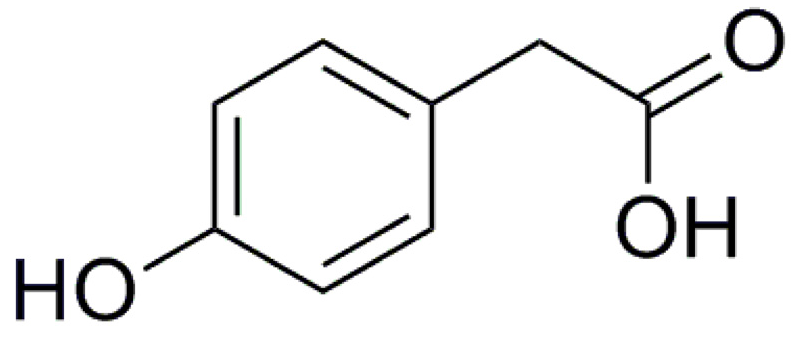
4-Hydroxyphenylacetic Acid Attenuated Inflammation and Edema via Suppressing HIF-1α in Seawater Aspiration-Induced Lung Injury in Rats

Abstract
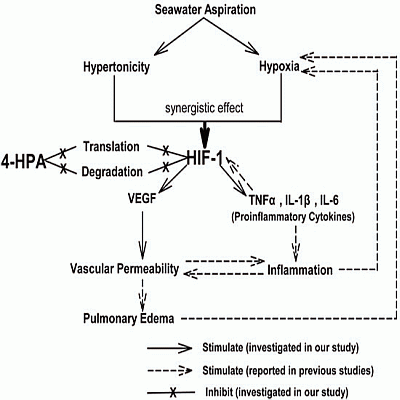
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
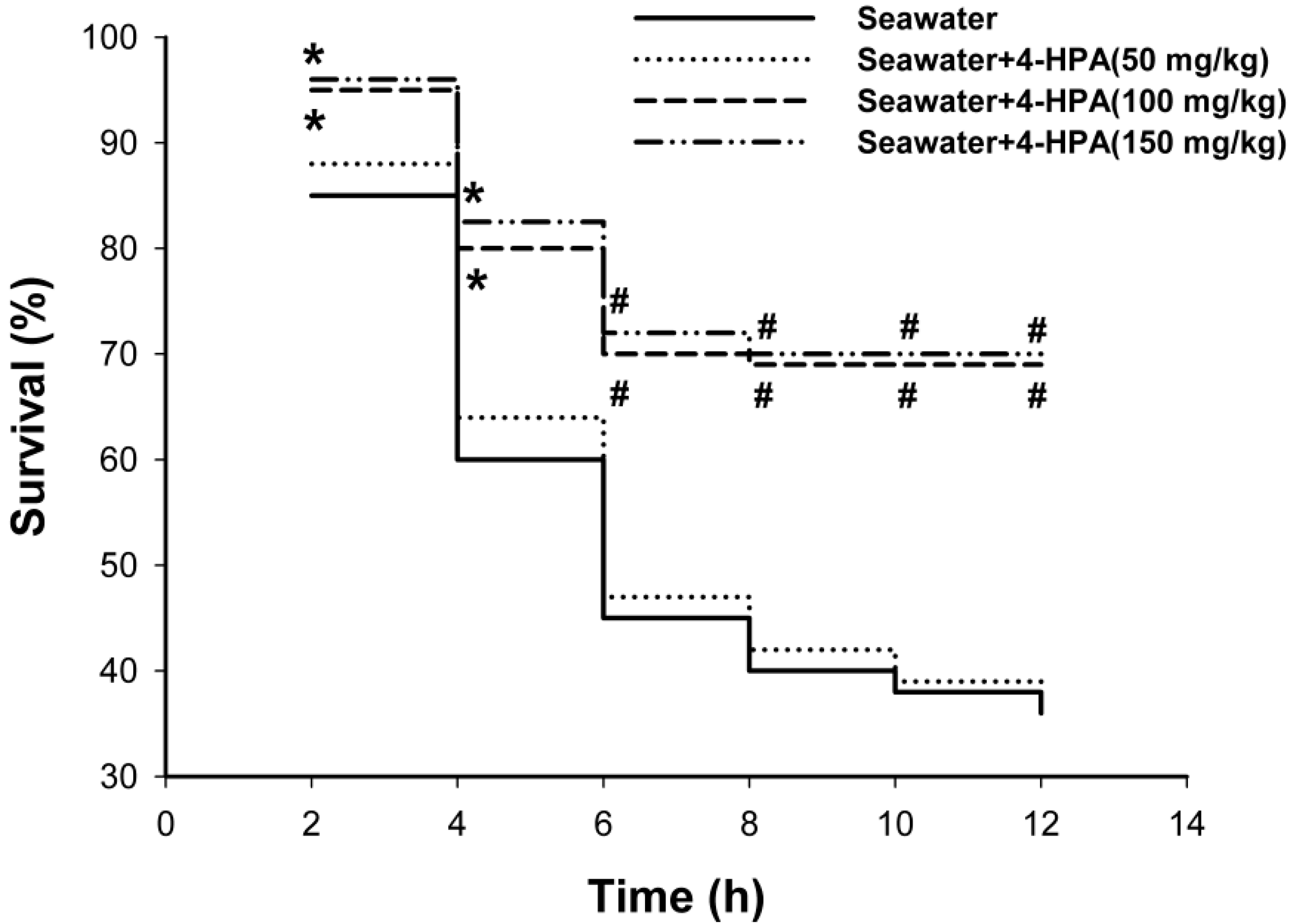
2.1. 4-Hydroxyphenylacetic Acid (4-HPA) Reduced Seawater Instillation-Induced Mortality in Rats

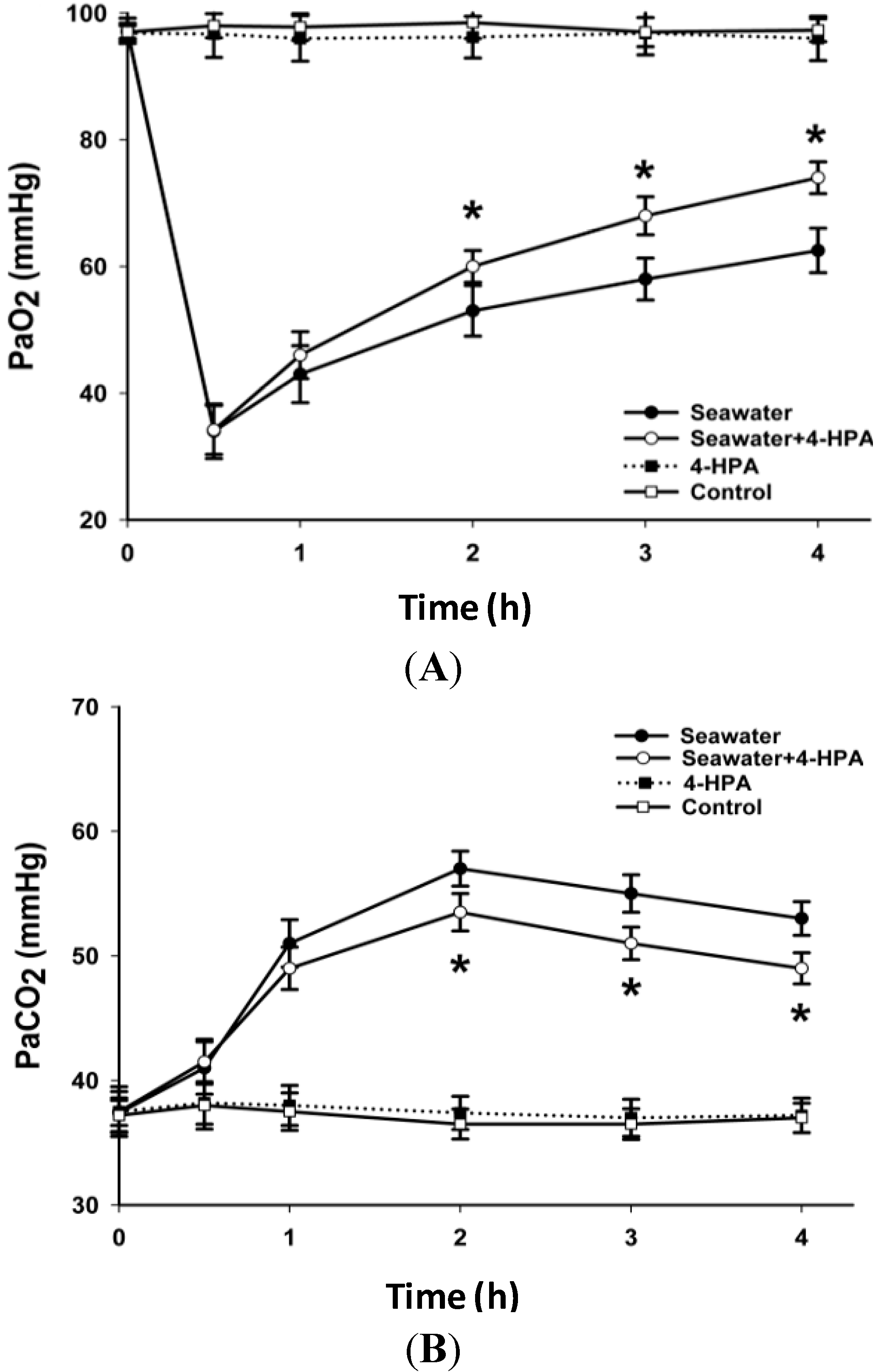
2.2. 4-HPA Increased PaO2 and Decreased PaCO2 in Seawater Instillation Rats

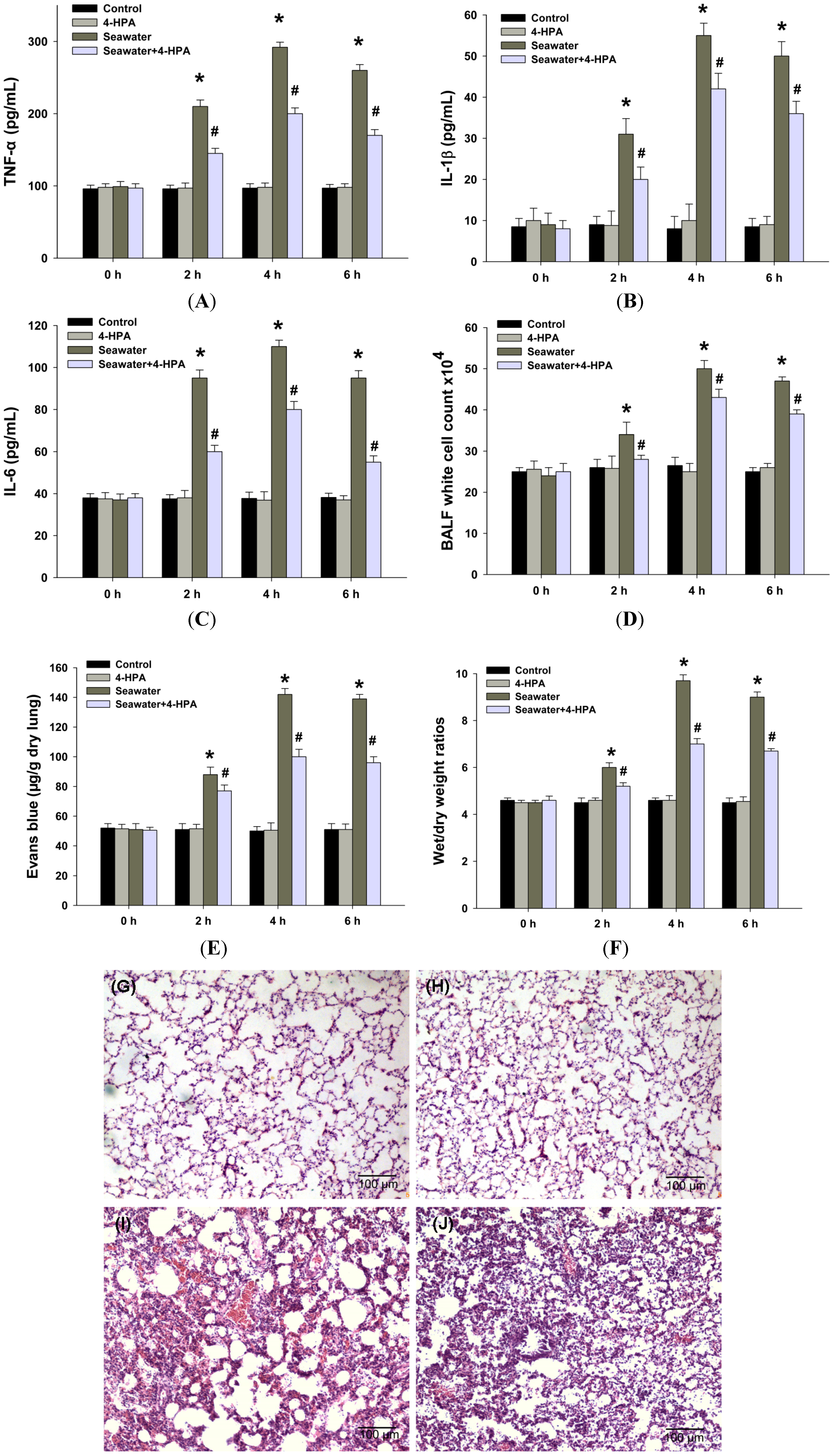
2.3. 4-HPA Attenuated Inflammation, Vascular Leak, and Edema in Seawater Instillation-Induced Lung Injury in Rats

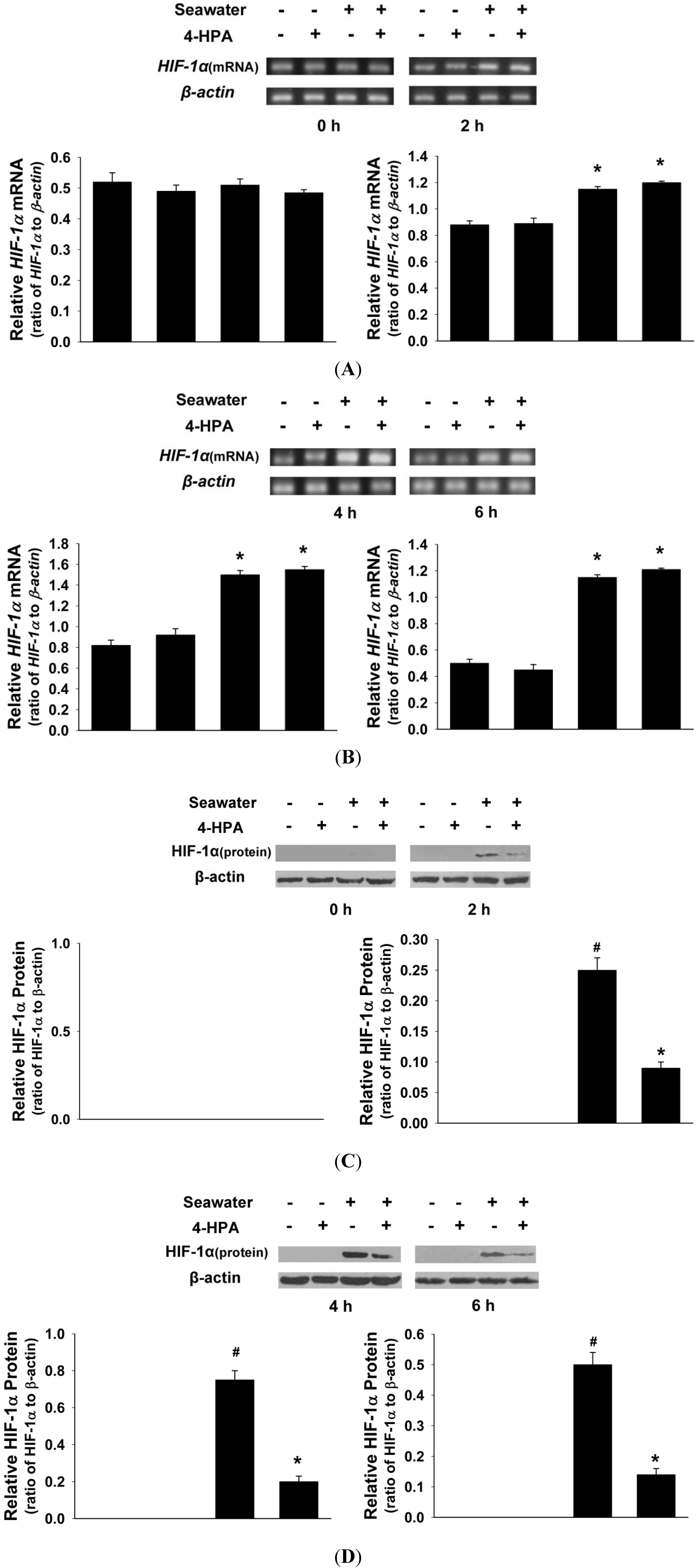
2.4. 4-HPA Decreased Seawater Instillation-Induced HIF-1α Protein Level, but not mRNA Level, in Lung Tissues in Rats

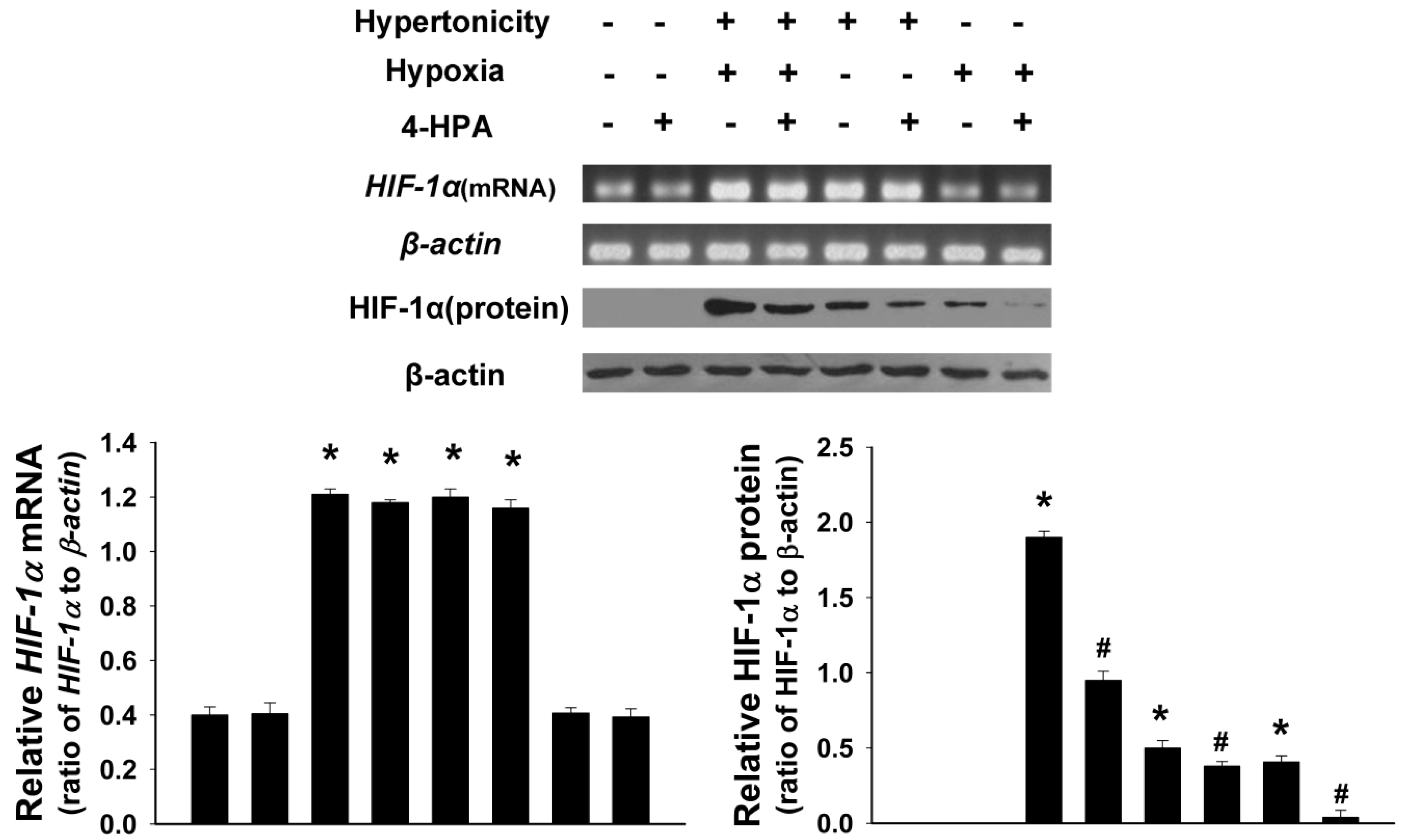
2.5. 4-HPA Decreased Hypertonicity and Hypoxia-Induced HIF-1α Protein Level, but not mRNA Level, in Primary Rat Alveolar Epithelial Cells (AEC)

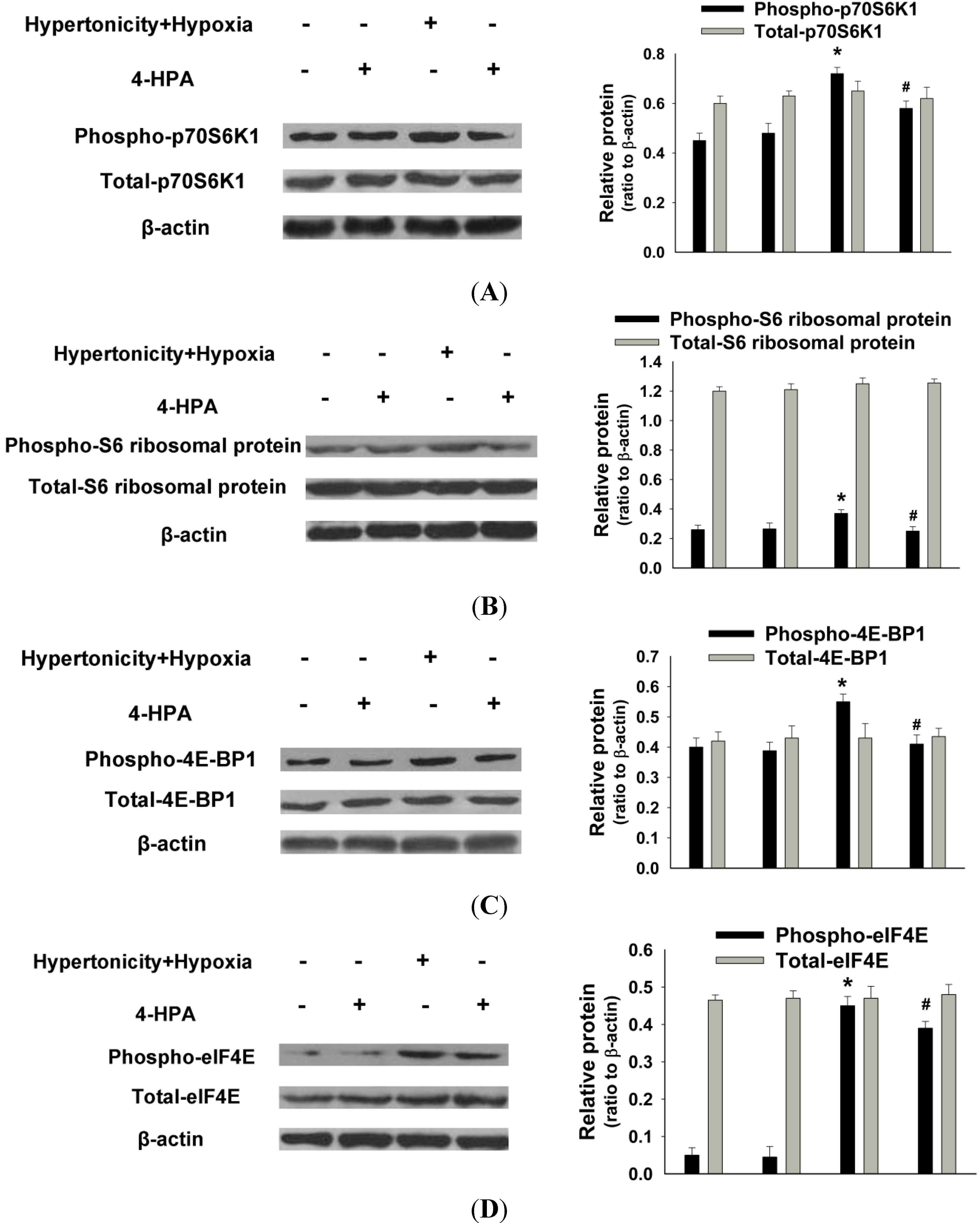
2.6. 4-HPA Decreased Hypertonicity and Hypoxia-Induced HIF-1α Protein Level through Inhibiting the Activations of Protein Translational Regulators, Including p70S6K1, S6 Ribosomal Protein, 4E-BP1, and eIF4E in Primary AEC

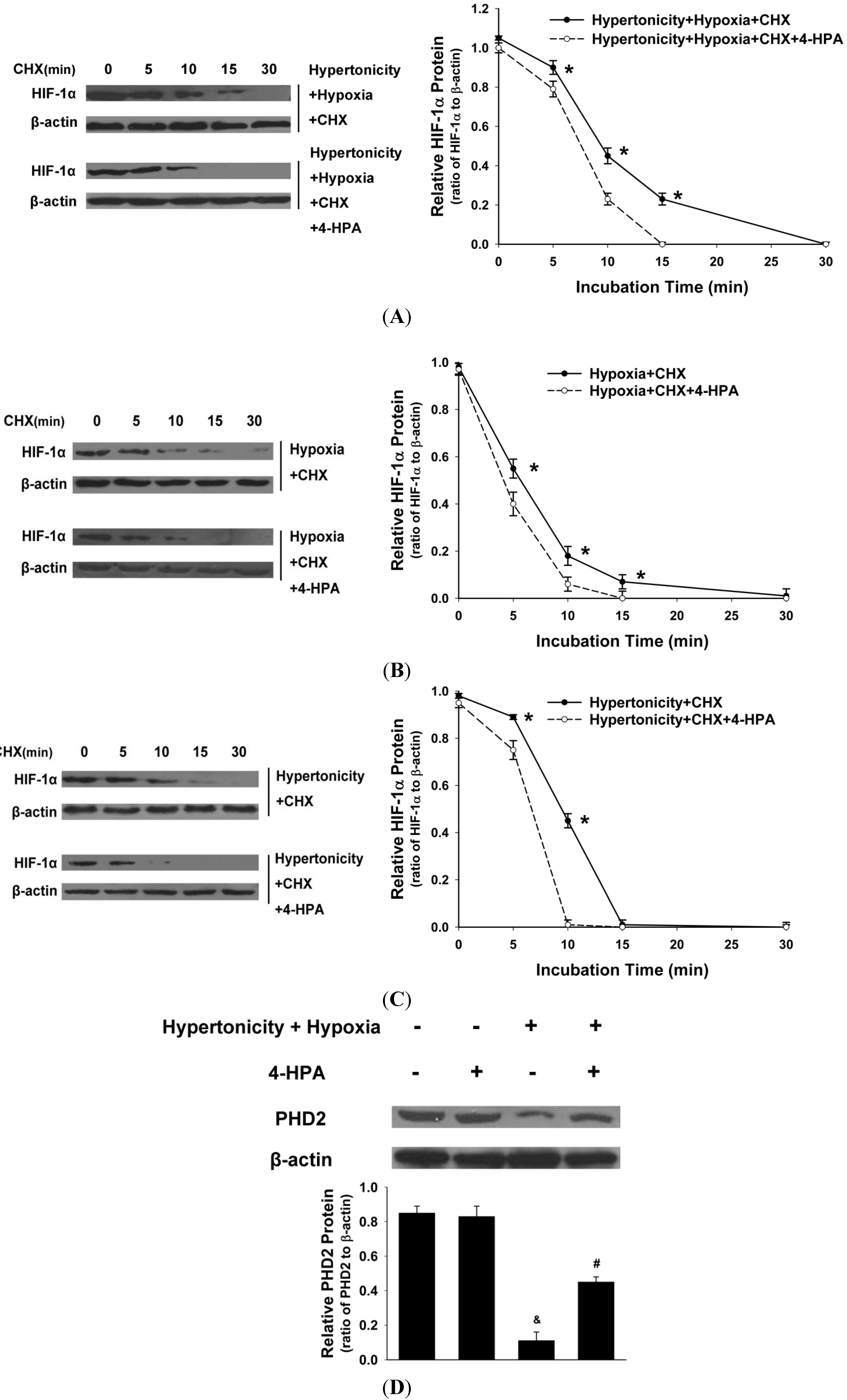
2.7. 4-HPA Decreased Hypertonicity and Hypoxia-Induced HIF-1α Protein Level through Promoting HIF-1α Protein Degradation, Which Was Associated with Prolyl Hydroxylase Domain Enzyme Isoform-2 (PHD2) Elevation in Primary AEC

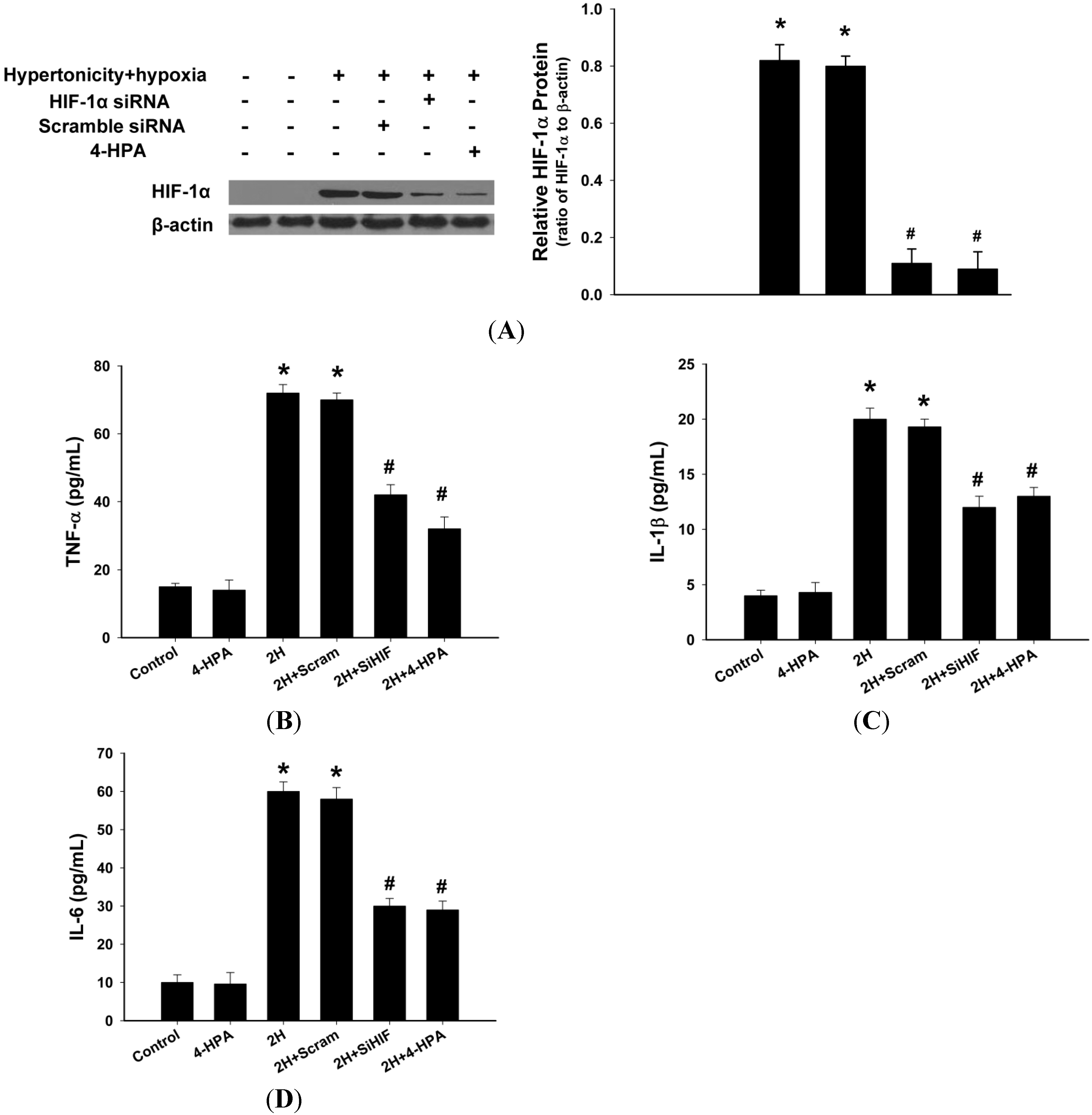
