Human Papillomavirus 16 E6 Contributes HIF-1α Induced Warburg Effect by Attenuating the VHL-HIF-1α Interaction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
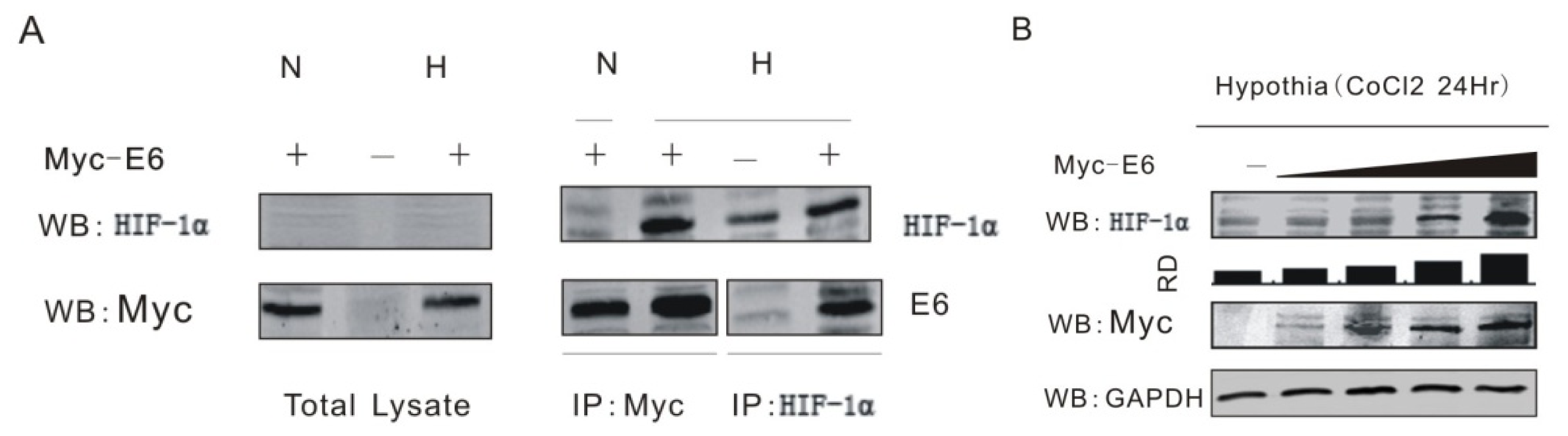
2.1. HPV16 E6 Forms Complex with HIF-1α and Enhances Its Expression
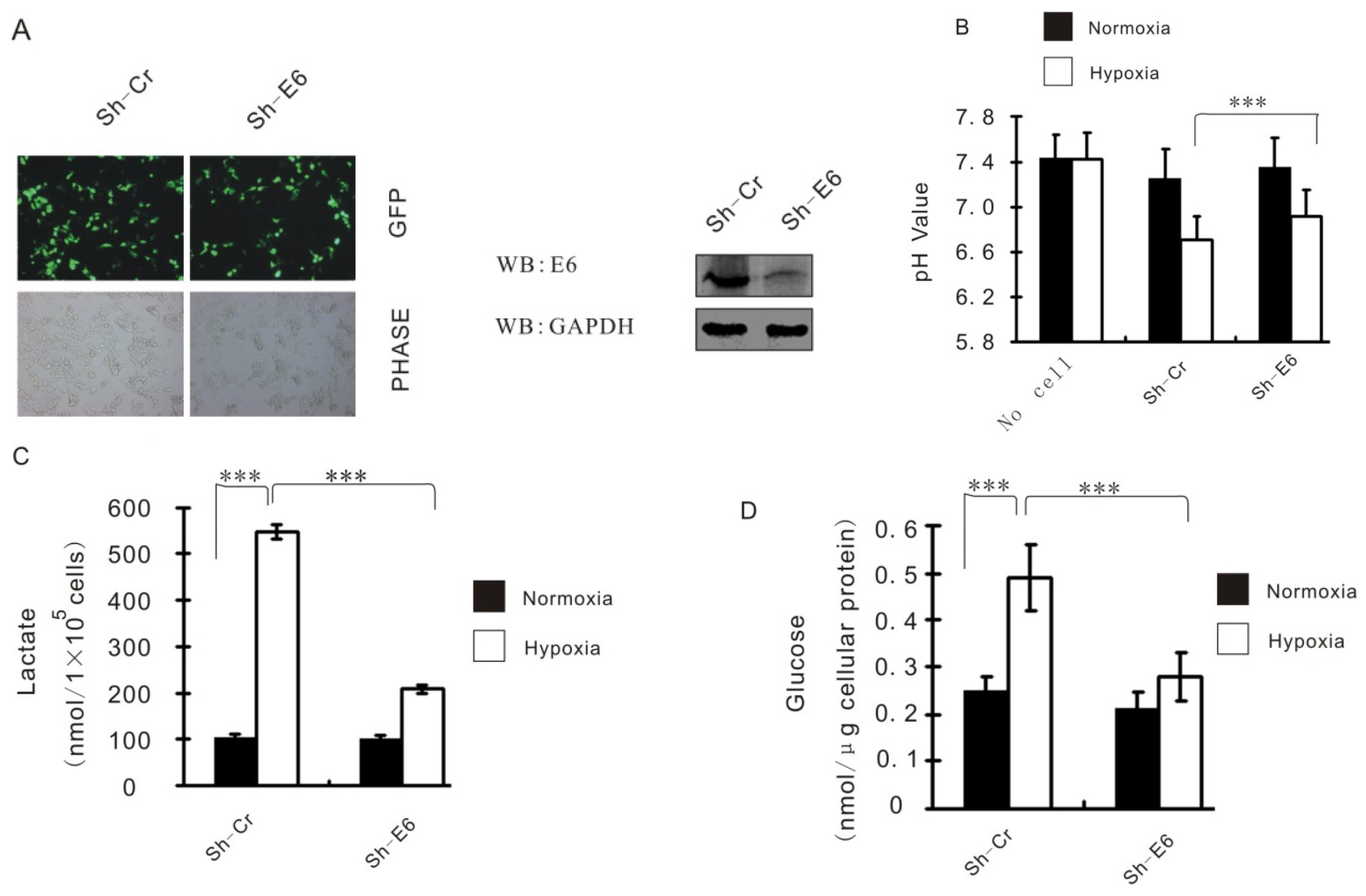
2.2. HPV16 E6 Augments HIF-1α Induced Glycolysis
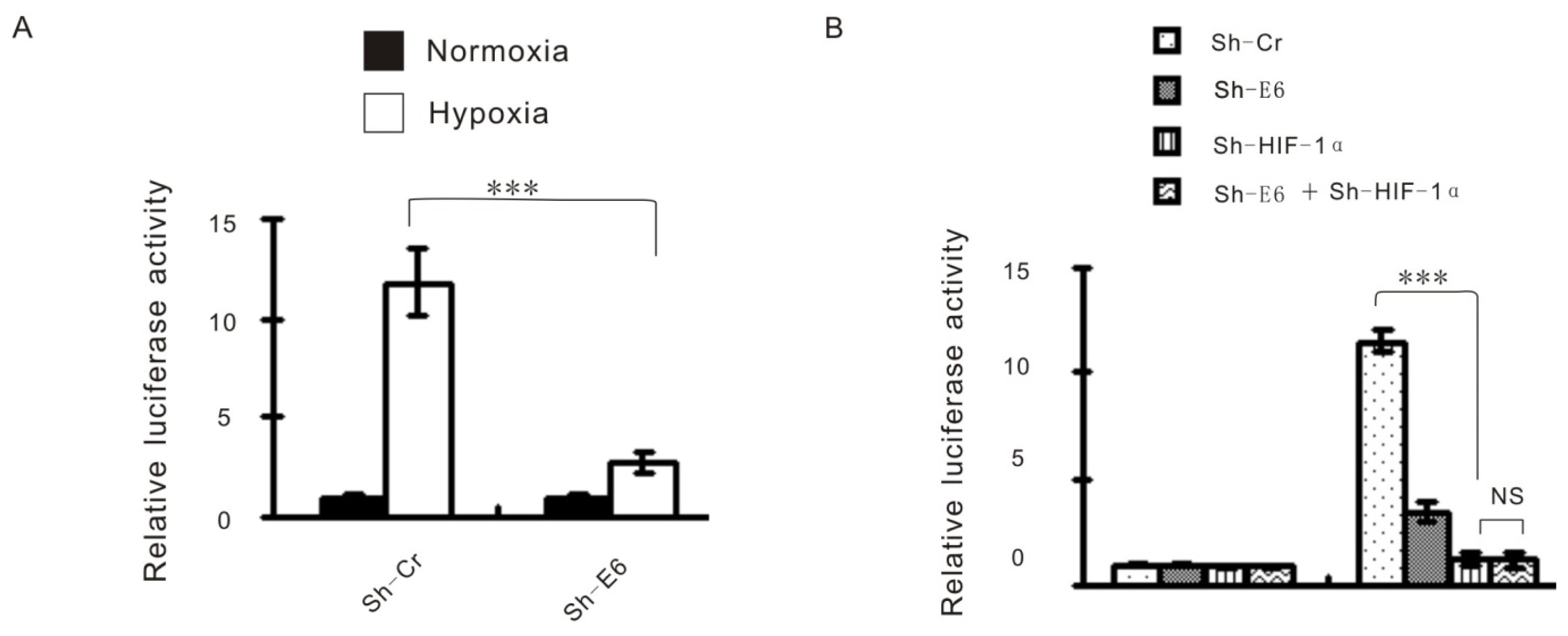
2.3. The Effect of HPV16 E6 on Hypoxia-Enhanced Glycolysis Is HIF-1α Dependent
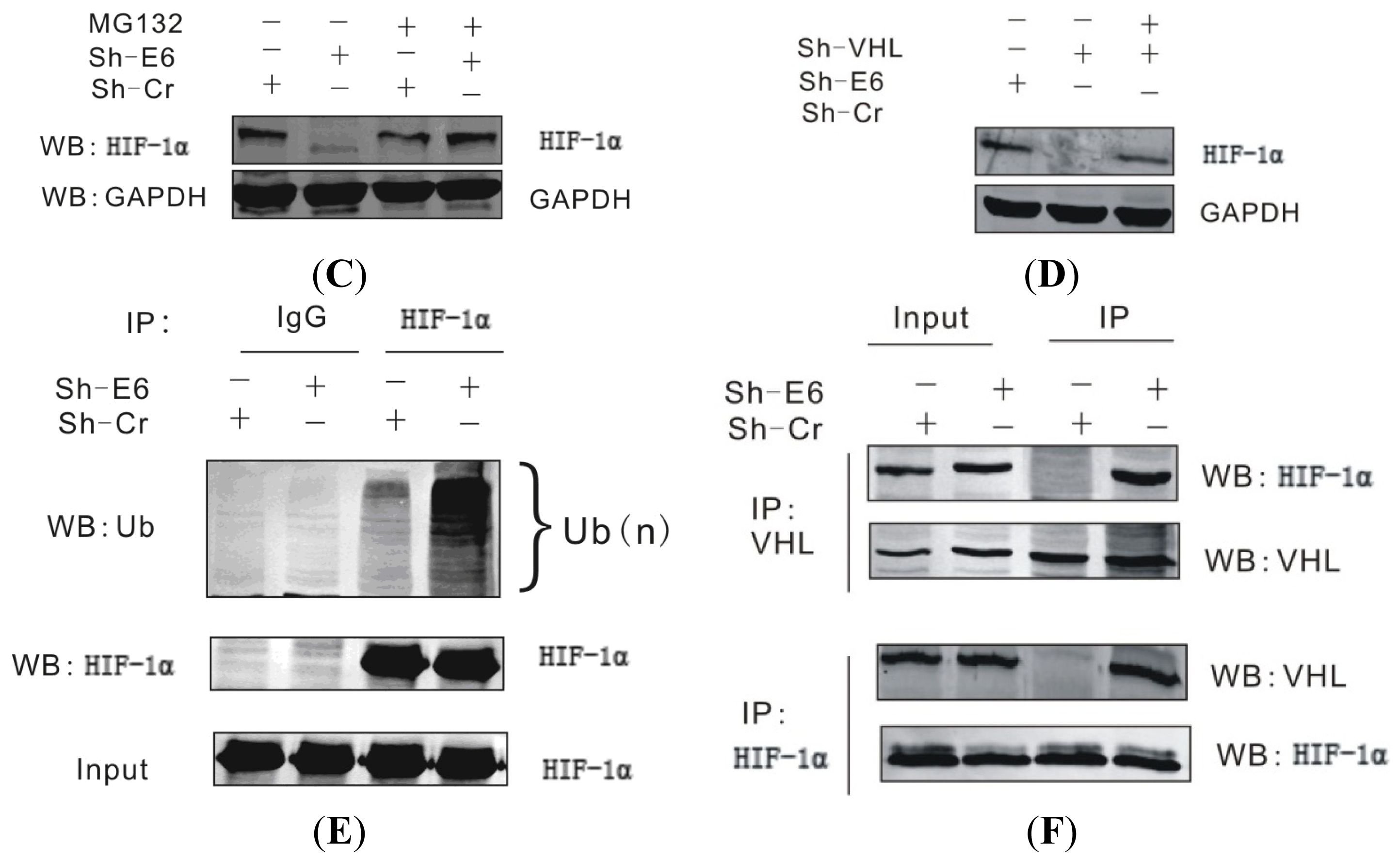
2.4. The HPV16 E6 Stabilizes HIF-1α through Attenuating VHL-HIF-1α Association
2.5. Discussion
3. Experimental Section
3.1. Reagents and Antibodies
3.2. Cell Culture and Transfection
3.3. Immunoprecipitation and Western Blotting
3.4. Glucose Uptake Assay
3.5. Lactate Production Assay
3.6. Luciferase Reporter Assay
3.8. Real-Time PCR
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar]
- Gatenby, R.A.; Gillies, R.J. Why do cancers have high aerobic glycolysis? Nat. Rev.Cancer 2004, 4, 891–899. [Google Scholar]
- Schofield, C.J.; Ratcliffe, P.J. Oxygen sensing by HIF hydroxylases. Nat. Rev. Mol. Cell Biol 2004, 5, 343–354. [Google Scholar]
- Kaelin, W.G., Jr. The von Hippel-Lindau protein, HIF hydroxylation, and oxygen sensing. Biochem. Biophys. Res. Commun 2005, 338, 627–638. [Google Scholar]
