β-Defensin 2 Ameliorates Lung Injury Caused by Pseudomonas Infection and Regulates Proinflammatory and Anti-Inflammatory Cytokines in Rat

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Construction and Analysis of cDNA and shRNA Expressing Vectors and Detection of Inhibitory Efficiencies

2.3. Histological Changes in Rat Lungs of rBD2 Over-Expression and Knockdown

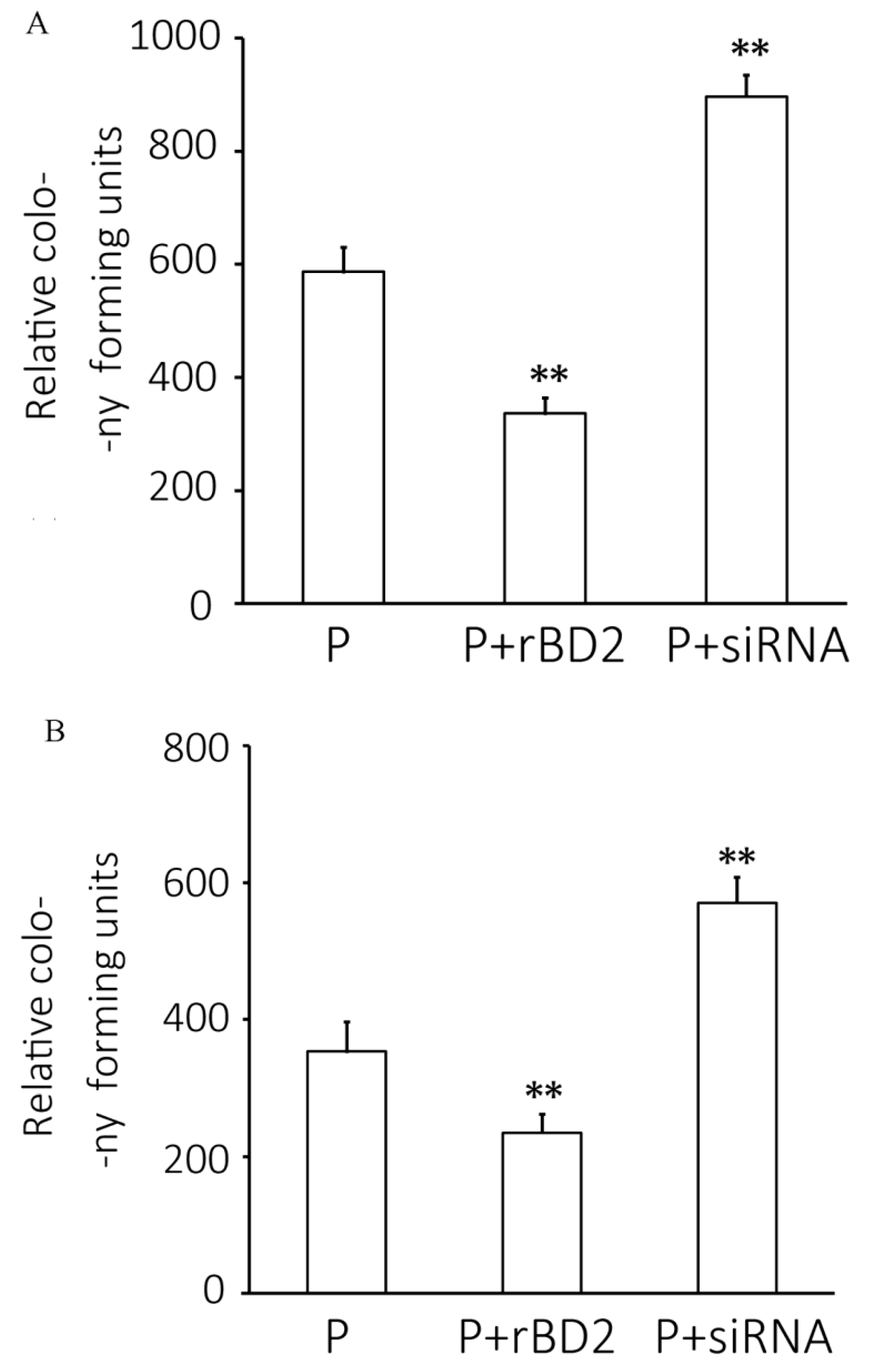
2.4. Pseudomonas aeruginosa Infection Level

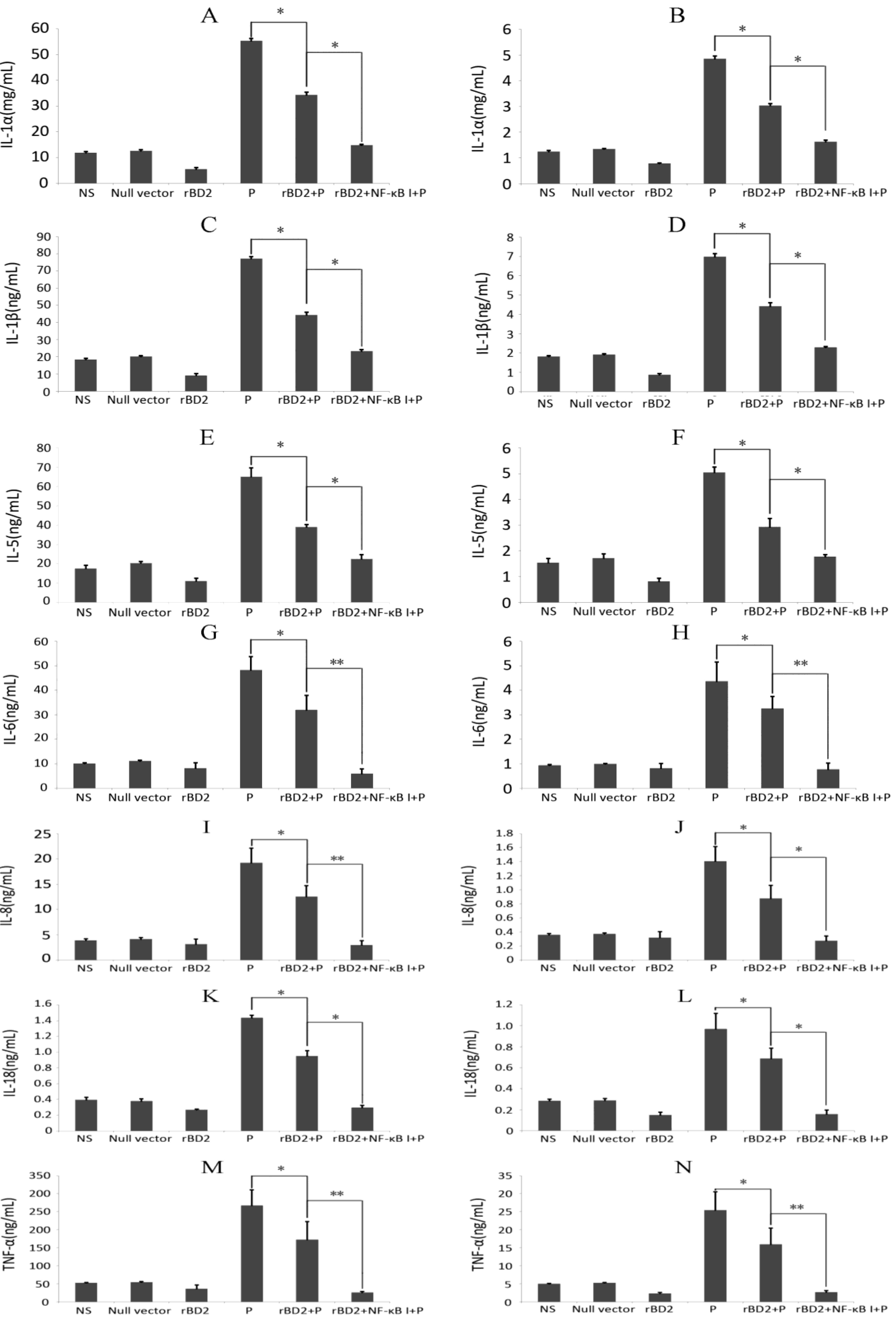
2.5. Effects of rBD2 on Cytokines


3. Discussion
4. Experimental Section
4.1. Expressing Vectors Construction, Virus Package and Infection
4.2. Animal Grouping and Treatment
4.3. Determination of rBD2 mRNA in Rat Lungs by Quantitative RT PCR
4.4. Western Blotting
4.5. Pathological Examination
4.6. Determination of Pseudomonas Infection Levels
4.7. Determination of Cytokines in Lung Tissues and Peripheral Blood
4.8. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rober, B. Epithelial antimicrobial peptides in host defense against infection. Respir. Res. 2000, 3, 141–150. [Google Scholar]
- Sang, Y.; Blecha, F. Antimicrobial peptides and bacteriocins: Alternatives to traditional antibiotics. Anim. Health Res. Rev. 2008, 9, 227–235. [Google Scholar]
- Wu, M.; McClellan, S.A.; Barrett, R.P.; Zhang, Y.; Hazlett, L.D. β-defensins 2 and 3 together promote resistance to Pseudomonas aeruginosa keratitis. J. Immunol. 2009, 183, 8054–8060. [Google Scholar]
- Guaní-Guerra, E.; Negrete-García, M.C.; Montes-Vizuet, R.; Asbun-Bojalil, J.; Terán, L.M. Human b-Defensin-2 induction in nasal mucosa after administration of bacterial lysates. Arch. Med. Res. 2011, 42, 189–194. [Google Scholar] [CrossRef]
- Kraus, D.; Deschner, J.; Jäger, A.; Wenghoefer, M.; Bayer, S.; Jepsen, S.; Allam, J.P.; Novak, N.; Meyer, R.; Winter, J. Human β-defensins differently affect proliferation, differentiation, and mineralization of osteoblast-like MG63 cells. J. Cell. Physiol. 2012, 227, 994–1003. [Google Scholar] [CrossRef]
- Kohlgraf, K.G.; Pingel, L.C.; Dietrich, D.E.; Brogden, K.A. Defensins as anti-inflammatory compounds and mucosal adjuvants. Future Microbiol. 2010, 5, 99–113. [Google Scholar] [CrossRef]