2.8. 4-HPA Decreased the Production of Inflammatory Cytokines through Suppressing Hypertonicity and Hypoxia-Induced HIF-1α in NR8383 Macrophages
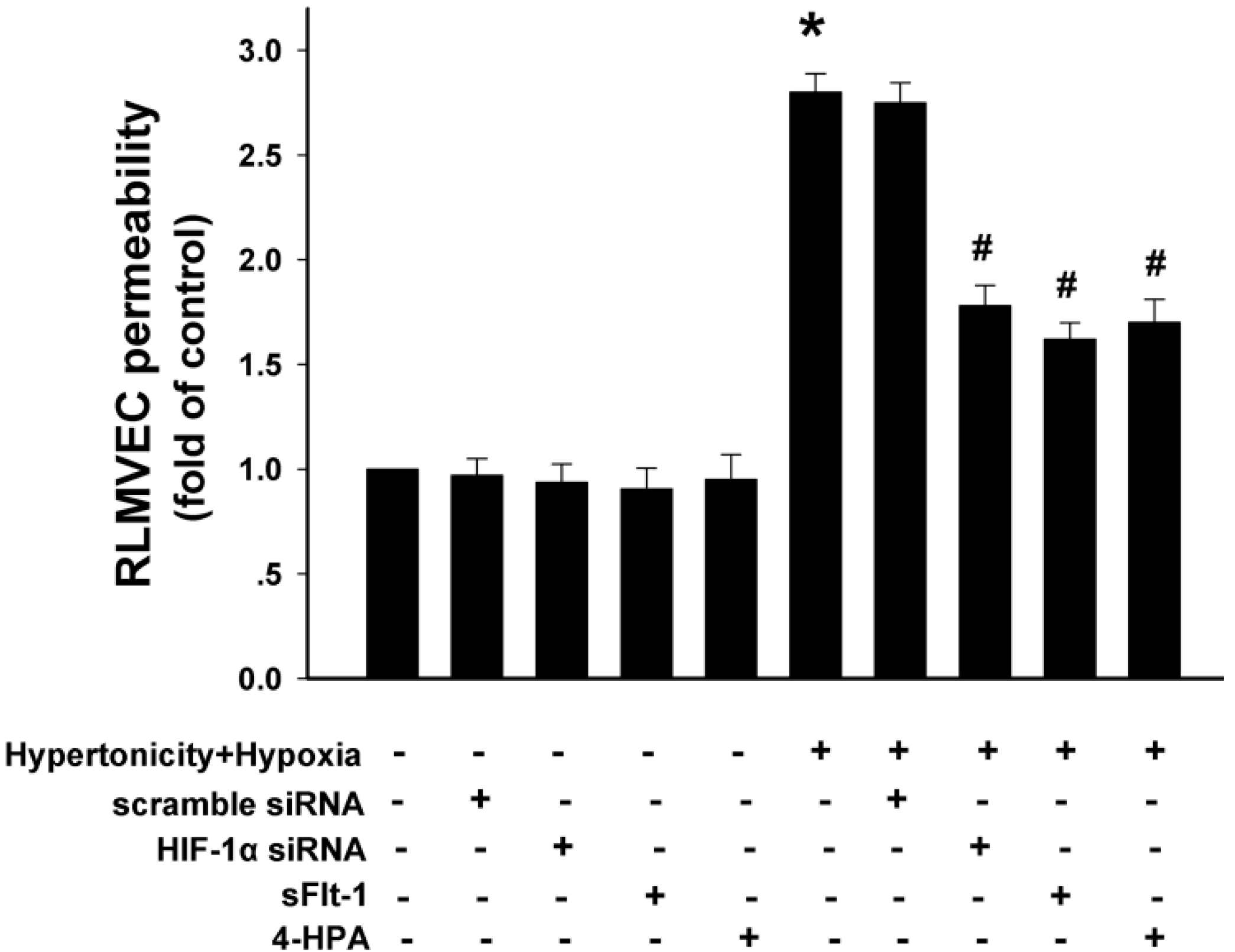
2.9. 4-HPA Decreased Monolayer Permeability through Suppressing Hypertonicity and Hypoxia-Induced HIF-1α, Which Was Mediated by Inhibiting VEGF in Rat Lung Microvascular Endothelial Cell Line (RLMVEC)


3. Discussion
4. Experimental Section
4.1. Animal Model and Grouping

4.2. PaO2 and PaCO2 Study
4.3. ELISA
4.4. Bronchoalveolar Lavage Fluid (BALF) White Cell Count
4.5. Assessment of Pulmonary Vascular Leakage
4.6. Wet-to-Dry Weight (W/D) Ratio
4.7. Histological Study
4.8. Cell Culture and Treatment
4.9. Plasmid Construction and Transfection
4.10. Permeability Assay
4.11. Western Blotting
4.12. Reverse Transcription-PCR
4.13. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Van Beeck, E.F.; Branche, C.M.; Szpilman, D.; Modell, J.H.; Bierens, J.J. A new definition of drowning: Towards documentation and prevention of a global public health problem. Bull. World Health Org. 2005, 83, 853–856. [Google Scholar]
- Li, J.; Xu, M.; Fan, Q.; Xie, X.; Zhang, Y.; Mu, D.; Zhao, P.; Zhang, B.; Cao, F.; Wang, Y.; et al. Tanshinone IIA ameliorates seawater exposure-induced lung injury by inhibiting aquaporins (AQP) 1 and AQP5 expression in lung. Respir. Physiol. Neurobiol. 2011, 176, 39–49. [Google Scholar] [CrossRef]
- Li, J.H.; Xu, M.; Xie, X.Y.; Fan, Q.X.; Mu, D.G.; Zhang, Y.; Cao, F.L.; Wang, Y.X.; Zhao, P.T.; Zhang, B.; et al. Tanshinone IIA suppresses lung injury and apoptosis, and modulates protein kinase B and extracellular signal-regulated protein kinase pathways in rats challenged with seawater exposure. Clin. Exp. Pharmacol. Physiol. 2011, 38, 269–277. [Google Scholar] [CrossRef]
