Sperm-Egg Fusion: A Molecular Enigma of Mammalian Reproduction


Abstract
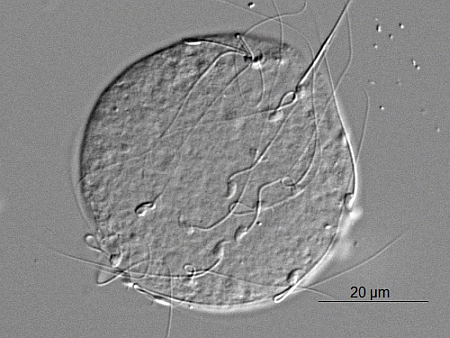
:
{kind=link}
{kind=link}
1. Introduction
2. Fusion as a Crucial Biological Event
3. Interaction of Gametes Culminating in Fusion
4. Identified Players in Gamete Fusion in Mouse
4.1. Essential Molecules on the Sperm Side
4.1.1. SLLP1 (Sperm Lyzozyme-Like Acrosomal Protein)
4.1.2. IZUMO1
Proteins Associated with IZUMO1
4.1.3. Integrins and Their Receptors
4.1.4. CD46
4.2. Essential Molecules on the Egg Side
4.2.1. Tetraspanin Family
CD9
CD81
4.2.2. Glycosilphospatidylinositol (GPI) Anchor
4.2.3. Folate Receptor 4—Juno
4.2.4. SAS1B (Sperm Acrosomal SLLP1 Binding)
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Yanagimachi, R. Sperm-egg fusion. Curr. Top. Membr. Transp. 1988, 32, 3–43. [Google Scholar]
- Bianchi, E.; Doe, B.; Goulding, D.; Wright, G. Juno is the egg Izumo receptor and is essential for mammalian fertilization. Nature 2014, 508, 483–487. [Google Scholar] [CrossRef]
- Inoue, N.; Ikawa, M.; A. Isotani, A.; Okabe, M. The immunoglobulin superfamily protein Izumo is required for sperm to fuse with eggs. Nature 2005, 434, 234–238. [Google Scholar] [CrossRef]
- Ellerman, D.A.; Pei, J.; Gupta, S.; Snell, W.J.; Myles, D.; Primakoff, P. Izumo is part of a multiprotein family whose members form large complexes on mammalian sperm. Mol. Reprod. Dev. 2009, 76, 1188–11995. [Google Scholar] [CrossRef]
- Jahn, R.; Lang, T.; Südhof, T.C. Membrane fusion. Cell 2003, 112, 519–533. [Google Scholar] [CrossRef]
- Stein, K.K.; Primakoff, P.; Myles, D. Sperm-egg fusion: Events at the plasma membrane. J. Cell Sci. 2004, 117, 6269–6274. [Google Scholar] [CrossRef]
- Aguilar, P.S.; Baylies, M.K.; Fleissner, A.; Helming, L.; Inoue, N.; Podbilewicz, B.; Wang, H.; Wong, M. Genetic basis of cell-cell fusion mechanisms. Trend Genet. 2013, 29, 427–437. [Google Scholar] [CrossRef]
- Dimitrov, D.S. Virus entry: Molecular mechanisms and biomedical applications. Nat. Rev. Microbiol. 2004, 2, 109–122. [Google Scholar] [CrossRef]
- Taylor, M.V. Muscle differentiation: How two cells become one. Curr. Biol. 2002, 12, 224–228. [Google Scholar] [CrossRef]
- Oren-Suisse, M.; Podbilewicz, B. Cell fusion during development. Trends Cell Biol. 2007, 17, 537–546. [Google Scholar] [CrossRef]
- Chan, D.C.; Kim, P.S. HIV entry and its inhibition. Cell 1998, 93, 681–684. [Google Scholar]
- Harrison, S.C. Viral membrane fusion. Nat. Struct. Mol. Biol. 2008, 15, 690–698. [Google Scholar] [CrossRef]
- Weber, T.; Zemelman, B.V.; McNew, J.A.; Westermann, B.; Gmachl, M.; Parlati, F.; Söllner, T.H.; Rothman, J.E. SNAREpins: Minimal machinery for membrane fusion. Cell 1998, 92, 759–772. [Google Scholar] [CrossRef]
- Rizo, J.; Rosenmund, C. Synaptic vesicle fusion. Nat. Struct. Mol. Biol. 2008, 15, 665–674. [Google Scholar] [CrossRef]
- Dupressoir, A.; Marceau, G.; Vernochet, C.; Bénit, L.; Kanellopoulos, C.; Sapin, V.; Heidmann, T. Syncytin-A and syncytin-B, two fusogenic placenta-specific murine envelope genes of retroviral origin conserved in Muridae. Proc. Natl. Acad. Sci. USA 2005, 102, 725–730. [Google Scholar]
