The lethal giant larvae Gene in Tribolium castaneum: Molecular Properties and Roles in Larval and Pupal Development as Revealed by RNA Interference

Abstract
:1. Introduction
2. Results and Discussion
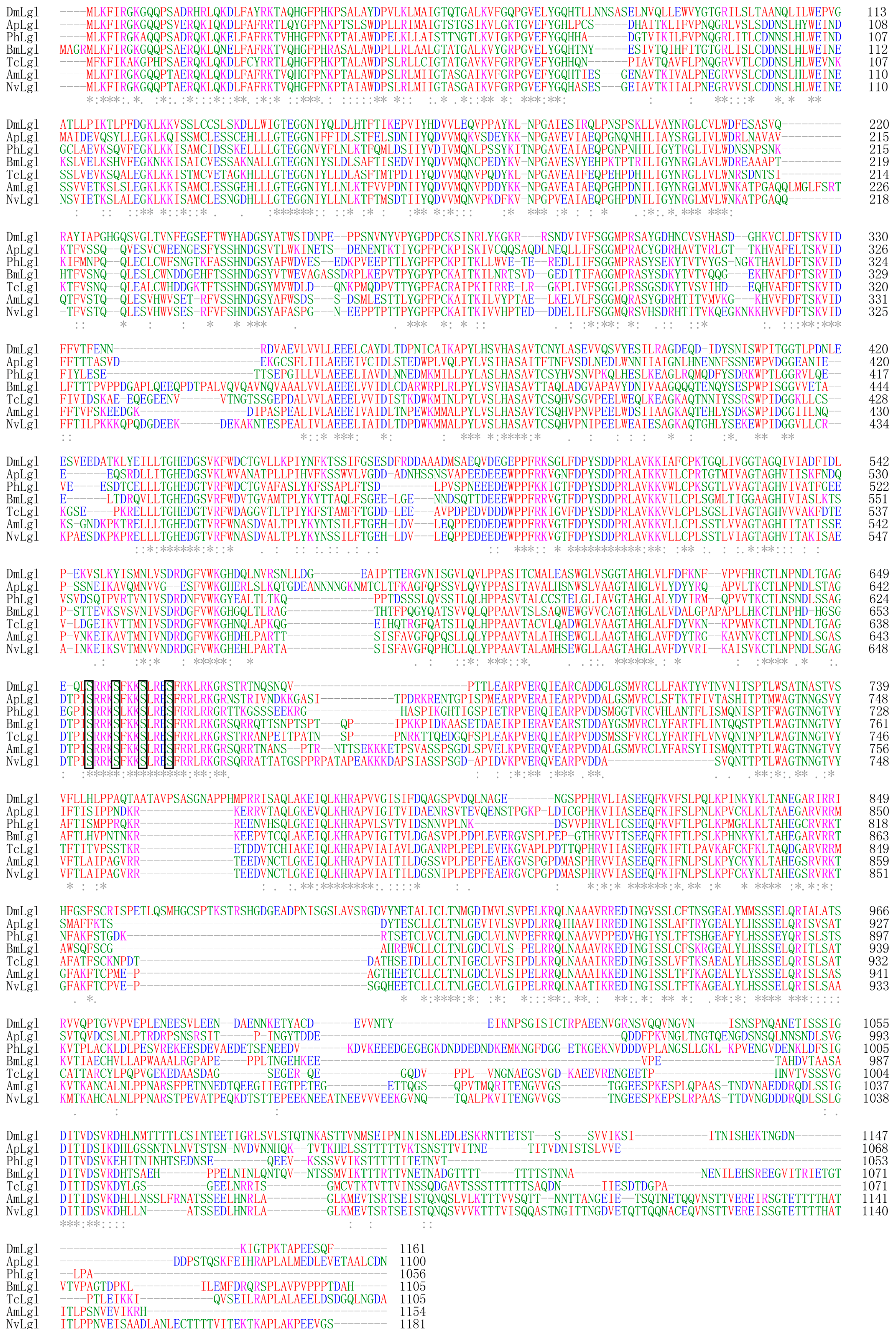
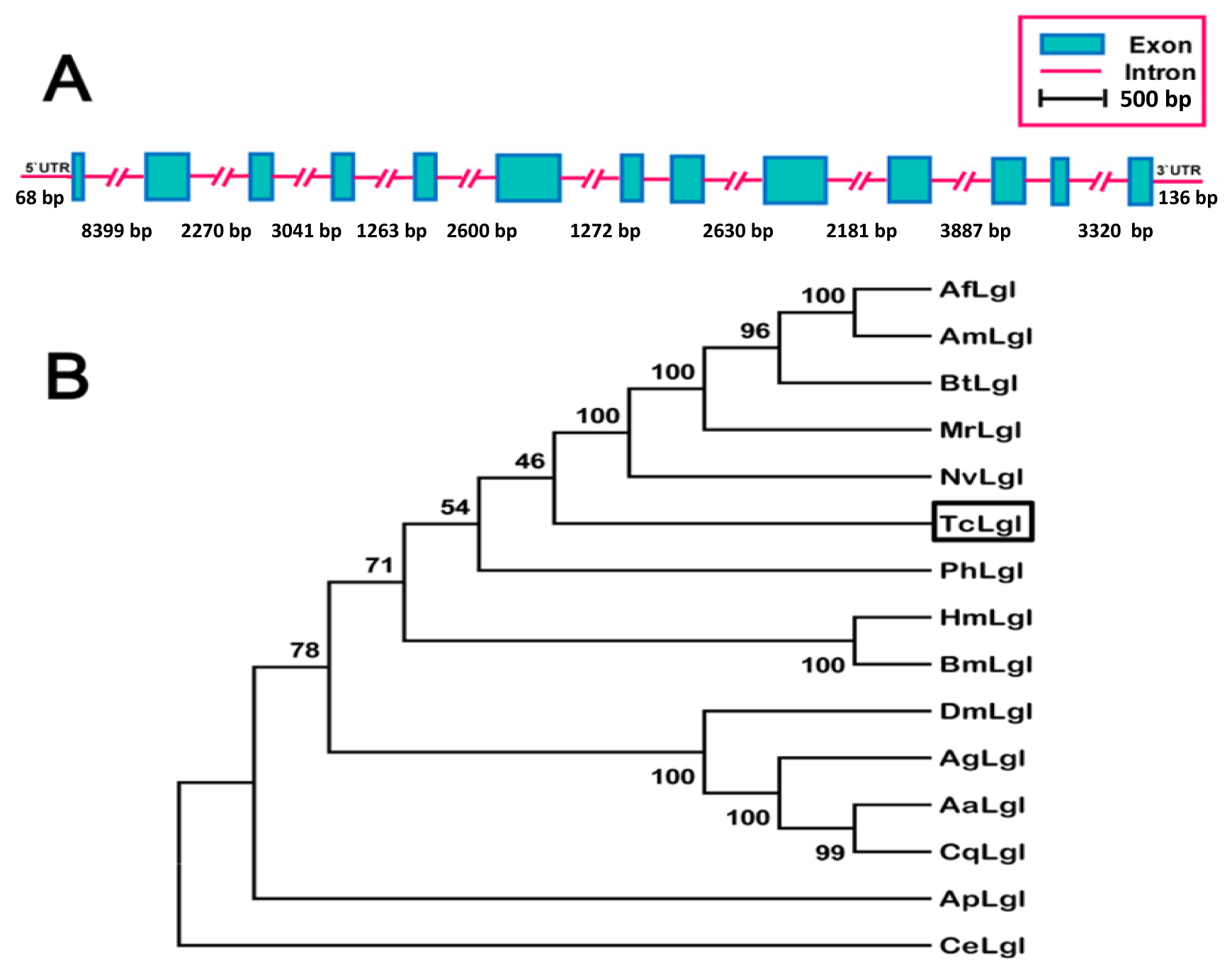
2.1. Analysis of TcLgl cDNA, Deduced Amino Acid, and Genomic Sequences
2.2. Phylogenetic Relationship of T. castaneum Lgl to Other Insect Lgls
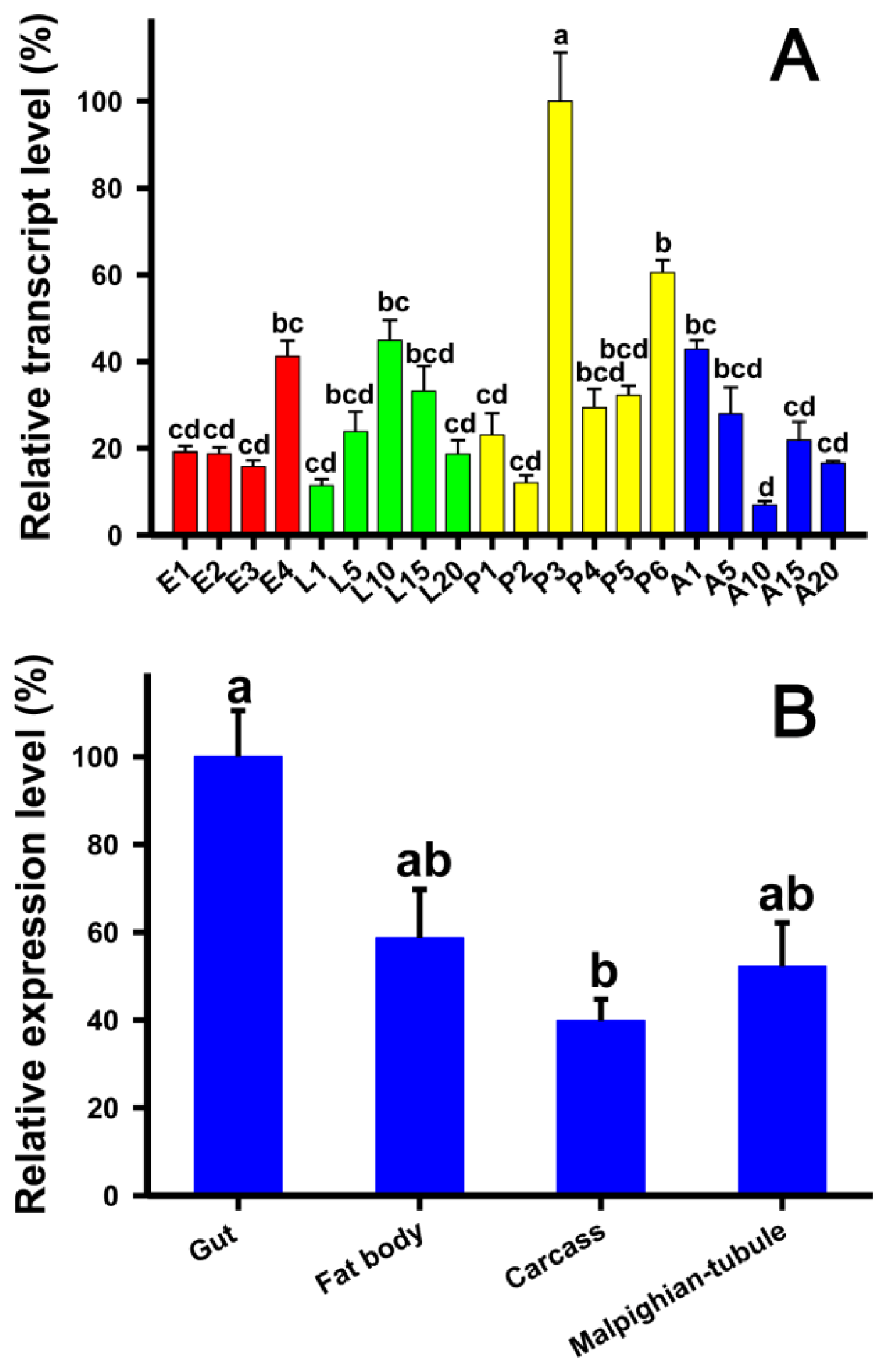
2.3. Developmental Stage- and Tissue-Specific Expression Patterns of TcLgl
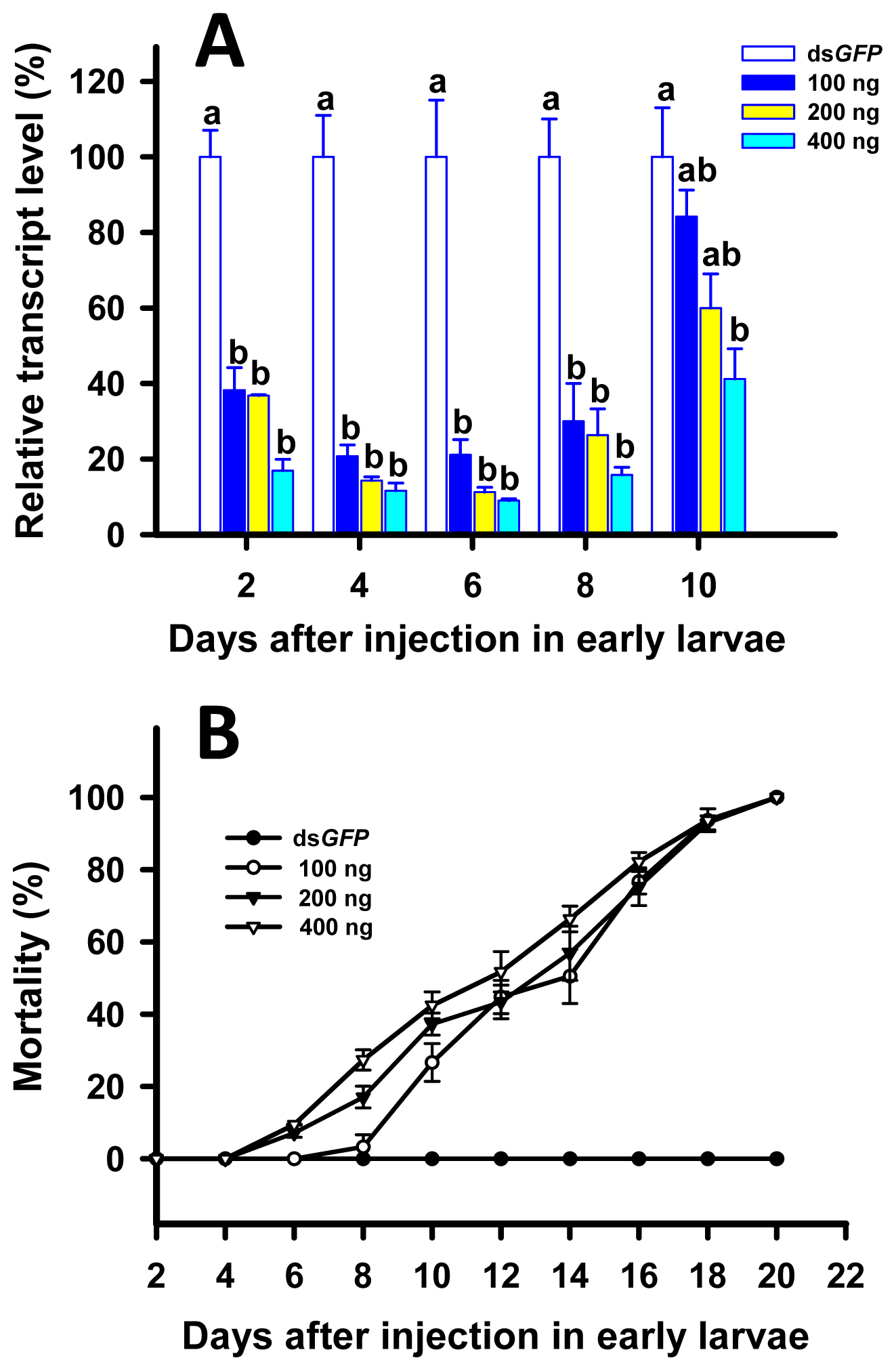
2.4. RNAi of TcLgl in Early (8-Day) Larvae and Its Effect on Survival and Pupation
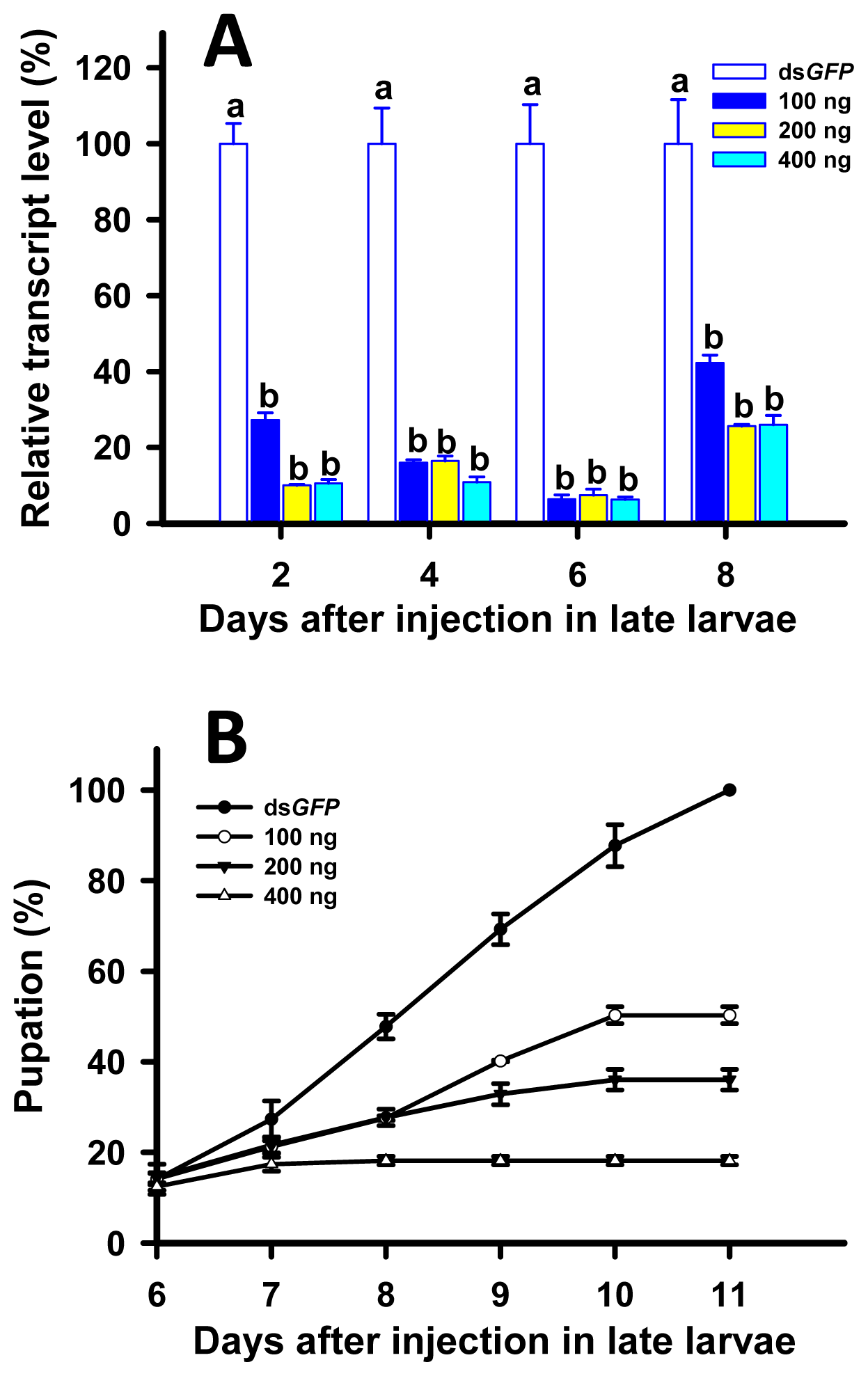
2.5. RNAi of TcLgl in Late (20-Day) Larvae and Its Effect on Survival and Pupation
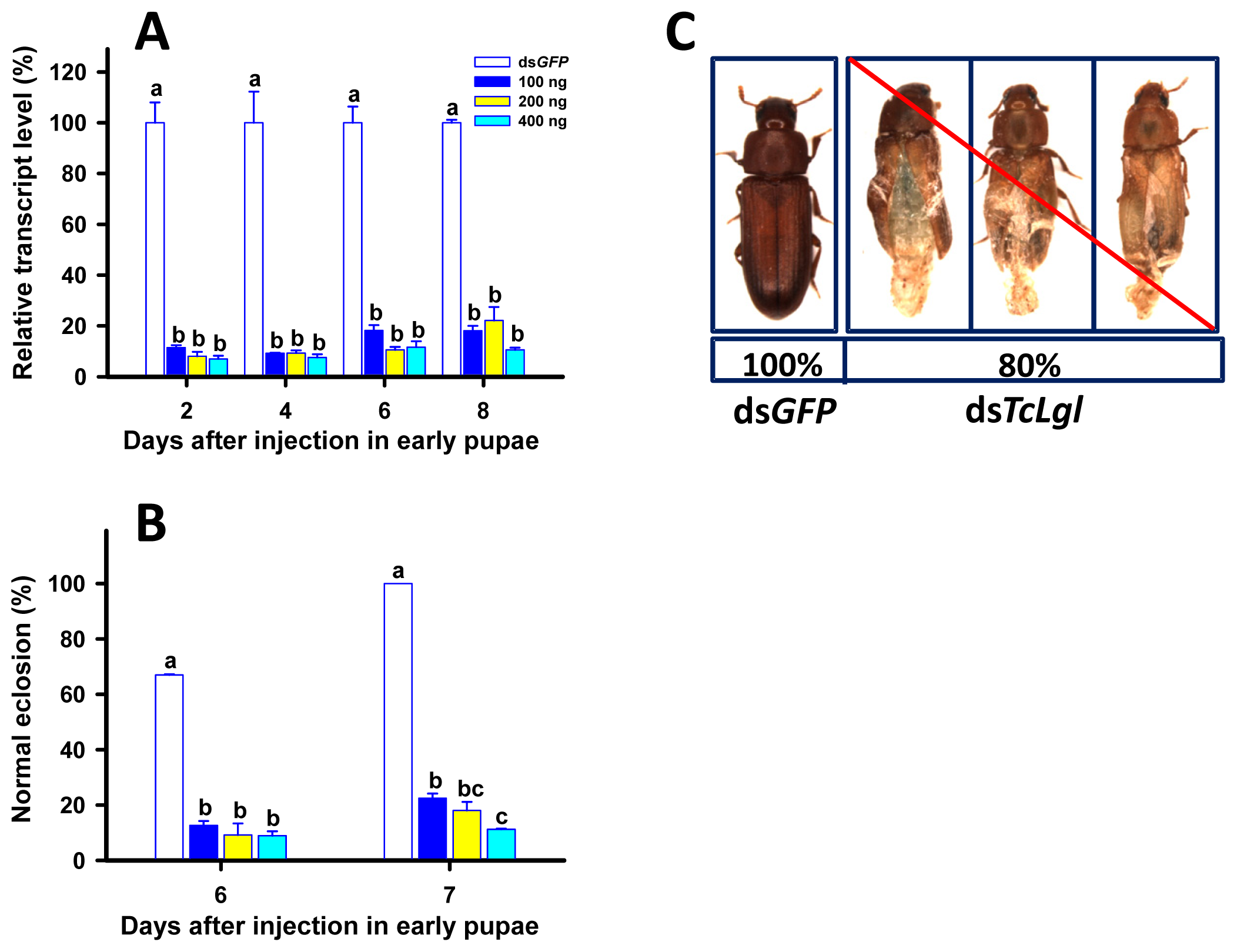
2.6. RNAi of TcLgl in Early (1-Day) Pupae and Its Effect on Survival and Eclosion
2.7. Discussion
3. Experimental Section
3.1. Insect Culture
3.2. Total RNA Isolation and Reverse Transcription
3.3. Subcloning and Sequencing of cDNA