- Fan, Q.; Zhao, P.; Li, J.; Xie, X.; Xu, M.; Zhang, Y.; Mu, D.; Li, W.; Sun, R.; Liu, W.; et al. 17β-Estradiol administration attenuates seawater aspiration-induced acute lung injury in rats. Pulm. Pharmacol. Ther. 2011, 24, 673–681. [Google Scholar] [CrossRef]
- Rui, M.; Duan, Y.Y.; Wang, H.L.; Zhang, X.H.; Wang, Y. Differences between seawater- and freshwater-induced lung injuries. Chin. Crit. Care Med. 2009, 21, 416–420. [Google Scholar]
- Wang, Q.; Zheng, X.; Cheng, Y.; Zhang, Y.L.; Wen, H.X.; Tao, Z.; Li, H.; Hao, Y.; Gao, Y.; Yang, L.M.; et al. Resolvin D1 stimulates alveolar fluid clearance through alveolar epithelial sodium channel, Na,K-ATPase via ALX/cAMP/PI3K pathway in lipopolysaccharide-induced acute lung injury. J. Immunol. 2014, 192, 3765–3777. [Google Scholar] [CrossRef]
- He, J.; Zhao, Y.; Deng, W.; Wang, D.X. Netrin-1 promotes epithelial sodium channel-mediated alveolar fluid clearance via activation of the adenosine 2B receptor in lipopolysaccharide-induced acute lung injury. Respiration 2014, 87, 394–407. [Google Scholar]
- Xie, X.Y.; Zhang, B.; Li, J.H.; Fan, Q.X.; Zhang, Y.; Mu, D.G.; Li, W.P.; Xu, M.; Zhao, P.T.; Jin, F.G.; et al. Sodium tanshinone iia sulfonate attenuates seawater aspiration-induced acute pulmonary edema by up-regulating Na+,K+-ATPase activity. Exp. Lung Res. 2011, 37, 482–491. [Google Scholar] [CrossRef]