- Chen, A.; Leikina, E.; Melikov, K.; Podbilewicz, B.; Kozlov, M.M.; Chernomordik, L.V. Fusion-pore expansion during syncytium formation is restricted by an actin network. J. Cell Sci. 2008, 121, 3619–3628. [Google Scholar] [CrossRef]
- White, J.M.; Delos, S.E.; Brecher, M.; Schornberg, K. Structures and mechanisms of viral membrane fusion proteins: Multiple variations on a common theme. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 189–219. [Google Scholar] [CrossRef]
- Zuccotti, M.; Piccinelli, A.; Rossi, P.G.; Garagna, S.; Redi, C.A. Chromatin organization during mouse oocyte growth. Mol. Reprod. Dev. 1995, 41, 479–485. [Google Scholar] [CrossRef]
- Tash, J.S.; Means, A.R. Cyclic adenosine 3',5' monophosphate, calcium and protein phosphorylation in flagellar motility. Biol. Reprod. 1983, 28, 75–104. [Google Scholar] [CrossRef]
- Sebkova, N.; Ded, L.; Vesela, K.; Dvorakova-Hortova, K. Progress of sperm IZUMO1 relocation during spontaneous acrosome reaction. Reproduction 2014, 147, 231–240. [Google Scholar] [CrossRef]
- Jin, M.; Fujiwara, E.; Kakiuchi, Y.; Okabe, M.; Satouh, Y.; Baba, S.A.; Chiba, K.; Hirohashi, N. Most fertilizing mouse spermatozoa begin their acrosome reaction before contact with the zona pellucida during in vitro fertilization. Proc. Natl Acad. Sci. USA 2011, 108, 4892–4896. [Google Scholar] [CrossRef]
- Yanagimachi, R.; Noda, Y.D. Physiological changes in the post-nuclear cap region of mammalian spermatozoa: A necessary preliminary to the membrane fusion between sperm and egg cells. J. Ultrastruct. Res. 1970, 31, 486–493. [Google Scholar] [CrossRef]
- Johnson, M.H.; Eager, D.; Muggleton-Harris, A.; Grave, H.M. Mosaicism in organisation concanavalin A receptors on surface membrane of mouse egg. Nature 1975, 257, 321–322. [Google Scholar] [CrossRef]
- Ebensperger, C.; Barros, C. Changes at the hamster oocyte surface from the germinal vesicle stage to ovulation. Gamete Res. 1984, 9, 387–397. [Google Scholar] [CrossRef]
- Primakoff, P.; Myles, D.G. Cell-cell membrane fusion during mammalian fertilization. FEBS Lett. 2007, 581, 2174–2180. [Google Scholar] [CrossRef]
- He, Z.Y.; Brakebusch, C.; Fässler, R.; Kreidberg, J.A.; Primakoff, P.; Myles, D.G. None of the integrins known to be present on the mouse egg or to be ADAM receptors are essential for sperm-egg binding and fusion. Dev. Biol. 2003, 254, 226–237. [Google Scholar] [CrossRef]
- Herrero, M.B.; Mandal, A.; Digilio, L.C.; Coonrod, S.A.; Maier, B.; Herr, J.C. Mouse SLLP1, a sperm lysozyme-like protein involved in sperm-egg binding and fertilization. Dev. Biol. 2005, 284, 126–142. [Google Scholar] [CrossRef]
- Ohnami, N.; Nakamura, A.; Miyado, M.; Sato, M.; Kawano, N.; Yoshida, K.; Harada, Y.; Takezawa, Y.; Kanai, S.; Ono, C.; et al. CD81 and CD9 work independently as extracellular components upon fusion of sperm and Oocyte. Biol. Open 2012, 1, 640–647. [Google Scholar] [CrossRef]
- Sachdev, M.; Mandal, A.; Mulders, S.; Digilio, L.C.; Panneerdoss, S.; Suryavathi, V.; Pires, E.; Klotz, K.L.; Hermens, L.; Herrero, M.B.; et al. Oocyte specific oolemmal SAS1B involved in sperm binding through intra-acrosomal SLLP1 during fertilization. Dev. Biol. 2012, 363, 40–51. [Google Scholar] [CrossRef]