- Jia, H.P.; Mills, J.N.; Barahmand-Pour, F.; Nishimura, D.; Mallampali, R.K.; Wang, G.; Wiles, K.; Tack, B.F.; Bevins, C.L.; McCray, P.B., Jr. Molecular cloning and characterization of rat genes encoding homologues of human b-defensins. Infect. Immun. 1999, 67, 4827–4833. [Google Scholar]
- Pluta, K.; Kacprzak, M.M. Use of HIV as a gene transfer vector. Acta Biochim. Pol. 2009, 56, 531–535. [Google Scholar]
- Lois, C.; Hong, E.J.; Pease, S.; Brown, E.J.; Baltimore, D. Germline transmission and tissue-specific expression of transgenes delivered by lentiviral vectors. Science 2002, 295, 868–872. [Google Scholar] [CrossRef]
- Pfeifer, A.; Ikawa, M.; Dayn, Y.; Verma, I.M. Transgenesis by lentiviral vectors, lack of gene silencing in mammalian embryonic stem cells and preimp lantation embryos. Proc. Natl. Acad. Sci. USA 2002, 99, 2140–2145. [Google Scholar] [CrossRef]
- Lai, Z.; Brady, R.O. Gene transfer into the central nervous system in vivo using a recombinant lentivirus vector. J. Neurosci. Res. 2002, 67, 363–371. [Google Scholar] [CrossRef]
- Yu, X.; Zhan, X.; D’Costa, J.; Tanavde, V.M.; Ye, Z.; Peng, T.; Malehorn, M.T.; Yang, X.; Civin, C.I.; Cheng, L. Lentiviral vectors with two independent internal promoters transfer high-level expression of multiple transgenes to human hematopoietic stem-progenitor cells. Mol. Ther. 2003, 7, 827–838. [Google Scholar] [CrossRef]
- Cockrell, A.S.; Kafri, T. Gene delivery by lentivirus vectors. Mol. Biotechnol. 2007, 36, 184–204. [Google Scholar] [CrossRef]
- Yang, D.; Chertov, O.; Bykovskaia, S.N.; Chen, Q.; Buffo, M.J.; Shogan, J.; Anderson, M.; Schröder, J.M.; Wang, J.M.; Howard, O.M.; et al. β-defensins: Linking innate and adaptive immunity through dendritic and T cell CCR6. Science 1999, 286, 525–528. [Google Scholar]
- Niyonsaba, F.; Ogawa, H.; Nagaoka, I. Human β-defensin-2 functions as a chemotactic agent for tumour necrosis factor-alpha-treated human neutrophils. Immunology 2004, 111, 273–281. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Iwabuchi, K.; Matsuda, H.; Ogawa, H.; Nagaoka, I. Epithelial cell-derived human β-defensin-2 acts as a chemotaxin for mast cells through a pertussis toxin-sensitive and phospholipase C-dependent pathway. Int. Immunol. 2002, 14, 421–426. [Google Scholar] [CrossRef]
