First Isolation of New Canine Parvovirus 2a from Tibetan Mastiff and Global Analysis of the Full-Length VP2 Gene of Canine Parvoviruses 2 in China



Abstract
:1. Introduction
2. Results
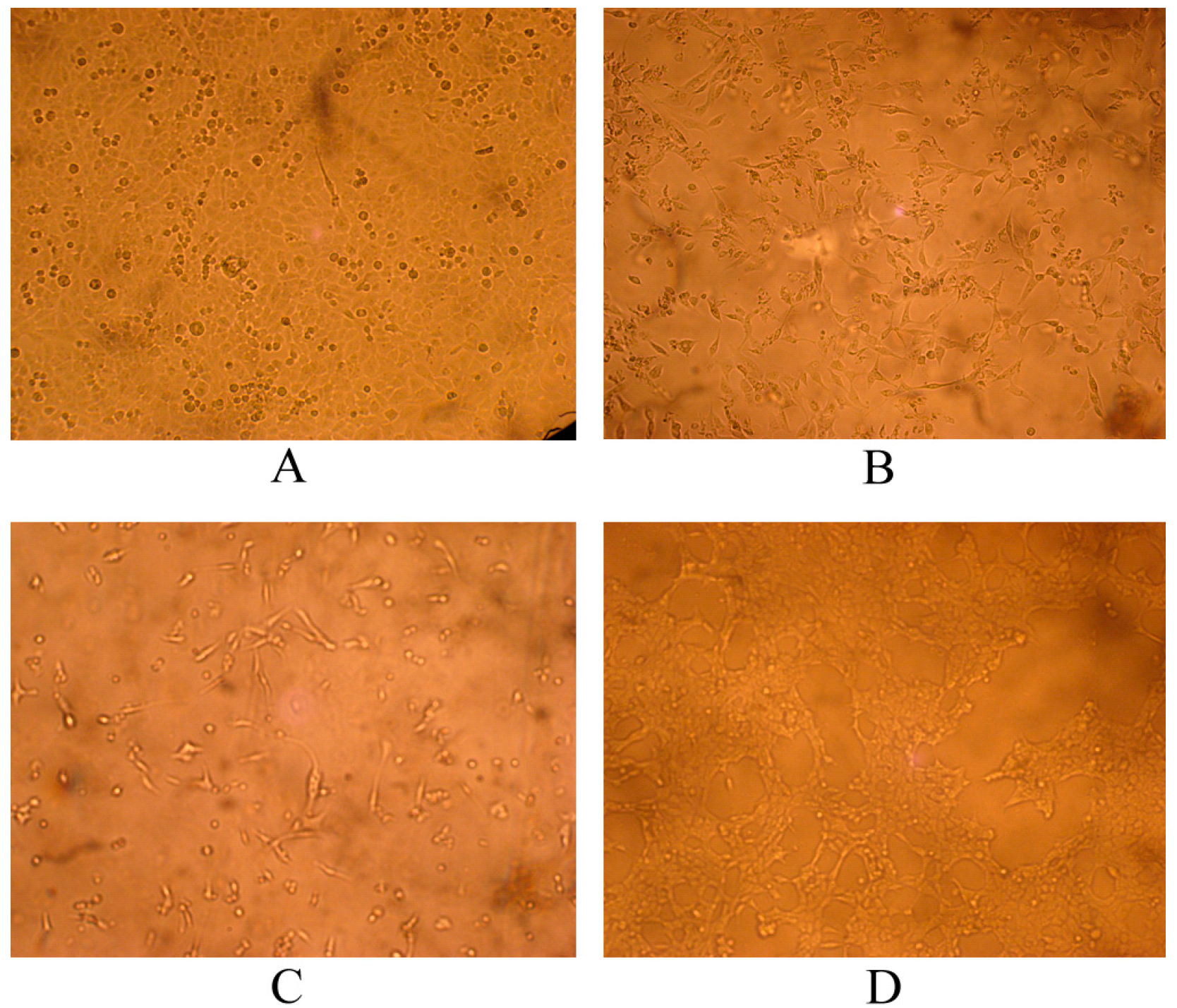
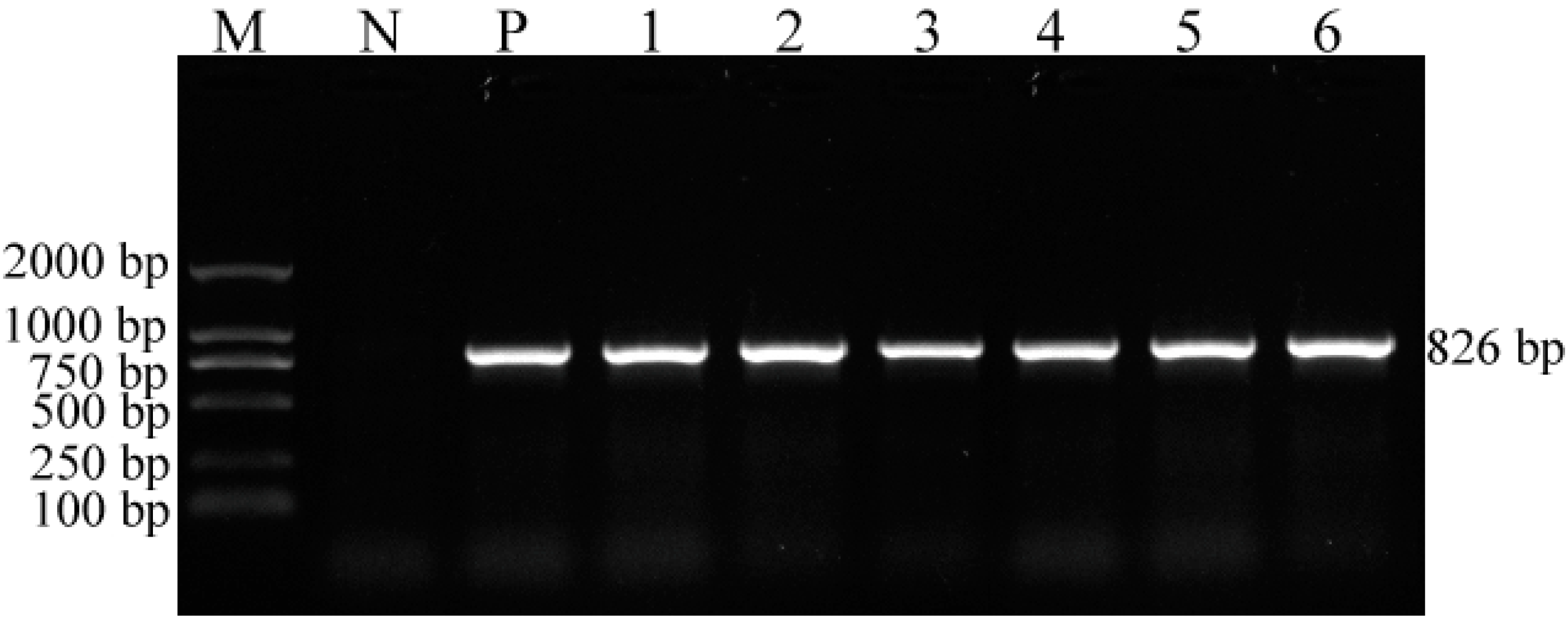
2.1. PCR Amplification of a Unique Fragment of the VP2 Gene and Virus Isolation
2.2. CPV-2 Characterization
2.2.1. TCID50 Test



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | SCDN TICD50/mL | CPV-YA-ZA1 TICD50/mL | CPV-YA-ZA2 TICD50/mL | CPV-YA-ZA4 TICD50/mL | CPV-YA-ZA5 TICD50/mL | CPV-CD-ZA1 TICD50/mL | CPV-LS-ZA1 TICD50/mL |
|---|---|---|---|---|---|---|---|
| Control | 1.6 × 105 | 1.3 × 105 | 1.0 × 105 | 1.6 × 105 | 1.3 × 105 | 2.0 × 105 | 0.8 × 105 |
| 60 °C | 0.9 × 105 | 1.0 × 105 | 0.8 × 105 | 1.4 × 105 | 0.9 × 105 | 1.8 × 105 | 0.4 × 105 |
| pH 3 | 0.6 × 105 | 0.5 × 105 | 0.3 × 105 | 0.8 × 105 | 0.6 × 105 | 1.3 × 105 | 0.1 × 105 |
| Ether | 0.7 × 105 | 0.6 × 105 | 0.4 × 105 | 0.9 × 105 | 0.5 × 105 | 1.2 × 105 | 0.2 × 105 |
| 5-IDUR | 0.4 × 103 | 0.7 × 103 | 0.6 × 103 | 0.9 × 103 | 0.9 × 103 | 1.9 × 103 | 0.9 × 103 |
2.2.2. Viral Hemagglutination (HA)
| Strain Name | Pig | Cow | Sheep | Mouse | Dog | Cat | Chicken |
|---|---|---|---|---|---|---|---|
| SCDN | 29 | <21 | <21 | <21 | <21 | 23 | <21 |
| CPV-YA-ZA1 | 211 | <21 | <21 | <21 | <21 | 24 | <21 |
| CPV-YA-ZA2 | 29 | <21 | <21 | <21 | <21 | 23 | <21 |
| CPV-YA-ZA4 | 29 | <21 | <21 | <21 | <21 | 22 | <21 |
| CPV-YA-ZA5 | 210 | <21 | <21 | <21 | <21 | 23 | <21 |
| CPV-CD-ZA1 | 211 | <21 | <21 | <21 | <21 | 24 | <21 |
| CPV-LS-ZA1 | 27 | <21 | <21 | <21 | <21 | 21 | <21 |
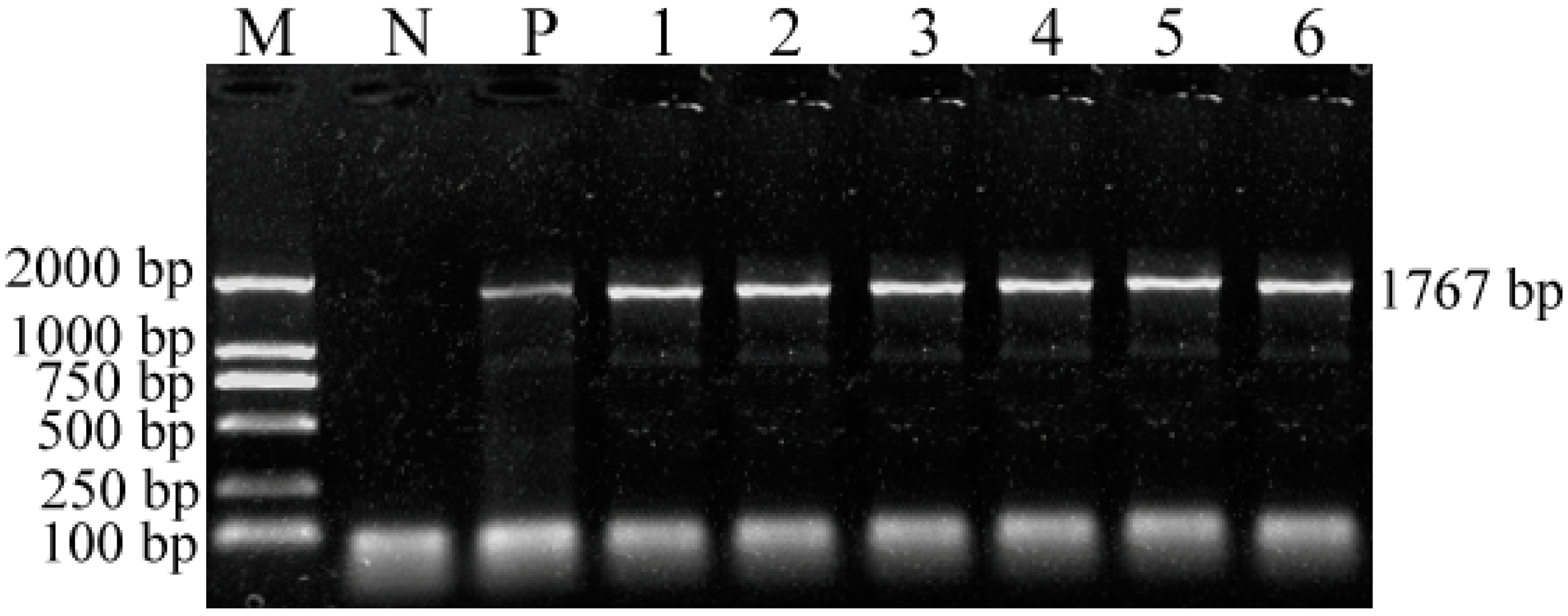
2.3. PCR Amplification of the Complete VP2 Gene and Its Sequence


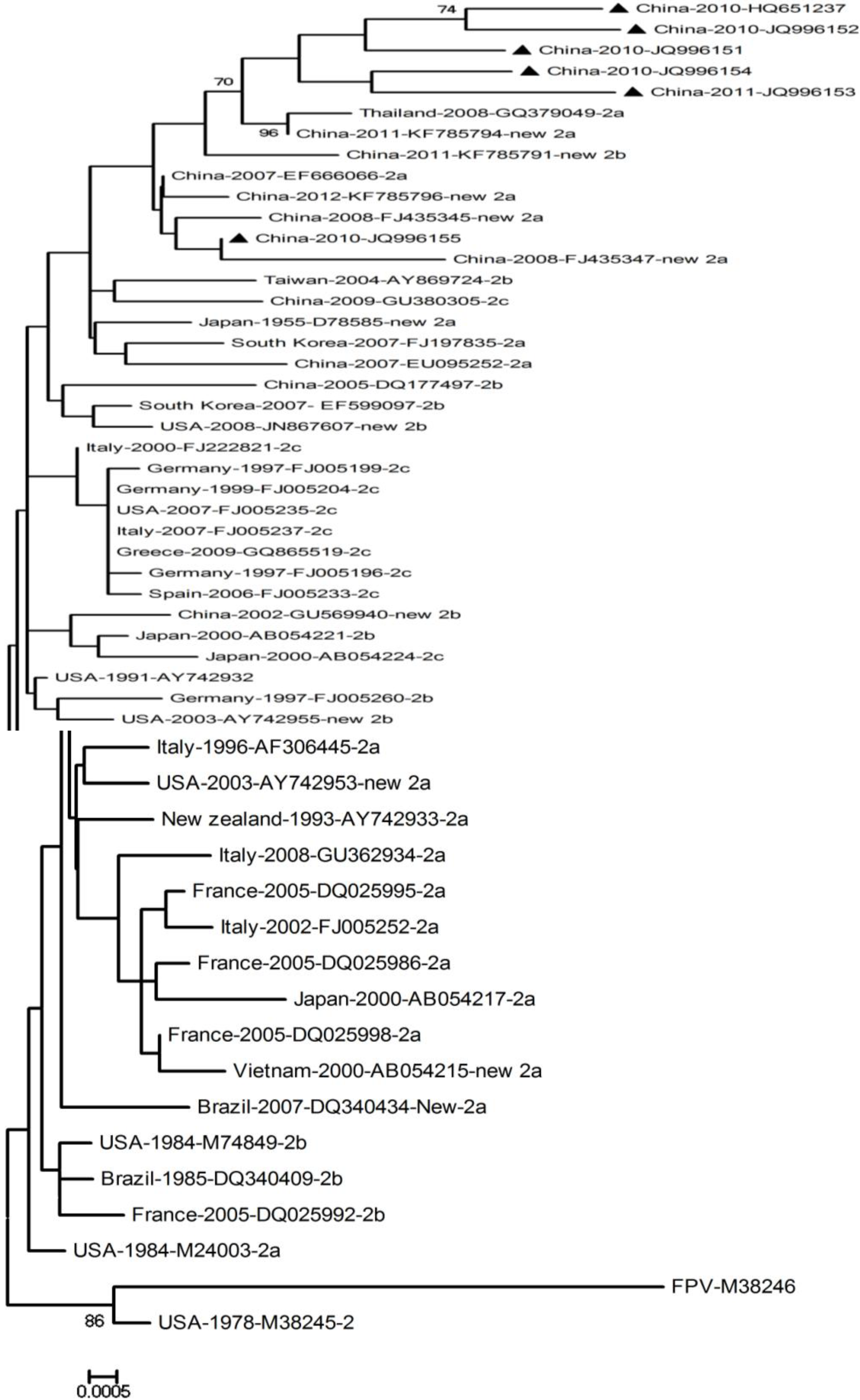
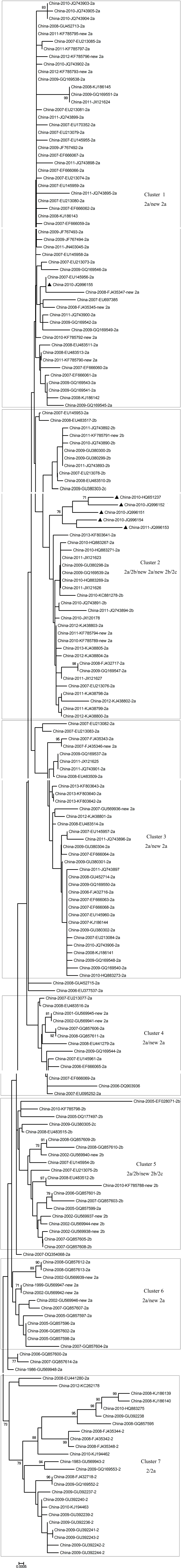
2.4. Phylogenetic Tree and Amino Acid Analysis


| Strain | Amino Acid Position | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 80 | 87 | 93 | 101 | 103 | 297 | 300 | 305 | 323 | 324 | 426 | 555 | 564 | 568 | |
