Molecular Cloning and Functional Characterization of the Lycopene ?-Cyclase Gene via Virus-Induced Gene Silencing and Its Expression Pattern in Nicotiana tabacum


Abstract
:1. Introduction
2. Results
2.1. Isolation and Characterization of Ntε-LCY Genes

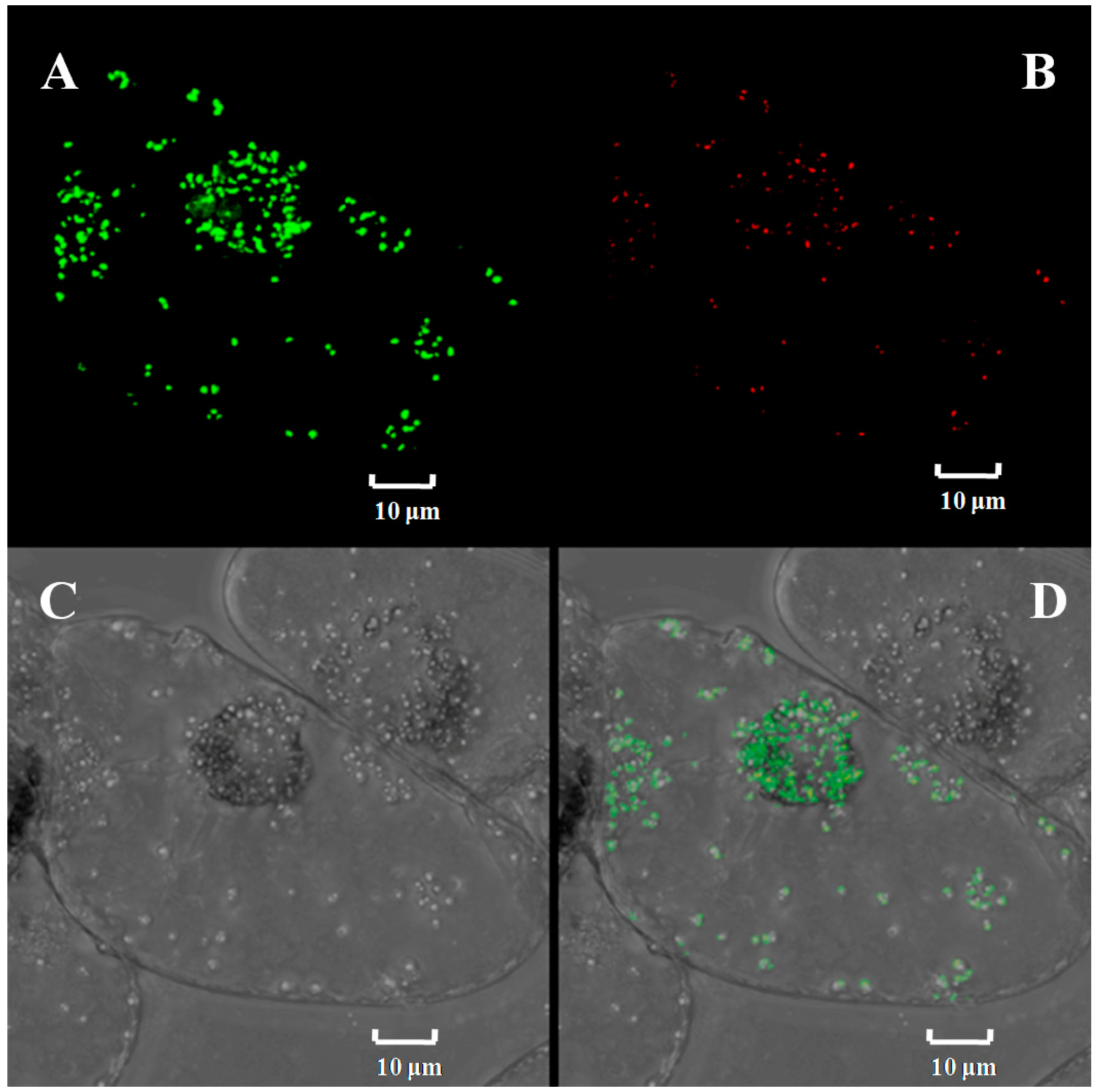
2.2. Subcellular Localization of Ntε-LCY1 Mature Protein


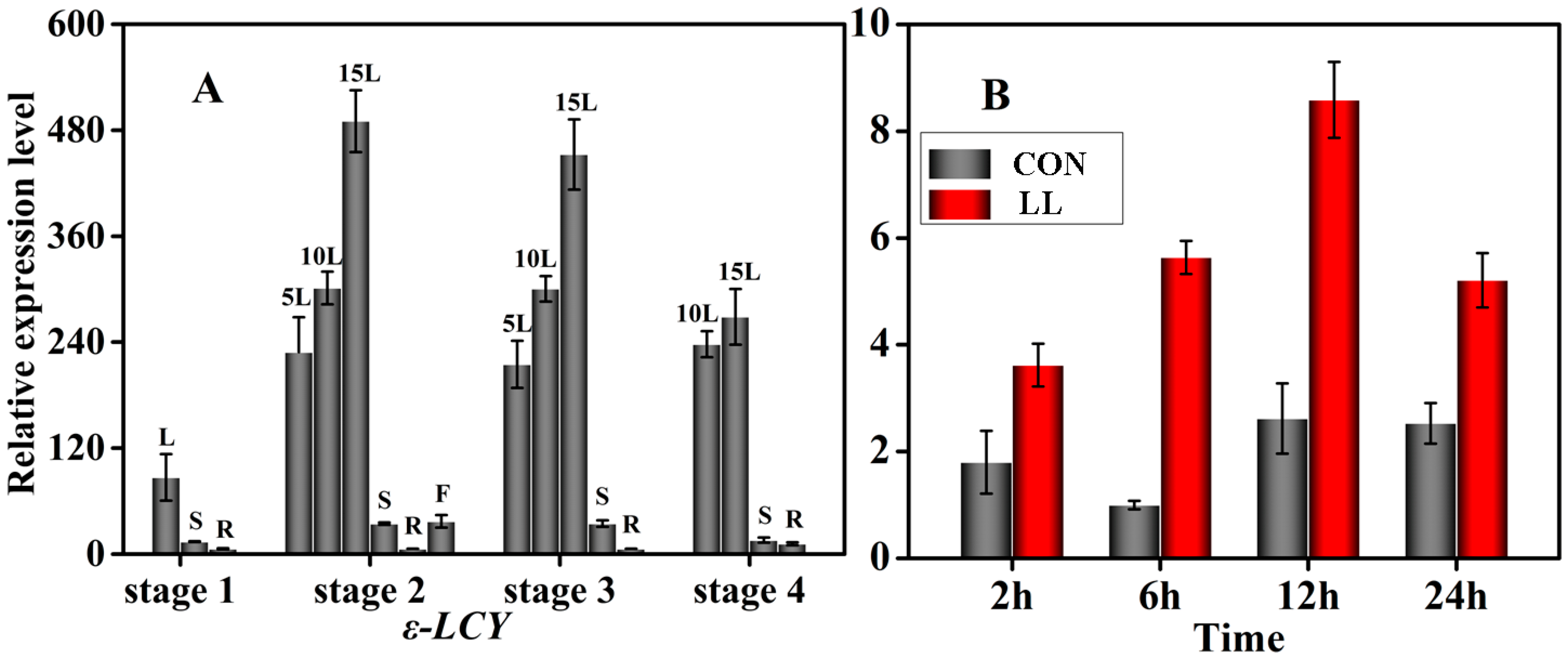