- Kaelin, W.G., Jr.; Ratcliffe, P.J. Oxygen sensing by metazoans: The central role of the HIF hydroxylase pathway. Mol. Cell 2008, 30, 393–402. [Google Scholar]
- Kaelin, W.G. Von Hippel-Lindau disease. Annu. Rev. Pathol 2007, 2, 145–173. [Google Scholar]
- Walboomers, J.M.; Jacobs, M.V.; Manos, M.M.; Bosch, F.X.; Kummer, J.A.; Shah, K.V.; Snijders, P.J.; Peto, J.; Meijer, C.J.; Muñoz, N. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J. Pathol 1999, 189, 12–19. [Google Scholar]
- Zur Hausen, H. Papillomaviruses and cancer: From basic studies to clinical application. Nat. Rev. Cancer 2002, 2, 342–350. [Google Scholar]
- Zuna, R.E.; Allen, R.A.; Moore, W.E.; Mattu, R.; Dunn, S.T. Comparison of human papillomavirus genotypes in high-grade squamous intraepithelial lesions and invasive cervical carcinoma: Evidence for differences in biologic potential of precursor lesions. Mod. Pathol 2004, 17, 1314–1322. [Google Scholar]
- Zur Hausen, H. Papillomaviruses causing cancer: Evasion from host-cell control in early events in carcinogenesis. J. Natl. Cancer Inst 2000, 92, 690–698. [Google Scholar]
- Scheffner, M.; Huibregtse, J.M.; Vierstra, R.D.; Howley, P.M. The HPV-16 E6 and E6-AP complex functions as a ubiquitin-protein ligase in the ubiquitination of p53. Cell 1993, 75, 495–505. [Google Scholar]