- Okabe, M.; Yagasaki, M.; Oda, H.; Matzno, S.; Kohama, Y.; Mimura, T. Effect of a monoclonal anti-mouse sperm antibody (OBF13) on the interaction of mouse sperm with zona-free mouse and hamster eggs. J. Reprod. Immunol. 1988, 13, 211–219. [Google Scholar] [CrossRef]
- Inoue, N.; Ikawa, M.; Okabe, M. The mechanism of sperm-egg interaction and the involvement of IZUMO1 in fusion. Asian J. Androl. 2011, 13, 81–87. [Google Scholar] [CrossRef]
- Inoue, N.; Hamada, D.; Kamikubo, H.; Hirata, K.; Kataoka, M.; Yamamoto, M.; Ikawa, M.; Okabe, M.; Hagihara, Y. Molecular dissection of IZUMO1, a sperm protein essential for sperm-egg fusion. Development 2013, 140, 3221–3229. [Google Scholar] [CrossRef]
- Inoue, N.; Kasahara, T.; Ikawa, M.; Okabe, M. Identification and disruption of sperm-specific angiotensin converting enzyme-3 (ACE3) in mouse. PLoS One 2010, 5, e10301. [Google Scholar] [CrossRef]
- Almeida, E.A.; Huovila, A.P.; Sutherland, A.E.; Stephens, L.E.; Calarco, P.G.; Shaw, L.M.; Mercurio, A.M.; Sonnenberg, A.; Primakoff, P.; Myles, D.G.; et al. Mouse egg integrin α6β1 functions as a sperm receptor. Cell 1995, 81, 1095–1104. [Google Scholar]
- Miller, B.J.; Georges-Labouesse, E.; Primakoff, P.; Myles, D.G. Normalfertilizationoccurs with eggslacking the integrinalpha6beta1 and is CD9-dependent. J. Cell Biol. 2000, 149, 1289–1296. [Google Scholar] [CrossRef]
- Barraud-Lange, V.; Naud-Barriant, N.; Saffar, L.; Gattegno, L.; Ducot, B.; Drillet, A.S.; Bomsel, M.; Wolf, J.P.; Ziyyat, A. Alpha6beta1 integrin expressed by sperm is determinant in mouse fertilization. BMC Dev. Biol. 2007, 7. [Google Scholar] [CrossRef]
- Miyado, K.; Yoshida, K.; Yamagata, K.; Sakakibara, K.; Okabe, M.; Wang, X.; Miyamoto, K.; Akutsu, H.; Kondo, T.; Takahashi, Y.; et al. The fusing ability of sperm is bestowed by CD9-containing vesicles released from eggs in mice. Proc. Natl Acad. Sci. USA 2008, 105, 12921–12926. [Google Scholar] [CrossRef]
- Chen, H.; Sampson, N.S. Mediation of sperm-egg fusion: Evidence that mouse egg α6β1 integrin is the receptor for sperm fertilinbeta. Chem. Biol. 1999, 6, 1–10. [Google Scholar] [CrossRef]
- Evans, J.P. Fertilin β and other ADAMs as integrin ligands: Insights into cell adhesion and fertilization. Bioessay 2001, 23, 628–639. [Google Scholar] [CrossRef]
- Baessler, K.A.; Lee, Y.; Sampson, N.S. β1 integrin is an adhesion protein for sperm binding to eggs. ACS Chem. Biol. 2009, 4, 357–366. [Google Scholar] [CrossRef]
- Evans, J.P.; Kopf, G.S.; Schultz, R.M. Characterization of the binding of recombinant mouse sperm fertilin β subunit to mouse eggs: Evidence for adhesive activity via an egg β1integrin-mediated interaction. Dev. Biol. 1997, 187, 79–93. [Google Scholar] [CrossRef]
- Cho, C.; Bunch, D.O.; Faure, J.E.; Goulding, E.H.; Eddy, E.M.; Primakoff, P.; Myles, D.G. Fertilizationdefects in sperm from mice lacking fertilin β. Science 1998, 281, 1857–1859. [Google Scholar]