| FPV | Lys | Met | Lys | Ile | Val | Ser | Ala | Asp | Asp | Tyr | Asn | Val | Asn | Ala |
| CPV2 | Arg | Met | Asn | Ile | Ala | Ser | Ala | Asp | Asn | Tyr | Asn | Val | Ser | Gly |
| CPV2a | Arg | Leu | Asn | Thr | Ala | Ser | Gly | Tyr | Asn | Tyr | Asn | Ile | Ser | Gly |
| CPV2b | Arg | Leu | Asn | Thr | Ala | Ser | Gly | Tyr | Asn | Tyr | Asp | Val | Ser | Gly |
| CPV2c | Arg | Leu | Asn | Thr | Ala | Ala | Gly | Tyr | Asn | Tyr | Glu | Val | Ser | Gly |
| new CPV 2a | Arg | Leu | Asn | Thr | Ala | Ala | Gly | Tyr | Asn | Tyr | Asn | Val | Ser | Gly |
| new CPV 2b | Arg | Leu | Asn | Thr | Ala | Ala | Gly | Tyr | Asn | Tyr | Asn | Val | Ser | Gly |
| YAZA1 | Arg | Leu | Asn | Thr | Ala | Ala | Gly | Tyr | Asn | Ile | Asn | Val | Ser | Gly |
| YAZA2 | Arg | Leu | Asn | Thr | Ala | Ala | Gly | Tyr | Asn | Ile | Asn | Val | Ser | Gly |
| YAZA4 | Arg | Leu | Asn | Thr | Ala | Ala | Gly | Tyr | Asn | Asn | Asn | Val | Ser | Gly |
| YAZA5 | Arg | Leu | Asn | Thr | Ala | Ala | Gly | Tyr | Asn | Phe | Asn | Val | Ser | Gly |
| CDZA1 | Arg | Leu | Asn | Thr | Ala | Ala | Gly | Tyr | Asn | Ile | Asn | Val | Ser | Gly |
| LSZA1 | Arg | Leu | Asn | Thr | Ala | Ala | Gly | Tyr | Asn | Ile | Asn | Val | Ser | Gly |
2.5. Homology of the Six Isolates from the Tibetan Mastiff
| Accession Number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 99.2 | 99.0 | 99.4 | 98.7 | 99.3 | 98.5 | 99.0 | 99.6 | 99.0 | 99.0 | 98.8 | 99.0 | 99.0 | 98.8 | 98.9 | |
| 2 | 0.8 | 99.0 | 99.2 | 99.1 | 99.0 | 99.0 | 99.6 | 99.6 | 99.5 | 99.3 | 99.4 | 99.5 | 99.5 | 99.4 | 99.5 | |
| 3 | 1.0 | 1.0 | 99.5 | 99.4 | 98.9 | 98.3 | 98.9 | 99.4 | 98.8 | 98.8 | 98.6 | 98.9 | 98.8 | 98.6 | 98.7 | |
| 4 | 0.6 | 0.8 | 0.5 | 99.0 | 99.1 | 98.5 | 99.0 | 99.5 | 99.0 | 99.0 | 98.8 | 99.0 | 99.0 | 98.8 | 98.9 | |
| 5 | 1.3 | 0.9 | 0.6 | 1.0 | 98.9 | 98.3 | 99.0 | 99.0 | 98.9 | 98.7 | 98.7 | 98.8 | 98.9 | 98.7 | 98.8 | |