- Scheffner, M.; Werness, B.A.; Huibregtse, J.M.; Levine, A.J.; Howley, P.M. The E6 oncoprotein encoded by human papillomavirus types 16 and 18 promotes the degradation of p53. Cell 1990, 63, 1129–1136. [Google Scholar]
- Guo, Y.; Meng, X.; Wang, Q.; Wang, Y.; Shang, H. The ING4 Binding with p53 and Induced p53 Acetylation were Attenuated by Human Papillomavirus 16 E6. PLoS One 2013, 8, e71453. [Google Scholar]
- Rodolico, V.; Arancio, W.; Amato, M.C.; Aragona, F.; Cappello, F.; di Fede, O.; Pannone, G.; Campisi, G. Hypoxia inducible factor-1 alpha expression is increased in infected positive HPV16 DNA oral squamous cell carcinoma and positively associated with HPV16 E7 oncoprotein. Infect. Agents Cancer 2011, 6, 18. [Google Scholar] [Green Version]
- Nakamura, M.; Bodily, J.M.; Beglin, M.; Kyo, S.; Inoue, M.; Laimins, L.A. Hypoxia-specific stabilization of HIF-1αlpha by human papillomaviruses. Virology 2009, 387, 442–448. [Google Scholar]
- Semenza, G.L.; Jiang, B.H.; Leung, S.W.; Passantino, R.; Concordet, J.P.; Maire, P.; Giallongo, A. Hypoxia response elements in the aldolase A, enolase 1, and lactate dehydrogenase A gene promoters contain essential binding sites for hypoxia-inducible factor 1. J. Biol. Chem 1996, 271, 32529–32537. [Google Scholar]
