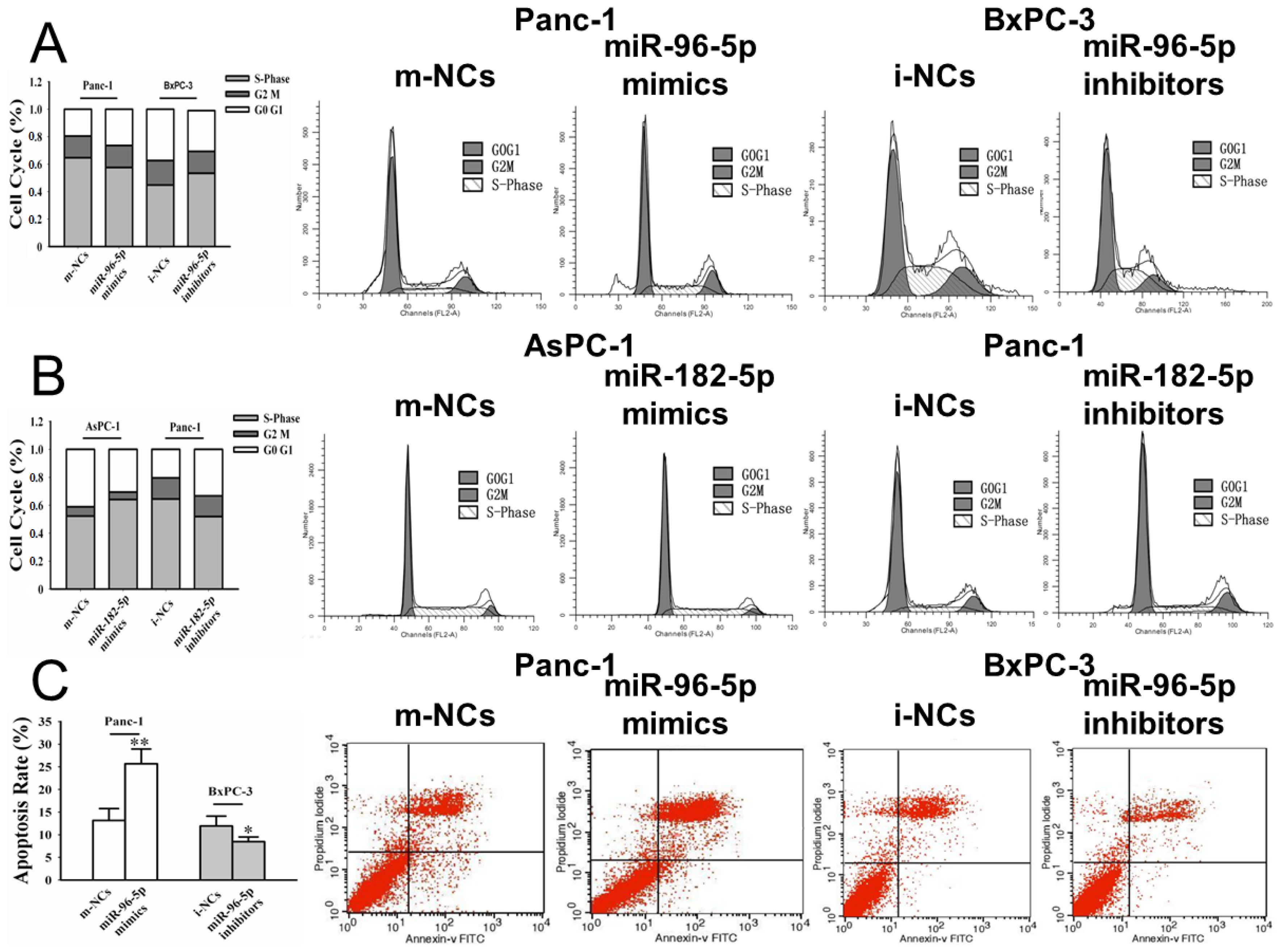
2.3. Effects of miR-96-5p and -182-5p on Cell Cycle and Apoptosis in PC Cells
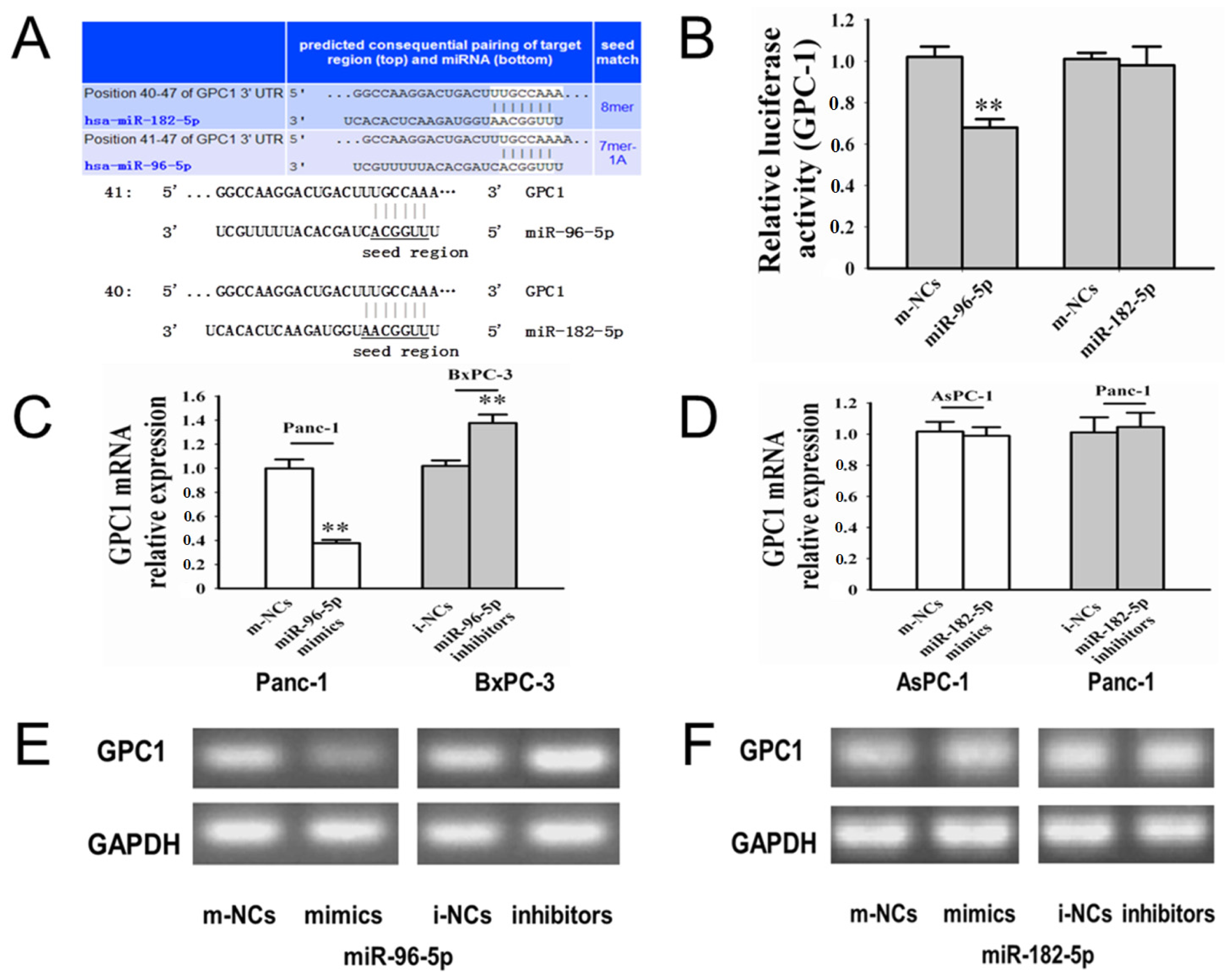
2.4. GPC1 Might Be a Direct Downstream Target for miR-96-5p
2.5. Discussion
2.5.1. Whether miR-96-5p and/or -182-5p Could Regulate GPC1 in PC
2.5.2. Whether miR-182-5p Could Play a Role in PC
2.5.3. The Validation of the miR-96-5p Effects on PC
3. Experimental Section
3.1. Patients and Tissue Samples
3.2. Immunohistochemistry Staining and Staining Evaluation
3.3. Cell Lines and Transfection
3.4. RNA Extraction and qRT-PCR
3.5. Cell Proliferation Assay
3.6. Cell Cycle Assay and Apoptosis Analysis
3.7. Dual-Luciferase Reporter Assay
3.8. Western Blot
3.9. Statistical Analysis
4. Conclusions
Supplementary Information
ijms-15-06314-s001.pdfAcknowledgments
Conflicts of Interest
References
- Vincent, A.; Herman, J.; Schulick, R.; Hruban, R.H.; Goggins, M. Pancreatic cancer. Lancet 2011, 378, 607–620. [Google Scholar]
- Hidalgo, M. Pancreatic cancer. N. Engl. J. Med 2010, 362, 1605–1617. [Google Scholar]
- Feng, M.; Gao, W.; Wang, R.; Chen, W.; Man, Y.G.; Figg, W.D.; Wang, X.W.; Dimitrov, D.S.; Ho, M. Therapeutically targeting glypican-3 via a conformation-specific single-domain antibody in hepatocellular carcinoma. Proc. Natl. Acad. Sci. USA 2013, 110, E1083–E1091. [Google Scholar]
- Aikawa Aikawa, T.; Whipple, C.A.; Lopez, M.E.; Gunn, J.; Young, A.; Lander, A.D.; Korc, M. Glypican-1 modulates the angiogenic and metastatic potential of human and mouse cancer cells. J. Clin. Investig 2008, 118, 89–99. [Google Scholar]
- Duan, L.; Hu, X.Q.; Feng, D.Y.; Lei, S.Y.; Hu, G.H. GPC-1 may serve as a predictor of perineural invasion and a prognosticator of survival in pancreatic cancer. Asian J. Surg./Asian Surg. Assoc 2013, 36, 7–12. [Google Scholar]
- Mounajjed, T.; Zhang, L.; Wu, T.T. Glypican-3 expression in gastrointestinal and pancreatic epithelial neoplasms. Hum. Pathol 2013, 44, 542–550. [Google Scholar]
