Ficus carica Polysaccharides Promote the Maturation and Function of Dendritic Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
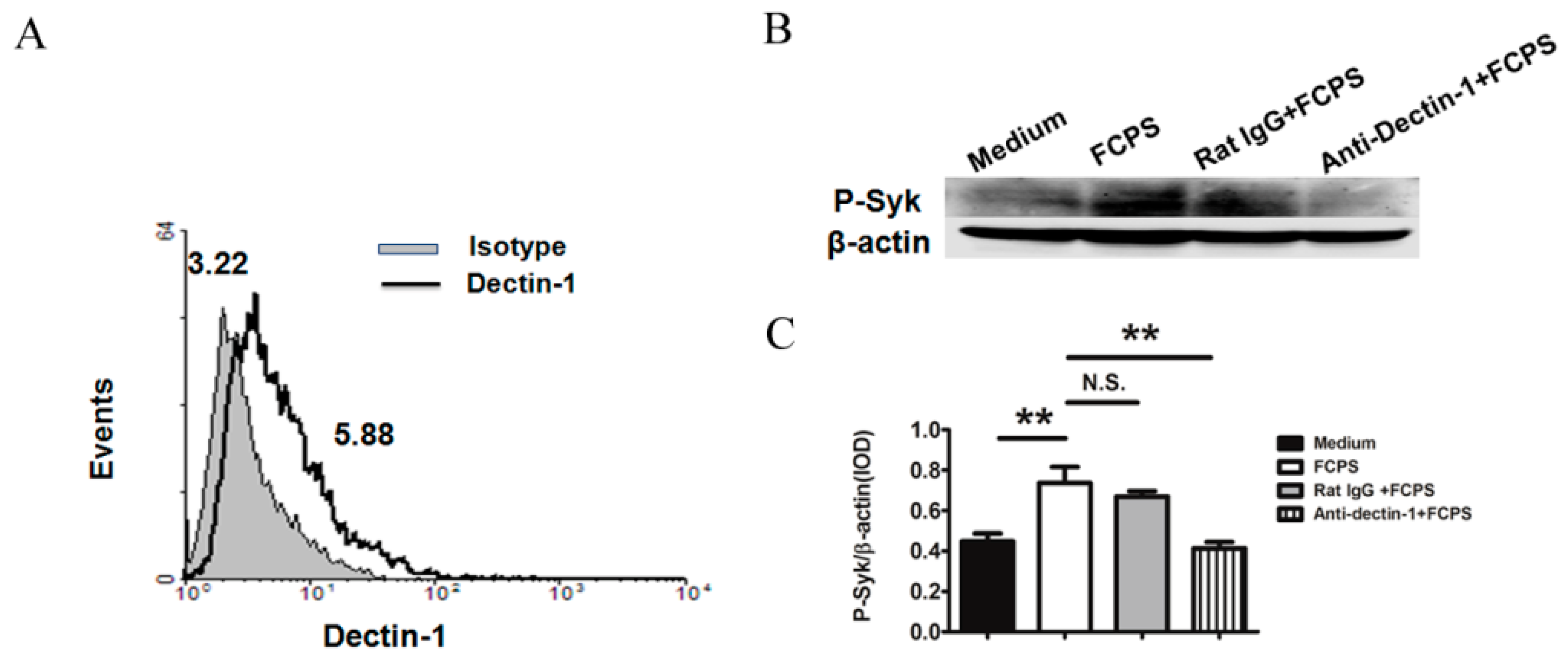
2.1. FCPS Activates Syk through the Dectin-1 Pathway in Bone Marrow Dendritic Cells (BMDCs)

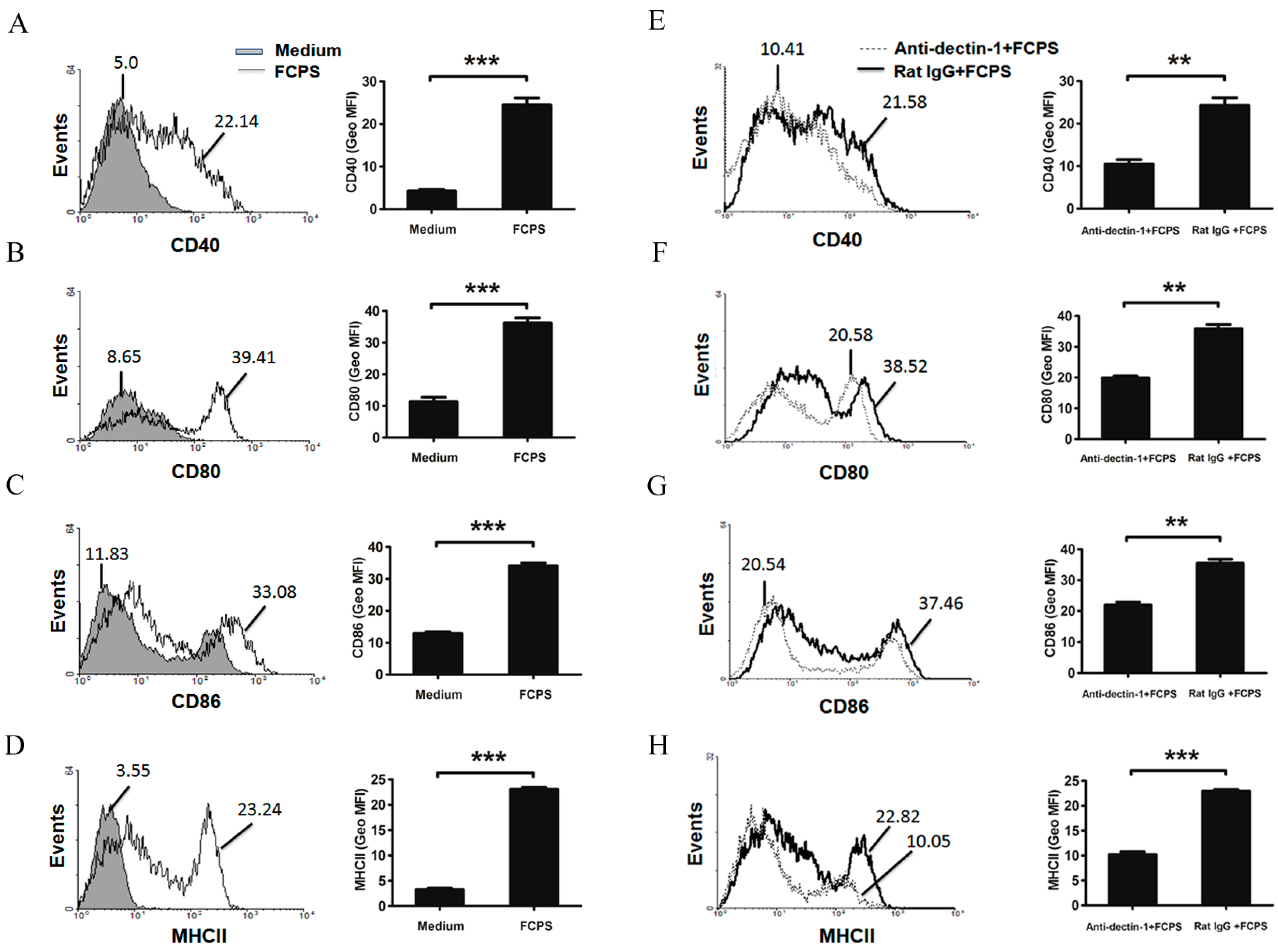
2.2. FCPS Promote the Activation and Maturation of BMDCs
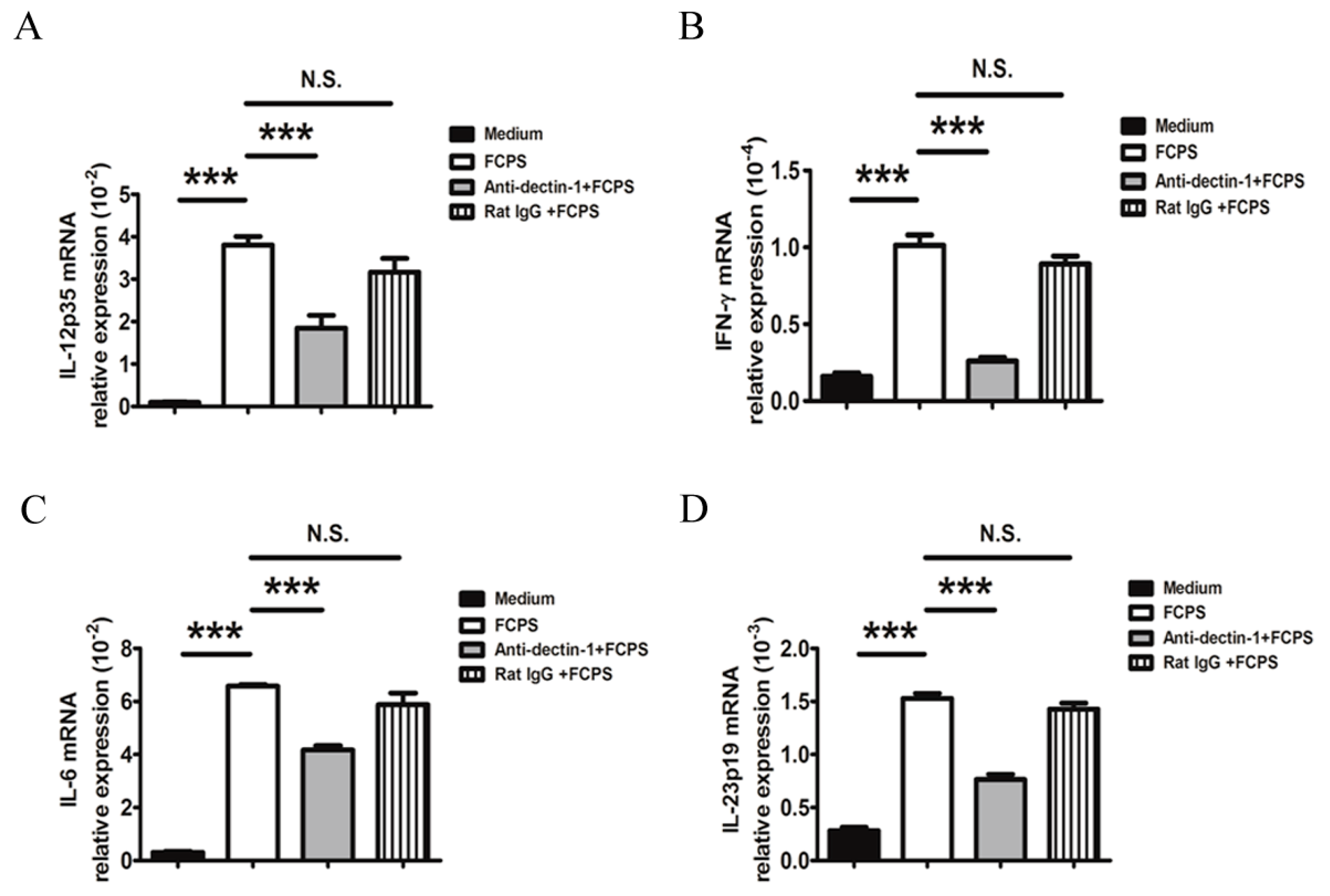
2.3. FCPS Enhance the Expression of Multiple Cytokines Secreted by DCs


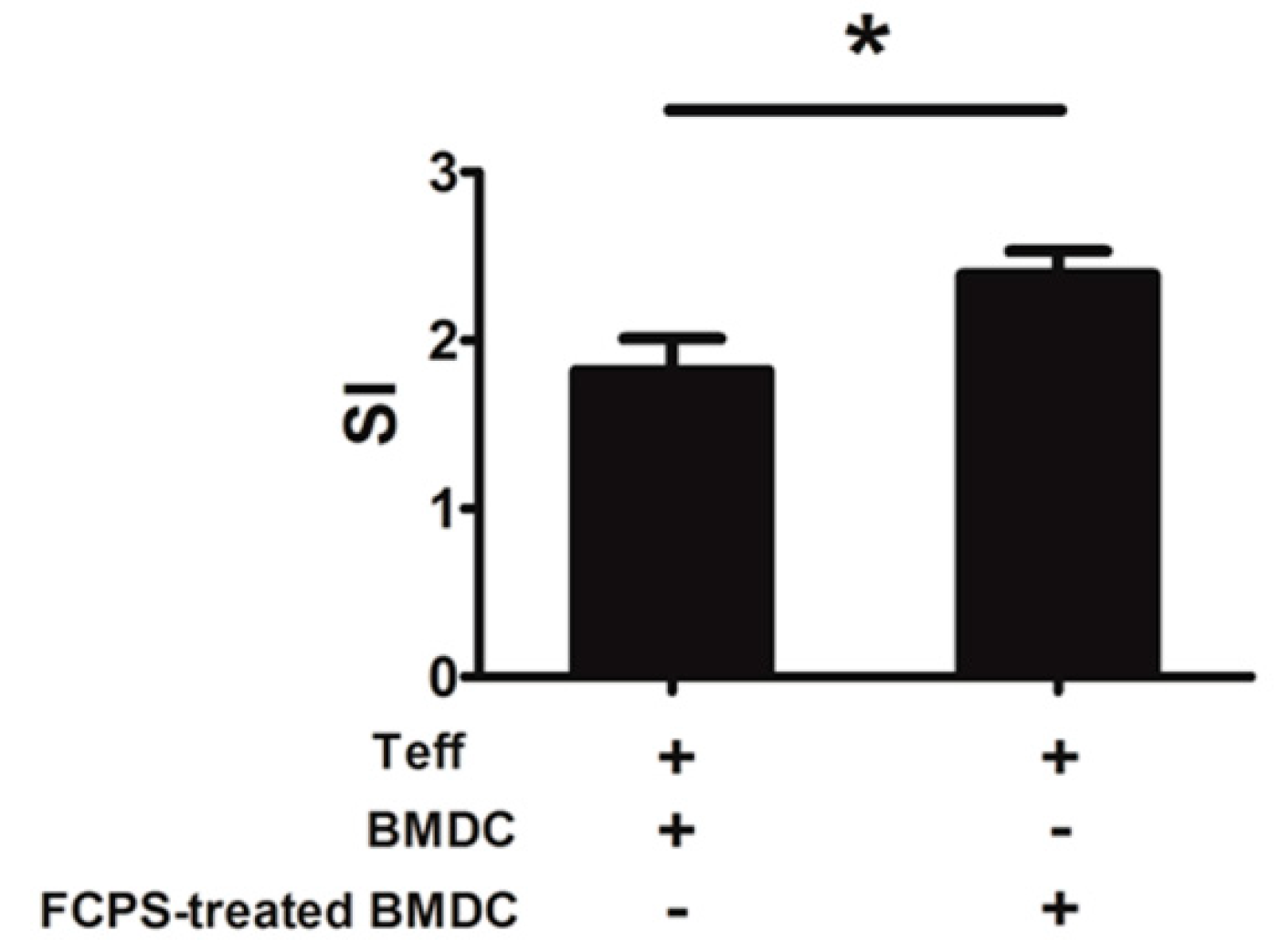
2.4. FCPS Augments the Immunostimulatory Capacity of DCs
3. Discussion
4. Experimental Section
4.1. Mice
4.2. FCPS
4.3. Cell Line
4.4. Bone Marrow-Derived DC (BMDC)
4.5. Western Blot Analysis
4.6. RNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
4.7. Flow Cytometry
4.8. In Vitro Proliferation Assays
4.9. Statistics
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Conflicts of Interest
References
- Park, E.J.; Pezzuto, J.M. Botanicals in cancer chemoprevention. Cancer Metastasis Rev. 2002, 21, 231–255. [Google Scholar] [CrossRef]
- Lau, K.M.; He, Z.D.; Dong, H.; Fung, K.P.; But, P.P. Anti-oxidative, anti-inflammatory and hepato-protective effects of Ligustrum robustum. J. Ethnopharmacol. 2002, 83, 63–71. [Google Scholar] [CrossRef]
- Lin, C.L.; Wang, C.C.; Chang, S.C.; Inbaraj, B.S.; Chen, B.H. Antioxidative activity of polysaccharide fractions isolated from Lycium barbarum Linnaeus. Int. J. Biol. Macromol. 2009, 45, 146–151. [Google Scholar] [CrossRef]