2.3. Transcriptional Expression Patterns of Ntε-LCY

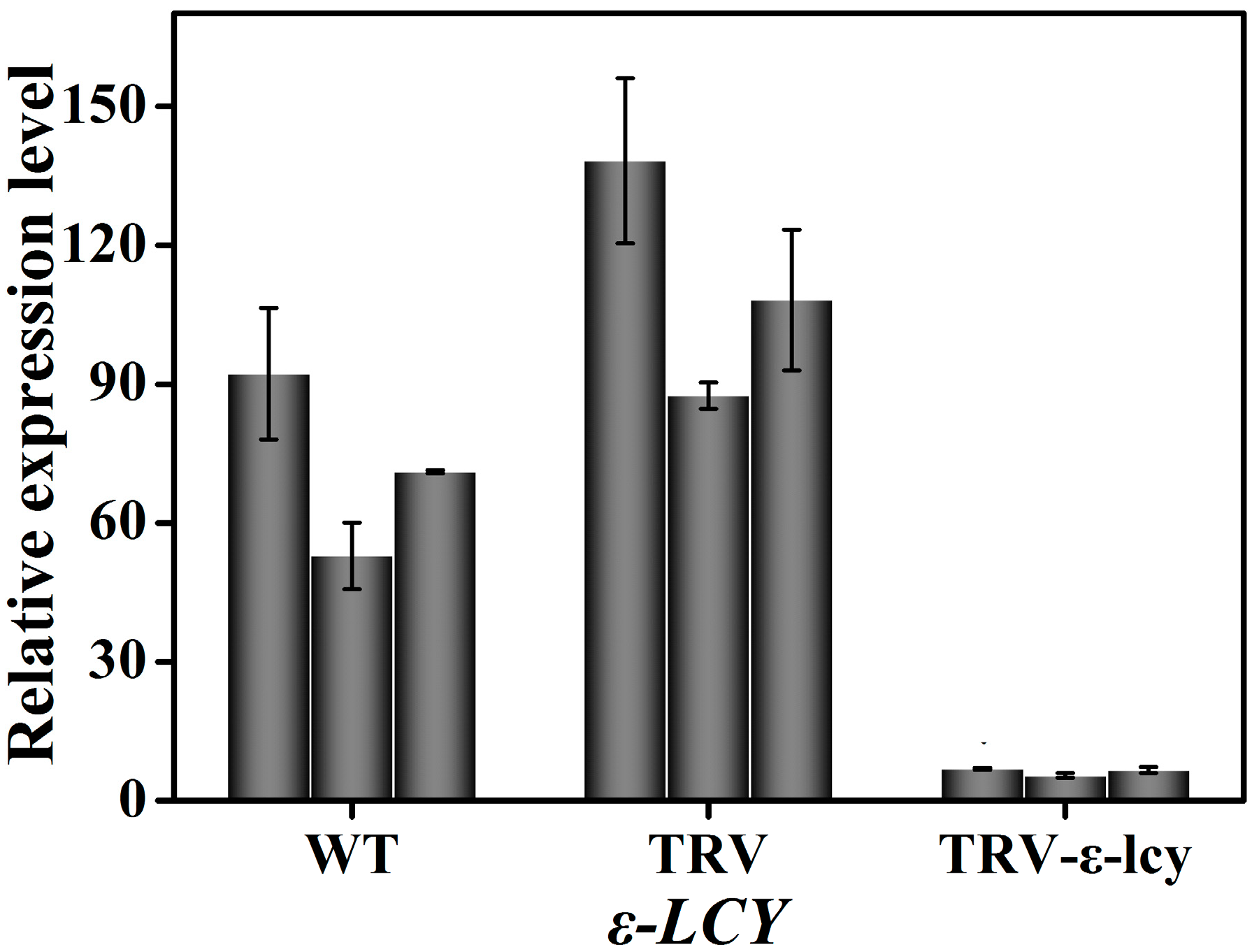
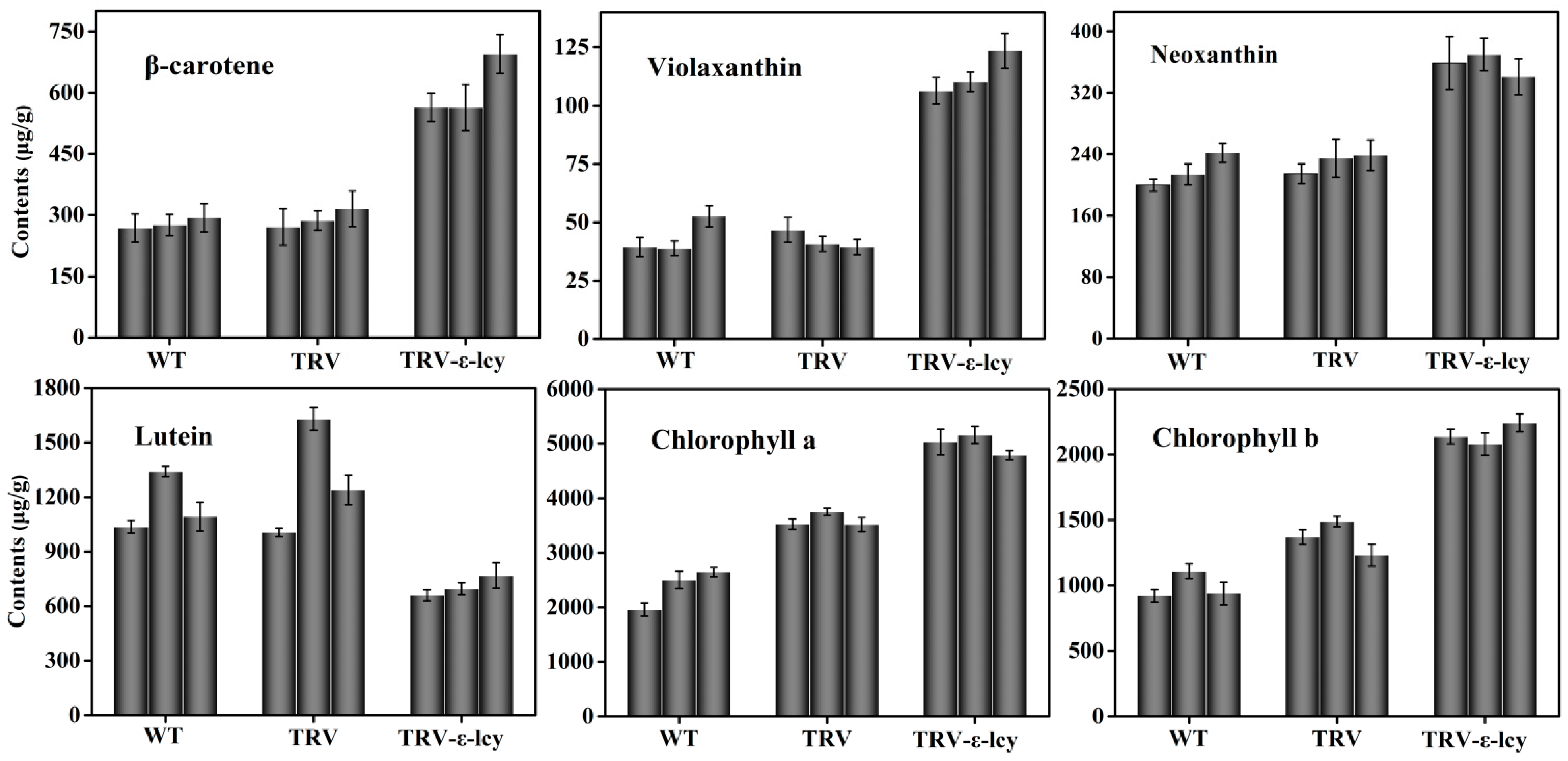
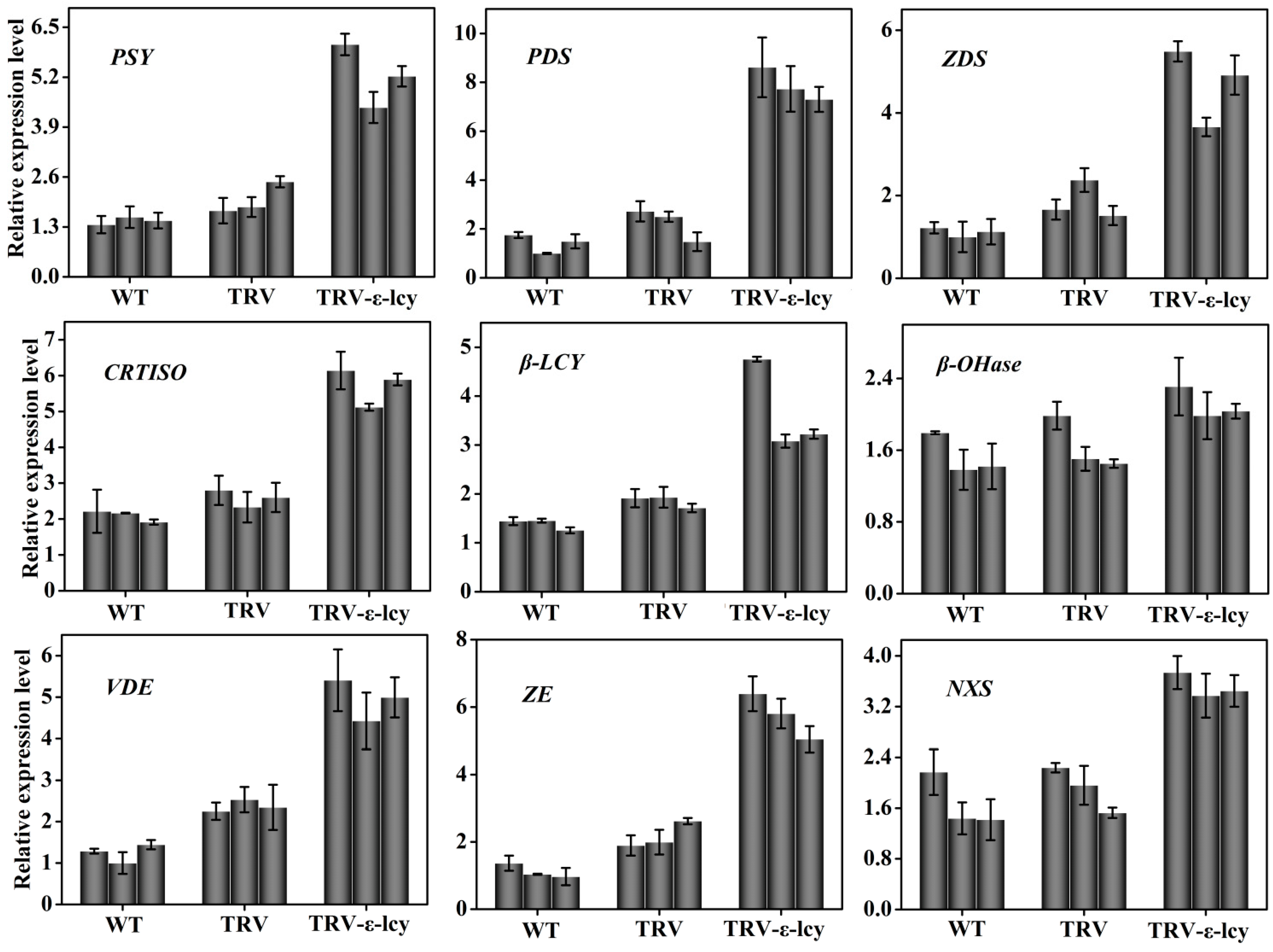
2.4. Manipulation of ε-LCY Expression Levels in Nicotiana Benthamiana and Its Effects on Carotenoid Content




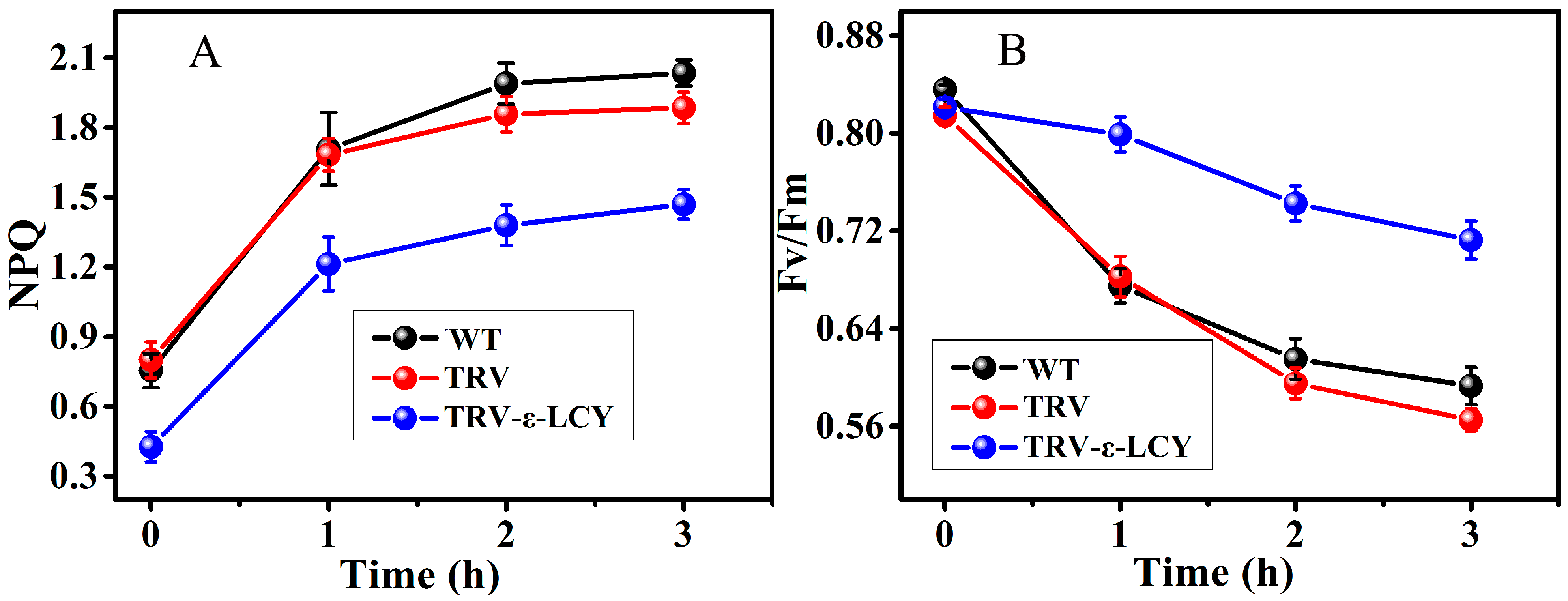
2.5. Chlorophyll Fluorescence Analysis