| 6 | 0.7 | 1.0 | 1.1 | 0.9 | 1.1 | 98.3 | 99.0 | 99.1 | 98.8 | 98.7 | 98.6 | 98.9 | 98.8 | 98.6 | 98.7 | |
| 7 | 1.6 | 1.0 | 1.7 | 1.6 | 1.7 | 1.7 | 99.3 | 98.9 | 99.3 | 99.1 | 99.1 | 99.1 | 99.3 | 99.1 | 99.2 | |
| 8 | 1.0 | 0.4 | 1.1 | 1.0 | 1.0 | 1.0 | 0.7 | 99.4 | 99.7 | 99.6 | 99.5 | 99.5 | 99.7 | 99.5 | 99.7 | |
| 9 | 0.4 | 0.4 | 0.6 | 0.5 | 1.0 | 0.9 | 1.1 | 0.6 | 99.4 | 99.4 | 99.2 | 99.4 | 99.4 | 99.2 | 99.3 | |
| 10 | 1.0 | 0.5 | 1.2 | 1.0 | 1.1 | 1.2 | 0.7 | 0.3 | 0.6 | 99.5 | 99.6 | 99.6 | 99.8 | 99.6 | 99.7 | |
| 11 | 1.0 | 0.7 | 1.2 | 1.0 | 1.3 | 1.3 | 0.9 | 0.4 | 0.6 | 0.5 | 99.5 | 99.5 | 99.7 | 99.5 | 99.6 | |
| 12 | 1.2 | 0.6 | 1.4 | 1.2 | 1.3 | 1.4 | 0.9 | 0.5 | 0.8 | 0.4 | 0.5 | 99.5 | 99.7 | 99.5 | 99.7 | |
| 13 | 1.0 | 0.5 | 1.1 | 1.0 | 1.2 | 1.1 | 0.9 | 0.5 | 0.6 | 0.4 | 0.5 | 0.5 | 99.7 | 99.5 | 99.7 | |
| 14 | 1.0 | 0.5 | 1.2 | 1.0 | 1.1 | 1.2 | 0.7 | 0.3 | 0.6 | 0.2 | 0.3 | 0.3 | 0.3 | 99.8 | 99.9 | |
| 15 | 1.2 | 0.6 | 1.4 | 1.2 | 1.3 | 1.4 | 0.9 | 0.5 | 0.8 | 0.4 | 0.5 | 0.5 | 0.5 | 0.2 | 99.9 | |
| 16 | 1.1 | 0.5 | 1.3 | 1.1 | 1.2 | 1.3 | 0.8 | 0.3 | 0.7 | 0.3 | 0.4 | 0.3 | 0.3 | 0.1 | 0.1 |
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Preprocessing
| Strain Number | Breed | Age | Gender | Vaccination Status | NCBI Accession Numbers |
|---|---|---|---|---|---|
| CPV-YA-ZA1 | Tibetan mastiff | Two months | Female | Once | HQ651237 |
| CPV-YA-ZA2 | Tibetan mastiff | One month | Female | Once | JQ996151 |
| CPV-YA-ZA3 | Tibetan mastiff | Three months | Female | Twice | / |
| CPV-YA-ZA4 | Tibetan mastiff | Two months | Female | Once | JQ996152 |
| CPV-YA-ZA5 | Tibetan mastiff | Two months | Female | Twice | JQ996153 |
| CPV-LS-ZA1 | Tibetan mastiff | Two months | Male | Once | JQ996155 |
| CPV-CD-ZA1 | Tibetan mastiff | Three months | Female | Twice | JQ996154 |
4.2. DNA Extraction and VP2 Gene Amplification
4.3. Virus Isolation and Characterization by HA and TCID50
4.4. Six Viral Isolates Identified with PCR