- Fan, Y.; Dong, W.; He, J.; Du, L.; Xu, G.; Liu, X.; Pei, Y.; Fu, G.; Ma, G. The effect of HJ-1 NO–HFJV respirator on treating pulmonary edema caused by seawater drowning. J. Biomed. Eng. 2002, 19, 526–528. [Google Scholar]
- Zhao, X.W.; Zhang, J.P.; Huang, X.; Liu, Y.N. Effects of positive end expiratory pressure ventilation upon respiratory function and hydrophobic surfactants proteins in rabbit with seawater respiratory distress syndrome. Zhonghua Yi Xue Za Zhi 2009, 89, 3266–3270. [Google Scholar]
- Sawada, S.; Ichiba, S.; Itano, H.; Shoga, K.; Shimizu, N. Experimental study of partial liquid ventilation in the setting of acute respiratory failure induced by sea water lung lavage in rabbits. Acta Med. Okayama 1998, 52, 131–137. [Google Scholar]
- Wang, G.L.; Jiang, B.H.; Rue, E.A.; Semenza, G.L. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc. Natl. Acad. Sci. USA 1995, 92, 5510–5514. [Google Scholar] [CrossRef]
- Dery, M.A.; Michaud, M.D.; Richard, D.E. Hypoxia-inducible factor 1: Regulation by hypoxic and non-hypoxic activators. Int. J. Biochem. Cell Biol. 2005, 37, 535–540. [Google Scholar]
- Oh, Y.T.; Lee, J.Y.; Yoon, H.; Lee, E.H.; Baik, H.H.; Kim, S.S.; Ha, J.; Yoon, K.S.; Choe, W.; Kang, I. Lipopolysaccharide induces hypoxia-inducible factor-1α mRNA expression and activation via NADPH oxidase and Sp1-dependent pathway in BV2 murine microglial cells. Neurosci. Lett. 2008, 431, 155–160. [Google Scholar] [CrossRef]
- Shein, N.A.; Horowitz, M.; Alexandrovich, A.G.; Tsenter, J.; Shohami, E. Heat acclimation increases hypoxia-inducible factor 1α and erythropoietin receptor expression: Implication for neuroprotection after closed head injury in mice. J. Cereb. Blood Flow Metabol. 2005, 25, 1456–1465. [Google Scholar] [CrossRef]