- Li, W.J.; Chen, Y.; Nie, S.P.; Xie, M.Y.; He, M.; Zhang, S.S.; Zhu, K.X. Ganoderma atrum polysaccharide induces anti-tumor activity via the mitochondrial apoptotic pathway related to activation of host immune response. J. Cell. Biochem. 2011, 112, 860–871. [Google Scholar] [CrossRef]
- Masuda, Y.; Ito, K.; Konishi, M.; Nanba, H. A polysaccharide extracted from Grifola frondosa enhances the anti-tumor activity of bone marrow-derived dendritic cell-based immunotherapy against murine colon cancer. Cancer Immunol. Immunother. 2010, 59, 1531–1541. [Google Scholar] [CrossRef]
- Liu, M.; Wu, K.; Mao, X.; Wu, Y.; Ouyang, J. Astragalus polysaccharide improves insulin sensitivity in KKAy mice: Regulation of PKB/GLUT4 signaling in skeletal muscle. J. Ethnopharmacol. 2010, 127, 32–37. [Google Scholar] [CrossRef]
- Kiho, T.; Sobue, S.; Ukai, S. Structural features and hypoglycemic activities of two polysaccharides from a hot-water extract of Agrocybe cylindracea. Carbohydr. Res. 1994, 251, 81–87. [Google Scholar]
- Kobayashi, M.; Magishi, N.; Matsushita, H.; Hashimoto, T.; Fujimoto, M.; Suzuki, M.; Tsuji, K.; Saito, M.; Inoue, E.; Yoshikawa, Y.; et al. Hypolipidemic effect of Shoyu polysaccharides from soy sauce in animals and humans. Int. J. Mol. Med. 2008, 22, 565–570. [Google Scholar]