3.4. Analyses of TcLgl cDNA, Deduced Amino Acid, and Genomic Sequences
3.5. Analysis of Expression Profiles by RT-qPCR
3.6. Functional Analysis of TcLgl
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
- Author ContributionsXiao, D., Gao, X., Yao, J. and Zhu, K.Y. conceived and designed the experiments; Xiao, D. and Liang, X. performed the experiments; Xiao, D. and Liang, X. analyzed the data; Zhu, K.Y. contributed reagents/materials/analysis tools; Xiao, D. and Zhu, K.Y. wrote the paper; Xiao, D., Gao, X., Yao, J. and Zhu, K.Y. contributed with revisions.
References
- Tian, A.G.; Deng, W.M. Lgl and its phosphorylation by aPKC regulate oocyte polarity formation inDrosophila. Development 2008, 135, 463–471. [Google Scholar]
- Zhang, X.; Wang, P.; Gangar, A.; Zhang, J.; Brennwald, P.; TerBush, D.; Guo, W. Lethal giant larvae proteins interact with the exocyst complex and are involved in polarized exocytosis. J. Cell Biol 2005, 170, 273–283. [Google Scholar]
- Elsum, I.; Yates, L.; Humbert, P.O.; Richardson, H.E. The Scribble-Dlg-Lgl polarity module in development and cancer: From flies to man. Essays Biochem 2012, 53, 141–168. [Google Scholar]