- Zhou, B.; Ann, D.K.; Li, X.; Kim, K.J.; Lin, H.; Minoo, P.; Crandall, E.D.; Borok, Z. Hypertonic induction of aquaporin-5: Novel role of hypoxia-inducible factor-1α. Am. J. Physiol. Cell Physiol. 2007, 292, C1280–C1290. [Google Scholar]
- Yeh, C.H.; Cho, W.; So, E.C.; Chu, C.C.; Lin, M.C.; Wang, J.J.; Hsing, C.H. Propofol inhibits lipopolysaccharide-induced lung epithelial cell injury by reducing hypoxia-inducible factor-1α expression. Br. J. Anaesth. 2011, 106, 590–599. [Google Scholar] [CrossRef]
- Hellwig-Burgel, T.; Stiehl, D.P.; Wagner, A.E.; Metzen, E.; Jelkmann, W. Review: Hypoxia-inducible factor-1 (HIF-1): A novel transcription factor in immune reactions. J. Interferon Cytokine Res. 2005, 25, 297–310. [Google Scholar] [CrossRef]
- Irwin, D.C.; McCord, J.M.; Nozik-Grayck, E.; Beckly, G.; Foreman, B.; Sullivan, T.; White, M.; Crossno, J.T., Jr.; Bailey, D.; Flores, S.C.; et al. A potential role for reactive oxygen species and the HIF-1α-VEGF pathway in hypoxia-induced pulmonary vascular leak. Free Radic. Biol. Med. 2009, 47, 55–61. [Google Scholar] [CrossRef]
- Ng, T.B.; Liu, F.; Lu, Y.; Cheng, C.H.; Wang, Z. Antioxidant activity of compounds from the medicinal herb Aster tataricus. Comp. Biochem. Physiol. 2003, 136, 109–115. [Google Scholar]
- Du, L.; Mei, H.F.; Yin, X.; Xing, Y.Q. Delayed growth of glioma by a polysaccharide from Aster tataricus involve upregulation of Bax/Bcl-2 ratio, activation of caspase-3/8/9, and downregulation of the Akt. Tumour Biol. 2014, 35, 1819–1825. [Google Scholar] [CrossRef]