- Liu, L.; Yuan, S.; Long, Y.; Guo, Z.; Sun, Y.; Li, Y.; Niu, Y.; Li, C.; Mei, Q. Immunomodulation of Rheum tanguticum polysaccharide (RTP) on the immunosuppressive effects of dexamethasone (DEX) on the treatment of colitis in rats induced by 2,4,6-trinitrobenzene sulfonic acid. Int. Immunopharmacol. 2009, 9, 1568–1577. [Google Scholar] [CrossRef]
- Tzianabos, A.O. Polysaccharide immunomodulators as therapeutic agents: Structural aspects and biologic function. Clin. Microbiol. Rev. 2000, 13, 523–533. [Google Scholar] [CrossRef]
- Lansky, E.P.; Paavilainen, H.M.; Pawlus, A.D.; Newman, R.A. Ficus spp. (fig): Ethnobotany and potential as anticancer and anti-inflammatory agents. J. Ethnopharmacol. 2008, 119, 195–213. [Google Scholar] [CrossRef]
- Solomon, A.; Golubowicz, S.; Yablowicz, Z.; Grossman, S.; Bergman, M.; Gottlieb, H.E.; Altman, A.; Kerem, Z.; Flaishman, M.A. Antioxidant activities and anthocyanin content of fresh fruits of common Fig (Ficus carica L.). J. Agric. Food Chem. 2006, 54, 7717–7723. [Google Scholar] [CrossRef]
- Yang, X.-M.; Yu, W.; Ou, Z.-P.; Ma, H.-L.; Liu, W.-M.; Ji, X.-L. Antioxidant and Immunity Activity of Water Extract and Crude Polysaccharide from Ficus carica L. Fruit. Plant Foods Hum. Nutr. 2009, 64, 167–173. [Google Scholar] [CrossRef]
- Joffre, O.; Nolte, M.A.; Sporri, R.; e Sousa, C.R. Inflammatory signals in dendritic cell activation and the induction of adaptive immunity. Immunol. Rev. 2009, 227, 234–247. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef]
- Chabner, B.A.; Roberts, T.G. Chemotherapy and the war on cancer. Nat. Rev. Cancer 2005, 5, 65–72. [Google Scholar] [CrossRef]