- Bilder, D.; Li, M.; Perrimon, N. Cooperative regulation of cell polarity and growth by Drosophila tumor suppressors. Science 2000, 289, 113–116. [Google Scholar]
- Klezovitch, O.; Fernandez, T.E.; Tapscott, S.J.; Vasioukhin, V. Loss of cell polarity causes severe brain dysplasia in Lgl1 knockout mice. Genes Dev 2004, 18, 559–571. [Google Scholar]
- Michaelis, U.R.; Chavakis, E.; Kruse, C.; Jungblut, B.; Kaluza, D.; Wandzioch, K.; Manavski, Y.; Heide, H.; Santoni, M.-J.; Potente, M.; et al. The polarity protein Scrib is essential for directed endothelial cell migration. Circ. Res 2013, 112, 924–934. [Google Scholar]
- Vasioukhin, V. Lethal giant puzzle of Lgl. Dev. Neurosci 2006, 28, 13–24. [Google Scholar]
- Staples, J.; Broadie, K. The cell polarity scaffold lethal giant larvae regulates synapse morphology and function. J. Cell Sci 2013, 126, 1992–2003. [Google Scholar]
- Suresh, B.; Ramakrishna, S.; Kim, Y.-S.; Kim, S.-M.; Kim, M.-S.; Baek, K.-H. Stability and function of mammalian lethal giant larvae-1 oncoprotein are regulated by the scaffolding protein RanBPM. J. Biol. Chem 2010, 285, 35340–35349. [Google Scholar]