4.5. Nucleotide Sequencing and Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Appel, M.J.; Cooper, B.J.; Greisen, H.; Scott, F.; Carmichael, L.E. Canine viral enteritis. I. Status report on corona- and parvo-like viral enteritides. Cornel. Vet. 1979, 69, 123–133. [Google Scholar]
- Appel, M.J.; Scott, F.W.; Carmichael, L.E. Isolation and immunisation studies of a canine parco-like virus from dogs with haemorrhagic enteritis. Vet. Rec. 1979, 105, 156–159. [Google Scholar]
- Decaro, N.; Buonavoglia, C. Canine parvovirus—A review of epidemiological and diagnostic aspects, with emphasis on type 2c. Vet. Microbiol. 2012, 155, 1–12. [Google Scholar] [CrossRef]
- Parrish, C.R.; Have, P.; Foreyt, W.J.; Evermann, J.F.; Senda, M.; Carmichael, L.E. The global spread and replacement of canine parvovirus strains. J. Gen. Virol. 1988, 69, 1111–1116. [Google Scholar] [CrossRef]
- Wang, H.C.; Chen, W.D.; Lin, S.L.; Chan, J.P.; Wong, M.L. Phylogenetic analysis of canine parvovirus VP2 gene in Taiwan. Virus Genes 2005, 31, 171–174. [Google Scholar] [CrossRef]
- Martella, V.; Decaro, N.; Buonavoglia, C. Evolution of CPV-2 and implication for antigenic/genetic characterization. Virus Genes 2006, 33, 11–13. [Google Scholar] [CrossRef]
- Chinchkar, S.R.; Mohana Subramanian, B.; Hanumantha Rao, N.; Rangarajan, P.N.; Thiagarajan, D.; Srinivasan, V.A. Analysis of VP2 gene sequences of canine parvovirus isolates in India. Arch. Virol. 2006, 151, 1881–1887. [Google Scholar] [CrossRef]
- Parrish, C.R.; Aquadro, C.F.; Strassheim, M.L.; Evermann, J.F.; Sgro, J.Y.; Mohammed, H.O. Rapid antigenic-type replacement and DNA sequence evolution of canine parvovirus. J. Virol. 1991, 65, 6544–6552. [Google Scholar]
- Buonavoglia, C.; Martella, V.; Pratelli, A.; Tempesta, M.; Cavalli, A.; Buonavoglia, D.; Bozzo, G.; Elia, G.; Decaro, N.; Carmichael, L. Evidence for evolution of canine parvovirus type 2 in Italy. J. Gen. Virol. 2001, 82, 3021–3025. [Google Scholar]