- Yen, M.H.; Lee, J.J.; Yeh, C.F.; Wang, K.C.; Chiang, Y.W.; Chiang, L.C.; Chang, J.S. Yakammaoto inhibited human coxsackievirus B4 (CVB4)-induced airway and renal tubular injuries by preventing viral attachment, internalization, and replication. J. Ethnopharmacol. 2014, 151, 1056–1063. [Google Scholar] [CrossRef]
- Cloward, J.M.; Shafer, W.M. MtrR control of a transcriptional regulatory pathway in Neisseria meningitidis that influences expression of a gene (NadA) encoding a vaccine candidate. PLoS One 2013, 8, e56097. [Google Scholar] [CrossRef]
- Metruccio, M.M.; Pigozzi, E.; Roncarati, D.; Berlanda Scorza, F.; Norais, N.; Hill, S.A.; Scarlato, V.; Delany, I. A novel phase variation mechanism in the meningococcus driven by a ligand-responsive repressor and differential spacing of distal promoter elements. PLoS Pathog. 2009, 5, e1000710. [Google Scholar] [CrossRef]
- Wen, K.C.; Chang, C.S.; Chien, Y.C.; Wang, H.W.; Wu, W.C.; Wu, C.S.; Chiang, H.M. Tyrosol and its analogues inhibit α-melanocyte-stimulating hormone induced melanogenesis. Int. J. Mol. Sci. 2013, 14, 23420–23440. [Google Scholar]
- Lang, K.J.; Kappel, A.; Goodall, G.J. Hypoxia-inducible factor-1α mRNA contains an internal ribosome entry site that allows efficient translation during normoxia and hypoxia. Mol. Biol. Cell 2002, 13, 1792–1801. [Google Scholar]
- Laughner, E.; Taghavi, P.; Chiles, K.; Mahon, P.C.; Semenza, G.L. HER2 (neu) signaling increases the rate of hypoxia-inducible factor 1α (HIF-1α) synthesis: Novel mechanism for HIF-1-mediated vascular endothelial growth factor expression. Mol. Cell. Biol. 2001, 21, 3995–4004. [Google Scholar] [CrossRef]