- Gateff, E. Malignant neoplasms of genetic origin inDrosophila melanogaster. Science 1978, 200, 1448–1459. [Google Scholar]
- Lehman, K.; Rossi, G.; Adamo, J.E.; Brennwald, P. Yeast homologues of tomosyn and lethal giant larvae function in exocytosis and are associated with the plasma membrane SNARE, Sec9. J. Cell Biol 1999, 146, 125–140. [Google Scholar]
- Lützelschwab, R.; Klämbt, C.; Rossa, R.; Schmidt, O. A protein product of the Drosophila recessive tumor gene, 1(2) giant gl, potentially has cell adhesion properties. EMBO J 1987, 6, 1791–1797. [Google Scholar]
- Strand, D.; Raska, I.; Mechler, B.M. The Drosophila lethal (2) giant larvae tumor suppressor protein is a component of the cytoskeleton. J. Cell Biol 1994, 127, 1345–1360. [Google Scholar]
- Jacob, L.; Opper, M.; Metzroth, B.; Phannavong, B.; Mechler, B.M. Structure of the l(2)gl gene of Drosophila and delimitation of its tumor suppressor domain. Cell 1987, 50, 215–225. [Google Scholar]
- Manfruelli, P.; Arquier, N.; Hanratty, W.P.; Sémériva, M. The tumor suppressor gene, lethal(2)giant larvae (l(2)gl), is required for cell shape change of epithelial cells during Drosophila development. Development 1996, 122, 2283–2294. [Google Scholar]
- Baek, K.-H.; Hanratty, W.P. The lethal (2) giant larvae (l(2)gl), a recessive oncogene is required during embryonic and post-embryonic development inDrosophila. Cancer Lett 1997, 111, 233–238. [Google Scholar]
- Arquier, N.; Perrin, L.; Manfruelli, P.; Sémériva, M. The Drosophila tumor suppressor gene lethal (2) giant larvae is required for the emission of the Decapentaplegic signal. Development 2001, 128, 2209–2220. [Google Scholar]
- Betschinger, J.; Eisenhaber, F.; Knoblich, J.A. Phosphorylation-induced autoinhibition regulates the cytoskeletal protein lethal (2) giant larvae. Curr. Biol 2005, 15, 276–282. [Google Scholar]
- Fichelson, P.; Jagut, M.; Lepanse, S.; Lepesant, J.A.; Huynh, J.R. lethal giant larvae is required with the par genes for the early polarization of the Drosophila oocyte. Development 2010, 137, 815–824. [Google Scholar]