- Liu, K.X.; Chen, S.Q.; Zhang, H.; Guo, J.Y.; Li, Y.S.; Huang, W.Q. Intestinal ischaemia/reperfusion upregulates β-defensin-2 expression and causes acute lung injury in the rat. Injury 2009, 40, 950–955. [Google Scholar]
- Ashitani, J.; Mukae, H.; Ihiboshi, H.; Taniguchi, H.; Mashimoto, H.; Nakazato, M.; Matsukura, S. Defensin in plasma and in bronchoalveolar lavage fluid from patients with acute respiratory distress syndrome. Nihon Kyobu Shikkan Gakkai Zasshi 1996, 34, 1349–1353. [Google Scholar]
- Sakamoto, N.; Mukae, H.; Fujii, T.; Ishii, H.; Yoshioka, S.; Kakugawa, T.; Sugiyama, K.; Mizuta, Y.; Kadota, J.; Nakazato, M.; et al. Differential effects of α- and β-defensin on cytokine production by cultured human bronchial epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 288, 508–513. [Google Scholar]
- Hu, Q.; Zuo, P.; Shao, B.; Yang, S.; Xu, G.; Lan, F.; Lu, X.; Xiong, W.; Xu, Y.; Xiong, S. Administration of nonviral gene vector encoding rat β-defensin-2 ameliorates chronic Pseudomonasaeruginosa lung infection in rats. J. Gene Med. 2010, 12, 276–286. [Google Scholar]
- Wu, M.; McClellan, S.A.; Barrett, R.P.; Hazlett, L.D. β-defensin-2 promotes resistance against infection with P. aeruginosa. J. Immunol. 2009, 182, 1609–1616. [Google Scholar]
- Bonfield, T.L.; Panuska, J.R.; Konstan, M.W.; Hilliard, K.A.; Hilliard, J.B.; Ghnaim, H.; Berger, M. Inflammatory cytokines in cystic fibrosis lungs. Am. J. Respir. Crit. Care Med. 1995, 152, 2111–2118. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar]
- Biragyn, A.; Ruffini, P.A.; Leifer, C.A.; Klyushnenkova, E.; Shakhov, A.; Chertov, O.; Shirakawa, A.K.; Farber, J.M.; Segal, D.M.; Oppenheim, J.J.; et al. Toll-like receptor 4-dependent activation of dendritic cells by β-defensin 2. Science 2002, 298, 1025–1029. [Google Scholar] [CrossRef]
- Boniotto, M.; Jordan, W.J.; Eskdale, J.; Tossi, A.; Antcheva, N.; Crovella, S.; Connell, N.D.; Gallagher, G. Human β-defensin 2 induces a vigorous cytokine response in peripheral blood mononuclear cells. Antimicrob. Agents Chemother. 2006, 50, 1433–1441. [Google Scholar] [CrossRef]