- Hoelzer, K.; Parrish, C.R. The emergence of parvoviruses of carnivores. Vet. Res. 2010, 41, 39. [Google Scholar] [CrossRef]
- Nandi, S.; Kumar, M. Canine parvovirus: Current perspective. Indian J. Virol. 2010, 21, 31–44. [Google Scholar] [CrossRef]
- Jeoung, S.Y.; Ahn, S.J.; Kim, D. Genetic analysis of VP2 gene of canine parvovirus isolates in Korea. J. Vet. Med. Sci. 2008, 70, 719–722. [Google Scholar] [CrossRef]
- Zhang, R.; Yang, S.; Zhang, W.; Zhang, T.; Xie, Z.; Feng, H.; Wang, S.; Xia, X. Phylogenetic analysis of the VP2 gene of canine parvoviruses circulating in China. Virus Genes 2010, 40, 397–402. [Google Scholar] [CrossRef]
- Zhao, Y.; Lin, Y.; Zeng, X.; Lu, C.; Hou, J. Genotyping and pathobiologic characterization of canine parvovirus circulating in Nanjing, China. Virol. J. 2013, 10, 272. [Google Scholar] [CrossRef]
- Zhang, Q.; Xu, X.M.; Zhai, G.Q.; Wang, Z.; Hou, S.H. Molecular characterisation of canine parvovirus strains circulating in China. Afr. J. Biotechnol. 2010, 9, 4556–4560. [Google Scholar]
- Zhang, R.Z.; Yang, S.T.; Feng, H.; Cui, C.S.; Xia, X.Z. The first detection of canine parvovirus type 2c in China. J. Pathog. Biol. 2010, 5, 246–249. [Google Scholar]
- Xu, J.; Guo, H.C.; Wei, Y.Q.; Shu, L.; Wang, J.; Li, J.S.; Cao, S.Z.; Sun, S.Q. Phylogenetic analysis of canine parvovirus isolates from Sichuan and Gansu provinces of China in 2011. Transbound. Emerg. Dis. 2014. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Z.; Li, Y.; Zhao, X.; Dong, L.; Pan, Z.; Sun, Y.; Li, N.; Xu, Y.; Xie, Z. Origin and phylogenetic analysis of Tibetan mastiff based on the mitochondrial DNA sequence. J. Genet. Genomics 2008, 35, 335–340. [Google Scholar] [CrossRef]
- Li, W.; Li, T.; Liu, Y.; Gao, Y.; Yang, S.; Feng, N.; Sun, H.; Wang, S.; Wang, L.; Bu, Z.; et al. Genetic characterization of an isolate of canine distemper virus from a Tibetan mastiff in China. Virus Genes 2014. [Google Scholar] [CrossRef]