- Li, Q.; Feng, S.; Yu, L.; Zhao, G.; Li, M. Requirements of Lgl in cell differentiation and motility during Drosophila ovarian follicular epithelium morphogenesis. Fly 2011, 5, 81–87. [Google Scholar]
- Peng, C.Y.; Manning, L.; Albertson, R.; Doe, C.Q. The tumour-suppressor genes lgl and dlg regulate basal protein targeting in Drosophila neuroblasts. Nature 2000, 408, 596–600. [Google Scholar]
- Lorenzo, C.D.; Strand, D.; Mechler, B.M. Requirement of Drosophila l(2)gl function for survival of the germline cells and organization of the follicle cells in a columnar epithelium during oogenesis. Int. J. Dev. Biol 1999, 43, 207–217. [Google Scholar]
- Cao, Y.; Shi, Y.; Qiao, H.; Yang, Y.; Liu, J.; Shi, Y.; Lin, J.; Zhu, G.; Jin, Y. Distribution of lethal giant larvae (Lgl) protein in the tegument and negative impact of siRNA-based gene silencing on worm surface structure and egg hatching inSchistosoma japonicum. Parasitol. Res 2014, 113, 1–9. [Google Scholar]
- Klämbt, C.; Schmidt, O. Developmental expression and tissue distribution of the lethal (2) giant larvae protein ofDrosophila melanogaster. EMBO J 1986, 5, 2955–2961. [Google Scholar]