- Tsutsumi-Ishii, Y.; Nagaoka, I. NF-kappa B-mediated transcriptional regulation of human β-defensin-2 gene following lipopolysaccharide stimulation. J. Leukoc. Biol. 2002, 71, 154–162. [Google Scholar]
- Shao, Z.J.; Zheng, X.W.; Feng, T.; Huang, J.; Chen, J.; Wu, Y.Y.; Zhou, L.M.; Tu, W.W.; Li, H. Andrographolide exerted its antimicrobial effects by upregulation of human β-defensin-2 induced through p38 MAPK and NF-κB pathway in human lung epithelial cells. Can. J. Physiol. Pharmacol. 2012, 90, 647–653. [Google Scholar] [CrossRef]
- Wada, A.; Ogushi, K.; Kimura, T.; Hojo, H.; Mori, N.; Suzuki, S.; Kumatori, A.; Se, M.; Nakahara, Y.; Nakamura, M.; et al. Helicobacter pylori-mediated transcriptional regulation of the human β-defensin 2 gene requires NF-kappaB. Cell. Microbiol. 2001, 3, 115–123. [Google Scholar] [CrossRef]
- Philip, N.H.; Dillon, C.P.; Snyder, A.G.; Fitzgerald, P.; Wynosky-Dolfi, M.A.; Zwack, E.E.; Hu, B.; Fitzgerald, L.; Mauldin, E.A.; Copenhaver, A.M.; et al. Caspase-8 mediates caspase-1 processing and innate immune defense in response to bacterial blockade of NF-κB and MAPK signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7385–7390. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shen, Z.; Fang, L.; Zhao, L.; Lei, H. β-Defensin 2 Ameliorates Lung Injury Caused by Pseudomonas Infection and Regulates Proinflammatory and Anti-Inflammatory Cytokines in Rat. Int. J. Mol. Sci. 2014, 15, 13372-13387. https://doi.org/10.3390/ijms150813372
Shen Z, Fang L, Zhao L, Lei H. β-Defensin 2 Ameliorates Lung Injury Caused by Pseudomonas Infection and Regulates Proinflammatory and Anti-Inflammatory Cytokines in Rat. International Journal of Molecular Sciences. 2014; 15(8):13372-13387. https://doi.org/10.3390/ijms150813372
Chicago/Turabian StyleShen, Zhenwei, Lu Fang, Liming Zhao, and Han Lei. 2014. "β-Defensin 2 Ameliorates Lung Injury Caused by Pseudomonas Infection and Regulates Proinflammatory and Anti-Inflammatory Cytokines in Rat" International Journal of Molecular Sciences 15, no. 8: 13372-13387. https://doi.org/10.3390/ijms150813372