- Teng, Q.; Zhang, X.; Xu, D.; Zhou, J.; Dai, X.; Chen, Z.; Li, Z. Characterization of an H3N2 canine influenza virus isolated from Tibetan mastiffs in China. Vet. Microbiol. 2013, 162, 345–352. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Y.H.; Ye, Q.; Meng, P.; Yin, H.; Zhang, D.L. Serological survey of toxoplasma gondii in Tibetan mastiffs (Canis lupus familiaris) and yaks (Bos grunniens) in Qinghai, China. Parasites Vectors 2012, 5, 35. [Google Scholar] [CrossRef]
- Parthiban, S.; Mukhopadhyay, H.K.; Panneer, D.; Antony, P.X.; Pillai, R.M. Isolation and typing of canine parvovirus in CRFK cell line in Puducherry, South India. Indian J. Microbiol. 2011, 51, 456–460. [Google Scholar] [CrossRef]
- Carmichael, L.E.; Joubert, J.C.; Pollock, R.V. Hemagglutination by canine parvovirus: Serologic studies and diagnostic applications. Am. J. Vet. Res. 1980, 41, 784–791. [Google Scholar]
- Goddard, A.; Leisewitz, A.L. Canine parvovirus. Vet. Clin. N. Am. Small Anim. Pract. 2010, 40, 1041–1053. [Google Scholar] [CrossRef]
- Pereira, C.A.; Leal, E.S.; Durigon, E.L. Selective regimen shift and demographic growth increase associated with the emergence of high-fitness variants of canine parvovirus. Infect. Genet. Evol. 2007, 7, 399–409. [Google Scholar] [CrossRef]
- Ohshima, T.; Hisaka, M.; Kawakami, K.; Kishi, M.; Tohya, Y.; Mochizuki, M. Chronological analysis of canine parvovirus type 2 isolates in Japan. J. Vet. Med. Sci. 2008, 70, 769–775. [Google Scholar] [CrossRef]
- Yoon, S.H.; Jeong, W.; Kim, H.J.; An, D.J. Molecular insights into the phylogeny of canine parvovirus 2 (CPV-2) with emphasis on Korean isolates: A bayesian approach. Arch. Virol. 2009, 154, 1353–1360. [Google Scholar] [CrossRef]
- Phromnoi, S.; Sirinarumitr, K.; Sirinarumitr, T. Sequence analysis of VP2 gene of canine parvovirus isolates in Thailand. Virus Genes 2010, 41, 23–29. [Google Scholar] [CrossRef]