- Wasser, S. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar]
- Jeong, S.C.; Koyyalamudi, S.R.; Jeong, Y.T.; Song, C.H.; Pang, G. Macrophage immunomodulating and antitumor activities of polysaccharides isolated from Agaricus bisporus white button mushrooms. J. Med. Food 2012, 15, 58–65. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, J.Y.; Kang, J.S.; Kim, H.M.; Kim, Y.O.; Hong, I.P.; Lee, M.K.; Hong, J.T.; Kim, Y.; Han, S.-B. Cordlan polysaccharide isolated from mushroom Cordyceps militaris induces dendritic cell maturation through toll-like receptor 4 signalings. Food Chem. Toxicol. 2010, 48, 1926–1933. [Google Scholar] [CrossRef]
- Ooi, V.E.; Liu, F. Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Curr. Med. Chem. 2000, 7, 715–729. [Google Scholar] [CrossRef]
- Kataoka, K.; Muta, T.; Yamazaki, S.; Takeshige, K. Activation of Macrophages by Linear (1→3)-β-d-Glucans: Implications for the recognition of fungi by innate immunity. J. Biol. Chem. 2002, 277, 36825–36831. [Google Scholar] [CrossRef]
- Ohno, N.; Furukawa, M.; Miura, N.N.; Adachi, Y.; Motoi, M.; Yadomae, T. Antitumor β glucan from the cultured fruit body of Agaricus blazei. Biol. Pharm. Bull. 2001, 24, 820–828. [Google Scholar]
- Brown, G.D.; Gordon, S. Fungal β-glucans and mammalian immunity. Immunity 2003, 19, 311–315. [Google Scholar] [CrossRef]