- Bigler, D.; Takahashi, Y.; Chen, M.S.; Almeida, E.A.; Osbourne, L.; White, J.M. Sequence-specific interaction between the disintegrin domain of mouse ADAM2 (fertilin β) and murine eggs. Role of the α6 integrin subunit. J. Biol. Chem. 2000, 275, 11576–11584. [Google Scholar]
- Yamaguchi, R.; Muro, Y.; Isotani, A.; Tokuhiro, K.; Takumi, K.; Adham, I.; Ikawa, M.; Okabe, M. Disruption of ADAM3 impairs the migration of sperm into oviduct in mouse. Biol. Reprod. 2009, 81, 142–146. [Google Scholar] [CrossRef]
- Nishimura, H.; Cho, C.; Brandiforte, D.R.; Myles, D.G.; Primakoff, P. Analysis of loss of adhesive function in sperm lacking cyritestin or fertilin beta. Dev. Biol. 2001, 233, 204–214. [Google Scholar] [CrossRef]
- Mizuno, M.; Harris, C.L.; Johnson, P.M.; Morgan, B.P. Rat membrane cofactor protein (MCP; CD46) is expressed only in the acrosome of developing and mature spermatozoa and mediates binding to immobilized activated C3. Biol. Reprod. 2004, 71, 1374–1384. [Google Scholar] [CrossRef]
- Taylor, C.T.; Biljan, M.M.; Kingland, C.R.; Johnson, P.M. Inhibition of human spermatozoon-oocyte interaction in vitro by monoclonal antibodies to CD46 (membrane cofactor protein). Hum. Reprod. 1994, 9, 907–911. [Google Scholar]
- Lozahic, S.; Christiansen, D.; Manié, S.; Gerlier, D.; Billard, M.; Boucheix, C.; Rubinstein, E. CD46 (membrane cofactor protein) associates with multiple beta1 integrins and tetraspa. Eur. J. Immunol. 2000, 30, 900–907. [Google Scholar] [CrossRef]
- Inoue, N.; Ikawa, M.; Nakanishi, T.; Matsumoto, M.; Nomura, M.; Seya, T.; Okabe, M. Disruption of mouse CD46 causes an accelerated spontaneous acrosome reaction in sperm. Mol. Cell. Biol. 2003, 23, 2614–2622. [Google Scholar] [CrossRef]
- Johnson, P.M.; Clift, L.E.; Andrlíková, P.; Jursová, M.; Flanagan, B.F.; Cummerson, J.A.; Stopka, P.; Dvořáková-Hortová, K. Rapid sperm acrosome reaction in the absence of acrosomal CD46 expression in promiscuous field mice (Apodemus). Reproduction 2007, 134, 739–747. [Google Scholar] [CrossRef]
- Stipp, C.S.; Kolesnikova, T.V.; Hemler, M.E. Functional domains in tetraspanin proteins. Trends Biochem. Sci. 2003, 28, 106–112. [Google Scholar] [CrossRef]
- Claas, C.; Stipp, C.S.; Hemler, M.E. Evaluation of prototype transmembrane 4 superfamily protein complexes and their relation to lipid rafts. J. Biol. Chem. 2001, 276, 7974–7984. [Google Scholar] [CrossRef]
- Zoller, M. Tetraspanins: Push and pull in suppressing and promoting metastasis. Nat. Rev. Cancer 2009, 9, 40–55. [Google Scholar] [CrossRef]
- Farquhar, M.J.; Harris, H.J.; McKeating, J.A. Hepatitis C virus entry and the tetraspanin CD81. Biochem. Soc. Trans. 2001, 39, 532–536. [Google Scholar] [CrossRef]
- Tachibana, I.; Hemler, M.E. Role of transmembrane 4 superfamily (TM4SF) proteins CD9 and CD81 in muscle cell fusion and myotube maintenance. J. Cell Biol. 1999, 146, 893–904. [Google Scholar] [CrossRef]
- Takeda, Y.; Tachibana, I.; Miyado, K.; Kobayashi, M.; Miyazaki, T.; Funakoshi, T.; Kimura, H.; Yamane, H.; Saito, Y.; Goto, H.; et al. Tetraspanins CD9 and CD81 function to prevent the fusion of mononuclear phagocytes. J. Cell Biol. 2003, 161, 945–956. [Google Scholar] [CrossRef]