- Soma, T.; Taharaguchi, S.; Ohinata, T.; Ishii, H.; Hara, M. Analysis of the VP2 protein gene of canine parvovirus strains from affected dogs in Japan. Res. Vet. Sci. 2013, 94, 368–371. [Google Scholar] [CrossRef]
- Mukhopadhyay, H.K.; Matta, S.L.; Amsaveni, S.; Antony, P.X.; Thanislass, J.; Pillai, R.M. Phylogenetic analysis of canine parvovirus partial VP2 gene in India. Virus Genes 2014, 48, 89–95. [Google Scholar] [CrossRef]
- Mittal, M.; Chakravarti, S.; Mohapatra, J.K.; Chug, P.K.; Dubey, R.; Upmanuyu, V.; Narwal, P.S.; Kumar, A.; Churamani, C.P.; Kanwar, N.S. Molecular typing of canine parvovirus strains circulating from 2008 to 2012 in an organized kennel in India reveals the possibility of vaccination failure. Infect. Genet. Evol. 2014, 23, 1–6. [Google Scholar] [CrossRef]
- Lin, C.N.; Chien, C.H.; Chiou, M.T.; Chueh, L.L.; Hung, M.Y.; Hsu, H.S. Genetic characterization of type 2a canine parvoviruses from Taiwan reveals the emergence of an Ile324 mutation in VP2. Virol. J. 2014, 11, 39. [Google Scholar] [CrossRef]
- Perez, R.; Bianchi, P.; Calleros, L.; Francia, L.; Hernandez, M.; Maya, L.; Panzera, Y.; Sosa, K.; Zoller, S. Recent spreading of a divergent canine parvovirus type 2a (CPV-2a) strain in a CPV-2c homogenous population. Vet. Microbiol. 2012, 155, 214–219. [Google Scholar] [CrossRef]
- Csagola, A.; Varga, S.; Lorincz, M.; Tuboly, T. Analysis of the full-length VP2 protein of canine parvoviruses circulating in Hungary. Arch. Virol. 2014. [Google Scholar] [CrossRef]
- Hueffer, K.; Parrish, C.R. Parvovirus host range, cell tropism and evolution. Curr. Opin. Microbiol. 2003, 6, 392–398. [Google Scholar] [CrossRef]
- Qin, Q.; Loeffler, I.K.; Li, M.; Tian, K.; Wei, F. Sequence analysis of a canine parvovirus isolated from a red panda (Ailurus fulgens) in China. Virus Genes 2007, 34, 299–302. [Google Scholar] [CrossRef]
- Chen, X.Y.; Xie, Z.J.; Zhao, Z.P.; Jiang, S.J.; Zhao, H.K.; Zhu, Y.L.; Zhang, X.X. Genetic diversity of parvovirus isolates from dogs and wild animals in China. J. Wildl. Dis. 2011, 47, 1036–1039. [Google Scholar] [CrossRef]