- Han, S.B.; Yoon, Y.D.; Ahn, H.J.; Lee, H.S.; Lee, C.W.; Yoon, W.K.; Park, S.K.; Kim, H.M. Toll-like receptor-mediated activation of B cells and macrophages by polysaccharide isolated from cell culture of Acanthopanax senticosus. Int. Immunopharmacol. 2003, 3, 1301–1312. [Google Scholar] [CrossRef]
- Ahn, K.-S.; Sim, W.; Kim, H.; Han, S.; Kim, I.-H. Immunostimulating polysaccharide from cell culture of Angelica gigas Nakai. Biotechnol. Lett. 1998, 20, 5–7. [Google Scholar]
- Cui, R.; He, J.; Wang, B.; Zhang, F.; Chen, G.; Yin, S.; Shen, H. Suppressive effect of Astragalus membranaceus Bunge on chemical hepatocarcinogenesis in rats. Cancer Chemother. Pharmacol. 2003, 51, 75–80. [Google Scholar] [CrossRef]
- Brown, G.D. Dectin-1: A signalling non-TLR pattern-recognition receptor. Nat. Rev. Immunol. 2006, 6, 33–43. [Google Scholar] [CrossRef]
- Gringhuis, S.I.; den Dunnen, J.; Litjens, M.; van der Vlist, M.; Wevers, B.; Bruijns, S.C.M.; Geijtenbeek, T.B.H. Dectin-1 directs T helper cell differentiation by controlling noncanonical NF-κB activation through Raf-1 and Syk. Nat. Immunol. 2009, 10, 203–213. [Google Scholar] [CrossRef]
- Reid, D.M.; Gow, N.A.R.; Brown, G.D. Pattern recognition: Recent insights from Dectin-1. Curr. Opin. Immunol. 2009, 21, 30–37. [Google Scholar] [CrossRef]
- Tian, J.; Ma, J.; Ma, K.; Ma, B.; Tang, X.; Baidoo, S.E.; Tong, J.; Yan, J.; Lu, L.; Xu, H.; et al. Up-regulation of GITRL on dendritic cells by WGP improves anti-tumor immunity in murine lewis lung carcinoma. PLoS One 2012, 7, e46936. [Google Scholar]