- Maxwell, P.H.; Pugh, C.W.; Ratcliffe, P.J. Activation of the HIF pathway in cancer. Curr. Opin. Genet. Dev 2001, 11, 293–299. [Google Scholar]
- Tang, X.; Zhang, Q.; Nishitani, J.; Brown, J.; Shi, S.; Le, A.D. Overexpression of human papillomavirus type 16 oncoproteins enhances hypoxia-inducible factor 1 alpha protein accumulation and vascular endothelial growth factor expression in human cervical carcinoma cells. Clin. Cancer Res 2007, 13, 2568–2576. [Google Scholar]
- Hockel, M.; Schlenger, K.; Hockel, S.; Aral, B.; Schaffer, U.; Vaupel, P. Tumor hypoxia in pelvic recurrences of cervical cancer. Int. J. Cancer 1998, 79, 365–369. [Google Scholar]
- Talks, K.L.; Turley, H.; Gatter, K.C.; Maxwell, P.H.; Pugh, C.W.; Ratcliffe, P.J.; Harris, A.L. The expression and distribution of the hypoxia-inducible factors HIF-1alpha and HIF-2alpha in normal human tissues, cancers, and tumor-associated macrophages. Am. J. Pathol 2000, 157, 411–421. [Google Scholar]
- Semenza, G.L. Defining the role of hypoxia-inducible factor 1 in cancer biology and therapeutics. Oncogene 2010, 29, 625–634. [Google Scholar]
- Bardos, J.I.; Ashcroft, M. Negative and positive regulation of HIF-1: A complex network. Biochim. Biophys. Acta 2005, 1755, 107–120. [Google Scholar]