- Mechler, B.M.; McGinnis, W.; Gehring, W.J. Molecular cloning of lethal(2)giant larvae, a recessive oncogene ofDrosophila melanogaster. EMBO J 1985, 4, 1551–1557. [Google Scholar]
- Merz, R.; Schmidt, M.; Török, I.; Protin, U.; Schuler, G.; Walther, H.-P.; Krieg, F.; Gross, M.; Strand, D.; Mechler, B.M. Molecular action of the l(2)gl tumor suppressor gene ofDrosophila melanogaster. Environ. Health Persp 1990, 88, 163–167. [Google Scholar]
- Gateff, E. Cancer, genes, and development: The Drosophila case. Adv. Cancer Res 1982, 37, 33–74. [Google Scholar]
- Hadorn, E. An accelerating effect of normal ring-glands on puparium formation in lethal larvae ofDrosophila melanogaster. Proc. Natl. Acad. Sci. USA 1937, 23, 478–484. [Google Scholar]
- Aggarwal, S.K.; King, R.C. A comparative study of the ring gland from the wild type and l(2)gl mutantDrosophila melanogaster. J. Morphol 1969, 129, 171–200. [Google Scholar]
- Compute pI/Mw Tool. Available online: http://web.expasy.org/compute_pi/ (accessed on 16 March 2014).