- Glazar, A.I.; Evans, J.P. Immunoglobulin superfamily member IgSF8 (EWI-2) and CD9 in fertilisation: Evidence of distinct functions for CD9 and a CD9-associated protein in mammalian sperm-egg interaction. Reprod. Fertil. Dev. 2009, 21, 293–303. [Google Scholar] [CrossRef]
- Le Naour, F.; André, M.; Boucheix, C.; Rubinstein, E. Membrane microdomains and proteomics: Lessons from tetraspanin microdomains and comparison with lipid rafts. Proteomics 2006, 6, 6447–6454. [Google Scholar] [CrossRef]
- Chen, M.S.; Tung, K.S.; Coonrod, S.A.; Takahashi, Y.; Bigler, D.; Chang, A.; Yamashita, Y.; Kincade, P.W.; Herr, J.C.; White, J.M. Role of the integrin-associated protein CD9 in binding between sperm ADAM 2 and the egg integrin α6β1: Implications for murine fertilization. Proc. Natl. Acad. Sci. USA 1999, 96, 11830–11835. [Google Scholar] [CrossRef]
- Miyado, K.; Yamada, G.; Yamada, S.; Hasuwa, H.; Nakamura, Y.; Ryu, F.; Suzuki, K.; Kosai, K.; Inoue, K.; Ogura, A.; et al. Requirement of CD9 on the egg plasma membrane for fertilization. Science 2000, 287, 321–324. [Google Scholar] [CrossRef]
- Le Naour, F.; Rubinstein, E.; Jasmin, C.; Prenant, M.; Boucheix, C. Severely reduced female fertility in CD9-deficient mice. Science 2000, 287, 319–321. [Google Scholar] [CrossRef]
- Kaji, K.; Oda, S.; Shikano, T.; Ohnuki, T.; Uematsu, Y.; Sakagami, J.; Tada, N.; Miyazaki, S.; Kudo, A. The gamete fusion process is defective in eggs of Cd9-deficient mice. Nat. Genet. 2000, 24, 279–282. [Google Scholar] [CrossRef]
- Waterhouse, R.; Ha, C.; Dveksler, G.S. Murine CD9 is the receptor for pregnancy-specific glycoprotein 17. J. Exp. Med. 2002, 195, 277–282. [Google Scholar] [CrossRef]
- Ellerman, D.A.; Ha, C.; Primakoff, P.; Myles, D.G.; Dveksler, G.S. Direct binding of the ligand PSG17 to CD9 requires a CD9 site essential for sperm-egg fusion. Mol. Biol. Cell. 2003, 14, 5098–5103. [Google Scholar] [CrossRef]
- Jégou, A.; Ziyyat, A.; Barraud-Lange, V.; Perez, E.; Wolf, J.P.; Pincet, F.; Gourier, C. CD9 tetraspanin generates fusion competent sites on the egg membrane for mammalian fertilization. Proc. Natl. Acad. Sci. USA 2011, 108, 10946–10951. [Google Scholar]
- Zhu, X.; Evans, J.P. Analysis of the roles of RGD-binding integrins, α4/α9 integrins, α6 integrins, and CD9 in the interaction of the fertilin β (ADAM2) disintegrin domain with the mouse egg membrane. Biol. Reprod. 2002, 66, 1193–1202. [Google Scholar] [CrossRef]
- Runge, K.E.; Evans, J.E.; He, Z.Y.; Gupta, S.; McDonald, K.L.; Stahlberg, H.; Primakoff, P.; Myles, D.G. Oocyte CD9 is enriched on the microvillar membrane and required for normal microvillar shape and distribution. Dev. Biol. 2007, 304, 317–325. [Google Scholar] [CrossRef]
- Horváth, G.; Serru, V.; Clay, D.; Billard, M.; Boucheix, C.; Rubinstein, E. CD19 is linked to the integrin-associated tetraspans CD9, CD81, and CD82. J. Biol. Chem. 1998, 273, 30537–30543. [Google Scholar] [CrossRef]
- Rubinstein, E.; Ziyyat, A.; Prenant, M.; Wrobel, E.; Wolf, J.P.; Levy, S.; le Naour, F.; Boucheix, C. Reduced fertility of female mice lacking CD81. Dev. Biol. 2006, 290, 351–358. [Google Scholar] [CrossRef]