- Page, E.L.; Robitaille, G.A.; Pouyssegur, J.; Richard, D.E. Induction of hypoxia-inducible factor-1α by transcriptional and translational mechanisms. J. Biol. Chem. 2002, 277, 48403–48409. [Google Scholar]
- Zhang, B.; Liu, Z.Y.; Li, Y.Y.; Luo, Y.; Liu, M.L.; Dong, H.Y.; Wang, Y.X.; Liu, Y.; Zhao, P.T.; Jin, F.G.; et al. Antiinflammatory effects of matrine in LPS-induced acute lung injury in mice. Eur. J. Pharm. Sci. 2011, 44, 573–579. [Google Scholar] [CrossRef]
- Iba, T.; Nagaoka, I.; Yamada, A.; Nagayama, M.; Miki, T. Effect of hemoperfusion using polymyxin B-immobilized fibers on acute lung injury in a rat sepsis model. Int. J. Med. Sci. 2014, 11, 255–261. [Google Scholar] [CrossRef]
- Gao, J.; Zhan, Y.; Chen, J.; Wang, L.; Yang, J. Triptolide ameliorates lipopolysaccharide-induced acute lung injury in rats. Eur. J. Med. Res. 2014. [Google Scholar] [CrossRef]
- Mi, Z.; Rapisarda, A.; Taylor, L.; Brooks, A.; Creighton-Gutteridge, M.; Melillo, G.; Varesio, L. Synergystic induction of HIF-1α transcriptional activity by hypoxia and lipopolysaccharide in macrophages. Cell Cycle 2008, 7, 232–241. [Google Scholar] [CrossRef]
- Semenza, G.L. HIF-1 mediates metabolic responses to intratumoral hypoxia and oncogenic mutations. J. Clin. Investig. 2013, 123, 3664–3671. [Google Scholar] [CrossRef]
- Vadlapatla, R.K.; Vadlapudi, A.D.; Mitra, A.K. Hypoxia-inducible factor-1 (HIF-1): A potential target for intervention in ocular neovascular diseases. Curr. Drug Targets 2013, 14, 919–935. [Google Scholar] [CrossRef]