- ClustalW. Available online: http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_clustalw.html (accessed on 16 March 2014).
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol 2011, 28, 2731–2739. [Google Scholar]
- BeetleBase. Tribolium castaneum. 16 March 2014. Available online: http://beetlebase.org/.
- Giulietti, A.; Overbergh, L.; Valckx, D.; Decallonne, B.; Bouillon, R.; Mathieu, C. An overview of real-time quantitative PCR: Applications to quantify cytokine gene expression. Methods 2001, 25, 386–401. [Google Scholar]
- Lu, Y.; Pang, Y.-P.; Park, Y.; Gao, X.; Yao, J.; Zhang, X.; Zhu, K.Y. Genome organization, phylogenies, expression patterns and three-dimensional protein models of two acetylcholinesterase genes from the red flour beetle. PLoS One 2012, 7, e32288. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 2001, 29. [Google Scholar] [CrossRef]
- Togawa, T.; Dunn, W.A.; Emmons, A.C.; Nagao, J.; Willis, J.H. Developmental expression patterns of cuticular protein genes with the R&R Consensus fromAnopheles gambiae. Insect Biochem. Mol. Biol 2008, 38, 508–519. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Application of Primers | Sequence (5′-3′) | Tm (°C) | Product Length (bp) | |
|---|---|---|---|---|
| PCR for cNDA sequence | TcLgl-1-F | TCGGCTGTTTTACCTGTATTTATC | 59.11 | 875 |
| TcLgl-1-R | GCCATCATTATGTGAGCTTGTG | 60.53 | ||
| TcLgl-2-F | GTGCCGTTGAGGCTATTTTT | 59.23 | 1036 | |
| TcLgl-2-R | CCGGCAACAATCAAACTTC | 59.11 | ||
| TcLgl-3-F | AGTACAGCGATGTTTTTCACTGG | 60.58 | 1030 | |
| TcLgl-3-R | CTGGAAGTGGTGCTACCCC | 60.52 | ||
| TcLgl-4-F | CTATACCTTCACGATAACGGTTCC | 60.15 | 1220 | |
| TcLgl-4-R | CTTAACACACAATTAAAAAGTTTTGGT | 58.32 | ||
| dsRNA synthesis | dsGFP(T7)-F | GGATCCTAATACGACTCACTAT | 60.23 | 305 |
| AGGGTGACCACCCTGACCTAC | ||||
| dsGFP(T7)-R | GGATCCTAATACGACTCACTAT | 60.4 | ||
| AGGGTTGATGCCGTTCTTCTGC | ||||
| dsTcLgl(T7)-F | TAATACGACTCACTATAGG | 60.11 | 385 | |
| GGACGTTGCAACACGGATTC | ||||
| dsTcLgl(T7)-R | TAATACGACTCACTATAGG | 60.40 | ||
| GTGTCATCGTAAAGCTTGCCA | ||||
| RT-qPCR | TcLgl(Q)-F | GACGGATGGCTTTTGCTA | 61.6 | 141 |
| TcLgl(Q)-R | CGGCATTCAACTGTCTCT | 61.2 | ||
| TcRps3-F | CCGTCGTATTCGTGAATTGACTT | 61.1 | 130 | |
| TcRps3-R | TCTAAGAGACTCTGCTTGTGCAATG | 61 | ||
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xiao, D.; Liang, X.; Gao, X.; Yao, J.; Zhu, K.Y. The lethal giant larvae Gene in Tribolium castaneum: Molecular Properties and Roles in Larval and Pupal Development as Revealed by RNA Interference. Int. J. Mol. Sci. 2014, 15, 6880-6896. https://doi.org/10.3390/ijms15046880
Xiao D, Liang X, Gao X, Yao J, Zhu KY. The lethal giant larvae Gene in Tribolium castaneum: Molecular Properties and Roles in Larval and Pupal Development as Revealed by RNA Interference. International Journal of Molecular Sciences. 2014; 15(4):6880-6896. https://doi.org/10.3390/ijms15046880
Chicago/Turabian StyleXiao, Da, Xiao Liang, Xiwu Gao, Jianxiu Yao, and Kun Yan Zhu. 2014. "The lethal giant larvae Gene in Tribolium castaneum: Molecular Properties and Roles in Larval and Pupal Development as Revealed by RNA Interference" International Journal of Molecular Sciences 15, no. 4: 6880-6896. https://doi.org/10.3390/ijms15046880