- Tian, J.; Ma, J.; Ma, K.; Guo, H.; Baidoo, S.E.; Zhang, Y.; Yan, J.; Lu, L.; Xu, H.; Wang, S. β-Glucan enhances antitumor immune responses by regulating differentiation and function of monocytic myeloid-derived suppressor cells. Eur. J. Immunol. 2013, 43, 1220–1230. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tian, J.; Zhang, Y.; Yang, X.; Rui, K.; Tang, X.; Ma, J.; Chen, J.; Xu, H.; Lu, L.; Wang, S. Ficus carica Polysaccharides Promote the Maturation and Function of Dendritic Cells. Int. J. Mol. Sci. 2014, 15, 12469-12479. https://doi.org/10.3390/ijms150712469
Tian J, Zhang Y, Yang X, Rui K, Tang X, Ma J, Chen J, Xu H, Lu L, Wang S. Ficus carica Polysaccharides Promote the Maturation and Function of Dendritic Cells. International Journal of Molecular Sciences. 2014; 15(7):12469-12479. https://doi.org/10.3390/ijms150712469
Chicago/Turabian StyleTian, Jie, Yue Zhang, Xiaomin Yang, Ke Rui, Xinyi Tang, Jie Ma, Jianguo Chen, Huaxi Xu, Liwei Lu, and Shengjun Wang. 2014. "Ficus carica Polysaccharides Promote the Maturation and Function of Dendritic Cells" International Journal of Molecular Sciences 15, no. 7: 12469-12479. https://doi.org/10.3390/ijms150712469
APA StyleTian, J., Zhang, Y., Yang, X., Rui, K., Tang, X., Ma, J., Chen, J., Xu, H., Lu, L., & Wang, S. (2014). Ficus carica Polysaccharides Promote the Maturation and Function of Dendritic Cells. International Journal of Molecular Sciences, 15(7), 12469-12479. https://doi.org/10.3390/ijms150712469