- Brat, D.J.; Kaur, B.; van Meir, E.G. Genetic modulation of hypoxia induced gene expression and angiogenesis: Relevance to brain tumors. Front. Biosci 2003, 8, 100–116. [Google Scholar]
- Chung, Y.L.; Nagy, E.; Zietkowski, D.; Payne, G.S.; Phillips, D.H.; deSouza, N.M. Molecular and metabolic consequences following E6 transfection in an isogenic ovarian cell line (A2780) pair. Cell. Physiol. Biochem 2013, 32, 1460–1472. [Google Scholar]






© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Guo, Y.; Meng, X.; Ma, J.; Zheng, Y.; Wang, Q.; Wang, Y.; Shang, H. Human Papillomavirus 16 E6 Contributes HIF-1α Induced Warburg Effect by Attenuating the VHL-HIF-1α Interaction. Int. J. Mol. Sci. 2014, 15, 7974-7986. https://doi.org/10.3390/ijms15057974
Guo Y, Meng X, Ma J, Zheng Y, Wang Q, Wang Y, Shang H. Human Papillomavirus 16 E6 Contributes HIF-1α Induced Warburg Effect by Attenuating the VHL-HIF-1α Interaction. International Journal of Molecular Sciences. 2014; 15(5):7974-7986. https://doi.org/10.3390/ijms15057974
Chicago/Turabian StyleGuo, Yi, Xiangkai Meng, Jiaming Ma, Yahong Zheng, Qian Wang, Yanan Wang, and Hong Shang. 2014. "Human Papillomavirus 16 E6 Contributes HIF-1α Induced Warburg Effect by Attenuating the VHL-HIF-1α Interaction" International Journal of Molecular Sciences 15, no. 5: 7974-7986. https://doi.org/10.3390/ijms15057974