- Lee, S.; Reha, J.L.; Tzeng, C.W.; Massarweh, N.N.; Chang, G.J.; Hetz, S.P.; Fleming, J.B.; Lee, J.E.; Katz, M.H. Race does not impact pancreatic cancer treatment and survival in an equal access federal health care system. Ann. Surg. Oncol 2013. [Google Scholar] [CrossRef]
- Whipple, C.A.; Young, A.L.; Korc, M. A. KrasG12D-driven genetic mouse model of pancreatic cancer requires glypican-1 for efficient proliferation and angiogenesis. Oncogene 2012, 31, 2535–2544. [Google Scholar]
- Jalvy-Delvaille, S.; Maurel, M.; Majo, V.; Pierre, N.; Chabas, S.; Combe, C.; Rosenbaum, J.; Sagliocco, F.; Grosset, C.F. Molecular basis of differential target regulation by miR-96 and miR-182: The Glypican-3 as a model. Nucleic Acids Res 2012, 40, 1356–1365. [Google Scholar]
- Moskwa, P.; Buffa, F.M.; Pan, Y.; Panchakshari, R.; Gottipati, P.; Muschel, R.J.; Beech, J.; Kulshrestha, R.; Abdelmohsen, K.; Weinstock, D.M.; et al. miR-182-mediated downregulation of BRCA1 impacts DNA repair and sensitivity to PARP inhibitors. Mol. Cell 2011, 41, 210–220. [Google Scholar]
- Hwang, H.W.; Mendell, J.T. MicroRNAs in cell proliferation, cell death, and tumorigenesis. Br. J. Cancer 2006, 94, 776–780. [Google Scholar]
- Lee, R.C.; Ambros, V. An extensive class of small RNAs in Caenorhabditis elegans. Science 2001, 294, 862–864. [Google Scholar]
- Tsuchiyama, K.; Ito, H.; Taga, M.; Naganuma, S.; Oshinoya, Y.; Nagano, K.; Yokoyama, O.; Itoh, H. Expression of microRNAs associated with Gleason grading system in prostate cancer: miR-182-5p is a useful marker for high grade prostate cancer. Prostate 2013, 73, 827–834. [Google Scholar]
- Guttilla, I.K.; White, B.A. Coordinate regulation of FOXO1 by miR-27a, miR-96, and miR-182 in breast cancer cells. J. Biol. Chem 2009, 284, 23204–23216. [Google Scholar]
- Guo, Y.; Liu, H.; Zhang, H.; Shang, C.; Song, Y. miR-96 regulates FOXO1-mediated cell apoptosis in bladder cancer. Oncol. Lett 2012, 4, 561–565. [Google Scholar]