- Pileri, P.; Uematsu, Y.; Campagnoli, S.; Galli, G.; Falugi, F.; Petracca, R.; Weiner, A.J.; Houghton, M.; Rosa, D.; Grandi, G.; et al. Binding of hepatitis C virus to human CD81. Science 1998, 282, 938–941. [Google Scholar] [CrossRef]
- Coonrod, S.A.; Naaby-Hansen, S.; Shetty, J.; Shibahara, H.; Chen, M.; White, J.M.; Herr, J.C. Treatment of mouse oocytes with PI-PLC releases 70-kDa (pI 5) and 35- to 45-kDa (pI 5.5) protein clusters from the egg surface and inhibits sperm-oolemma binding and fusion. Dev. Biol. 1999, 207, 334–349. [Google Scholar] [CrossRef]
- Alfieri, J.A.; Martin, A.D.; Takeda, J.; Kondoh, G.; Myles, D.G.; Primakoff, P. Infertility in female mice with an oocyte-specific knockout of GPI-anchored proteins. J. Cell Sci. 2003, 116, 2149–2155. [Google Scholar]
- Lefèvre, B.; Wolf, J.P.; Ziyyat, A. Sperm-egg interaction: Is there a link between tetraspanin(s) and GPI-anchored protein(s)? Bioessays 2010, 32, 143–152. [Google Scholar] [CrossRef]
- Talbot, P.; Shur, B.D.; Myles, D.G. Cell adhesion and fertilization: Steps in oocyte transport, sperm-zona pellucida interactions, and sperm-egg fusion. Biol. Reprod. 2003, 68, 1–9. [Google Scholar]
- Yamagata, K.; Nakanishi, T.; Ikawa, M.; Yamaguchi, R.; Moss, S.B.; Okabe, M. Sperm from the calmegin-deficient mouse have normal abilities for binding and fusion to the egg plasma membrane. Dev. Biol. 2002, 250, 348–357. [Google Scholar] [CrossRef]
- Inoue, N.; Nishikawa, T.; Ikawa, M.; Okabe, M. Tetraspanin-interacting protein IGSF8 is dispensable for mouse fertility. Fertil. Steril. 2012, 98, 465–470. [Google Scholar] [CrossRef]
- Kaji, K.; Oda, S.; Miyazaki, S.; Kudo, A. Infertility of CD9-deficient mouse eggs is reversed by mouse CD9, human CD9, or mouse CD81; polyadenylated mRNA injection developed for molecular analysis of sperm-egg fusion. Dev. Biol. 2002, 247, 327–334. [Google Scholar] [CrossRef]
- Evans, J.P. Sperm-egg interaction. Annu. Rev. Physiol. 2012, 74, 477–502. [Google Scholar] [CrossRef]
- Grayson, P.; Civetta, A. Positive selection and the evolution of izumo genes in mammals. Int. J. Evol. Biol. 2012. [Google Scholar] [CrossRef]
- Grayson, P.; Civetta, A. Positive selection in the adhesion domain of Mus sperm Adam genes through gene duplication and function-driven gene complex formations. BMC Evol. Biol. 2013, 13. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Klinovska, K.; Sebkova, N.; Dvorakova-Hortova, K. Sperm-Egg Fusion: A Molecular Enigma of Mammalian Reproduction. Int. J. Mol. Sci. 2014, 15, 10652-10668. https://doi.org/10.3390/ijms150610652
Klinovska K, Sebkova N, Dvorakova-Hortova K. Sperm-Egg Fusion: A Molecular Enigma of Mammalian Reproduction. International Journal of Molecular Sciences. 2014; 15(6):10652-10668. https://doi.org/10.3390/ijms150610652
Chicago/Turabian StyleKlinovska, Karolina, Natasa Sebkova, and Katerina Dvorakova-Hortova. 2014. "Sperm-Egg Fusion: A Molecular Enigma of Mammalian Reproduction" International Journal of Molecular Sciences 15, no. 6: 10652-10668. https://doi.org/10.3390/ijms150610652