- Nandi, S.; Chidri, S.; Kumar, M.; Chauhan, R.S. Occurrence of canine parvovirus type 2c in the dogs with haemorrhagic enteritis in India. Res. Vet. Sci. 2010, 88, 169–171. [Google Scholar] [CrossRef]
- Ge, R.; Peng, G.; Xia, X.; Yang, S.; Su, J.; Zou, X.; Feng, N.; Ma, X.; Shi, J. Isolation and identification of canine parvovirus Sichuan strains. Chin. J. Prev. Vet. Med. 2007, 29, 836–839. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhong, Z.; Liang, L.; Zhao, J.; Xu, X.; Cao, X.; Liu, X.; Zhou, Z.; Ren, Z.; Shen, L.; Geng, Y.; et al. First Isolation of New Canine Parvovirus 2a from Tibetan Mastiff and Global Analysis of the Full-Length VP2 Gene of Canine Parvoviruses 2 in China. Int. J. Mol. Sci. 2014, 15, 12166-12187. https://doi.org/10.3390/ijms150712166
Zhong Z, Liang L, Zhao J, Xu X, Cao X, Liu X, Zhou Z, Ren Z, Shen L, Geng Y, et al. First Isolation of New Canine Parvovirus 2a from Tibetan Mastiff and Global Analysis of the Full-Length VP2 Gene of Canine Parvoviruses 2 in China. International Journal of Molecular Sciences. 2014; 15(7):12166-12187. https://doi.org/10.3390/ijms150712166
Chicago/Turabian StyleZhong, Zhijun, Luqi Liang, Juan Zhao, Xiaoyang Xu, Xuefeng Cao, Xuehan Liu, Ziyao Zhou, Zhihua Ren, Liuhong Shen, Yi Geng, and et al. 2014. "First Isolation of New Canine Parvovirus 2a from Tibetan Mastiff and Global Analysis of the Full-Length VP2 Gene of Canine Parvoviruses 2 in China" International Journal of Molecular Sciences 15, no. 7: 12166-12187. https://doi.org/10.3390/ijms150712166
APA StyleZhong, Z., Liang, L., Zhao, J., Xu, X., Cao, X., Liu, X., Zhou, Z., Ren, Z., Shen, L., Geng, Y., Gu, X., & Peng, G. (2014). First Isolation of New Canine Parvovirus 2a from Tibetan Mastiff and Global Analysis of the Full-Length VP2 Gene of Canine Parvoviruses 2 in China. International Journal of Molecular Sciences, 15(7), 12166-12187. https://doi.org/10.3390/ijms150712166