- Weeraratne, S.D.; Amani, V.; Teider, N.; Pierre-Francois, J.; Winter, D.; Kye, M.J.; Sengupta, S.; Archer, T.; Remke, M.; Bai, A.H.; et al. Pleiotropic effects of miR-183~96~182 converge to regulate cell survival, proliferation and migration in medulloblastoma. Acta Neuropathol 2012, 123, 539–552. [Google Scholar]
- Tanaka, M.; Suzuki, H.I.; Shibahara, J.; Kunita, A.; Isagawa, T.; Yoshimi, A.; Kurokawa, M.; Miyazono, K.; Aburatani, H.; Ishikawa, S.; et al. EVI1 oncogene promotes KRAS pathway through suppression of microRNA-96 in pancreatic carcinogenesis. Oncogene 2013. [Google Scholar] [CrossRef]
- Yu, S.; Lu, Z.; Liu, C.; Meng, Y.; Ma, Y.; Zhao, W.; Liu, J.; Yu, J.; Chen, J. miRNA-96 suppresses KRAS and functions as a tumor suppressor gene in pancreatic cancer. Cancer Res 2010, 70, 6015–6025. [Google Scholar]
- Haflidadottir, B.S.; Larne, O.; Martin, M.; Persson, M.; Edsjo, A.; Bjartell, A.; Ceder, Y. Upregulation of miR-96 Enhances Cellular Proliferation of Prostate Cancer Cells through FOXO1. PLoS One 2013, 8, e72400. [Google Scholar]
- Xu, D.; He, X.; Chang, Y.; Xu, C.; Jiang, X.; Sun, S.; Lin, J. Inhibition of miR-96 expression reduces cell proliferation and clonogenicity of HepG2 hepatoma cells. Oncol. Rep 2013, 29, 653–661. [Google Scholar]
- Zhong, X.Y.; Yu, J.H.; Zhang, W.G.; Wang, Z.D.; Dong, Q.; Tai, S.; Cui, Y.F.; Li, H. MicroRNA-421 functions as an oncogenic miRNA in biliary tract cancer through down-regulating farnesoid X receptor expression. Gene 2012, 493, 44–51. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Characteristics | miR-96-5p | miR-182-5p | |||
|---|---|---|---|---|---|
| Down (%) | Normal/Up (%) | Up (%) | Normal/Down (%) | ||
| Gender | Male | 21 (75.0) | 7 (25.0) | 22 (78.6) | 6 (21.4) |
| Female | 8 (80.0) | 2 (20.0) | 8 (80.0) | 2 (20.0) | |
| Age (years) | <60 | 16 (72.7) | 6 (27.3) | 17 (77.3) | 5 (22.7) |
| ≥60 | 13 (81.2) | 3 (18.8) | 13 (81.3) | 3 (18.7) | |
| Serum CA19-9 | Negative | 5 (71.4) | 2 (28.6) | 5 (71.4) | 2 (28.6) |
| Positive | 24 (77.4) | 7 (22.6) | 25 (80.6) | 6 (19.4) | |