- Nangaku, M.; Rosenberger, C.; Heyman, S.N.; Eckardt, K.U. Regulation of hypoxia-inducible factor in kidney disease. Clin. Exp. Pharmacol. Physiol. 2013, 40, 148–157. [Google Scholar] [CrossRef]
- Liu, L.; Ning, X.; Sun, L.; Shi, Y.; Han, S.; Guo, C.; Chen, Y.; Sun, S.; Yin, F.; Wu, K.; et al. Involvement of MGr1-Ag/37LRP in the vincristine-induced HIF-1 expression in gastric cancer cells. Mol. Cell. Biochem. 2007, 303, 151–160. [Google Scholar] [CrossRef]
- Stel, V.S.; Dekker, F.W.; Tripepi, G.; Zoccali, C.; Jager, K.J. Survial analysis I: the Kaplan-Meier method. Nephron. Clin. Pract. 2011, 119, 83–88. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, Z.; Xi, R.; Zhang, Z.; Li, W.; Liu, Y.; Jin, F.; Wang, X. 4-Hydroxyphenylacetic Acid Attenuated Inflammation and Edema via Suppressing HIF-1α in Seawater Aspiration-Induced Lung Injury in Rats. Int. J. Mol. Sci. 2014, 15, 12861-12884. https://doi.org/10.3390/ijms150712861
Liu Z, Xi R, Zhang Z, Li W, Liu Y, Jin F, Wang X. 4-Hydroxyphenylacetic Acid Attenuated Inflammation and Edema via Suppressing HIF-1α in Seawater Aspiration-Induced Lung Injury in Rats. International Journal of Molecular Sciences. 2014; 15(7):12861-12884. https://doi.org/10.3390/ijms150712861
Chicago/Turabian StyleLiu, Zhongyang, Ronggang Xi, Zhiran Zhang, Wangping Li, Yan Liu, Faguang Jin, and Xiaobo Wang. 2014. "4-Hydroxyphenylacetic Acid Attenuated Inflammation and Edema via Suppressing HIF-1α in Seawater Aspiration-Induced Lung Injury in Rats" International Journal of Molecular Sciences 15, no. 7: 12861-12884. https://doi.org/10.3390/ijms150712861