| Tumor size | <2 | 18 (66.7) | 9 (33.3) * | 21 (77.8) | 6 (22.2) |
| ≥2 | 11 (100.0) | 0 (0.0) | 9 (81.8) | 2 (18.2) | |
| Histological differentiation | Well | 14 (63.6) | 8 (36.4) * | 14 (63.6) | 8 (36.4) * |
| Poorly | 15 (93.7) | 1 (6.3) | 16 (100.0) | 0 (0.0) | |
| Pancreatitis | No | 21 (77.8) | 6 (22.2) | 23 (85.2) | 4 (14.8) |
| Yes | 8 (72.7) | 3 (27.3) | 7 (63.6) | 4 (36.4) | |
| Regional lymph node metastasis | No | 23 (74.2) | 8 (25.8) | 24 (77.4) | 7 (22.6) |
| Yes | 6 (85.7) | 1 (14.3) | 6 (85.7) | 1 (14.3) | |
| TNM staging | I-IIA | 23 (74.2) | 8 (25.8) | 24 (77.4) | 7 (22.6) |
| IIB | 6 (85.7) | 1 (14.3) | 6 (85.7) | 1 (14.3) | |
| GPC1 | n | miR-96-5p | Correlation | miR-182-5p | Correlation | ||||
|---|---|---|---|---|---|---|---|---|---|
| down | up/Normal | r | p | up | Normal/down | r | p | ||
| Normal/underexpression(−/+) | 13 | 7 | 6 | −0.381 | 0.018 | 11 | 2 | 0.100 | 0.549 |
| Overexpression(++/+++) | 25 | 22 | 3 | 19 | 6 | ||||
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, C.; Du, X.; Tai, S.; Zhong, X.; Wang, Z.; Hu, Z.; Zhang, L.; Kang, P.; Ji, D.; Jiang, X.; et al. GPC1 Regulated by miR-96-5p, Rather than miR-182-5p, in Inhibition of Pancreatic Carcinoma Cell Proliferation. Int. J. Mol. Sci. 2014, 15, 6314-6327. https://doi.org/10.3390/ijms15046314
Li C, Du X, Tai S, Zhong X, Wang Z, Hu Z, Zhang L, Kang P, Ji D, Jiang X, et al. GPC1 Regulated by miR-96-5p, Rather than miR-182-5p, in Inhibition of Pancreatic Carcinoma Cell Proliferation. International Journal of Molecular Sciences. 2014; 15(4):6314-6327. https://doi.org/10.3390/ijms15046314
Chicago/Turabian StyleLi, Chunlong, Xuefei Du, Sheng Tai, Xiangyu Zhong, Zhidong Wang, Zhanliang Hu, Lei Zhang, Pengcheng Kang, Daolin Ji, Xingming Jiang, and et al. 2014. "GPC1 Regulated by miR-96-5p, Rather than miR-182-5p, in Inhibition of Pancreatic Carcinoma Cell Proliferation" International Journal of Molecular Sciences 15, no. 4: 6314-6327. https://doi.org/10.3390/ijms15046314