3. Discussion
4. Experimental Section
4.1. Plant Materials
4.2. BY-2 Cell Culture Conditions and Growth Measurement
4.3. Genomic DNA and RNA Isolation and cDNA Synthesis
4.4. Cloning of ε-LCY and Vector Construction
4.5. Evolutionary Analysis of ε-LCY
4.6. BY-2 Suspension Cell Bombardment and Fluorescence Microscopy
4.7. Agrobacterium Transformation and Infiltration
4.8. Gene Expression Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Primer Sequence |
|---|---|---|
| PSY | PSY-Q-F | TGTTGGAGAAGATGCCAGAAGAG |
| PSY-Q-R | ATAAGCAATAGGTAAGGAAATTAGCTTC | |
| PDS | PDS-Q-F | ATAAACCCTGACGAGCTTTC |
| PDS-Q-R | AATATGTTCAACAATCGGCAT | |
| ZDS | ZDS-Q-R | TGAAATAGGGGAGCTTGATTTCCGC |
| ZDS-Q-F | GAGCATATGCGACAGGATCCCAC | |
| CRTISO | CRTISO-Q-F | CGTGTACACCGAGAATATGATG |
| CRTISO-Q-R | GTAGGCGAGAGTCAAGCACTC | |
| β-LCY | β-LCY-Q-F | GATGACAATACAACTAAAGATCTTGATAG |
| β-LCY-Q-R | CATAAGCTACTTGATATCCAGGAT | |
| ε-LCY | ε-LCY-Q-F | CAGGAGTCTTTTTCGAGGAAACTTG |
| ε-LCY-Q-R | GTGTTCCAAGCTTGAGTTGAGAT | |
| β-OHase | β-OHase-Q-F | ATGGCCGCCAGCAGAATTTC |
| β-OHase-Q-R | CTCAATTTTCATTTCAATCTCCTCTGTC | |
| VDE | VDE-Q-F | ATGATGCATGGGATGGATATG |
| VDE-Q-R | CGTTGGAGCTCTTTAAAACCTTC | |
| ZE | ZE-Q-F | GTGGTGGGATTGGAGGGTTAGTG |
| ZE-Q-R | AGGATCTGCTGCAAAGTCATGC | |
| NXS | NXS-Q-F | GCCGGGCTCTATTCGACGTGAT |
| NXS-Q-R | ACTGACTCTACCATATGGTCTTCCCAAAT | |
| 26S-RNA | 26S-RNA-Q-F | GAAGAAGGTCCCAAGGGTTC |
| 26S-RNA-Q-R | TCTCCCTTTAACACCAACGG |
4.9. Carotenoid Extraction and Quantification
4.10. In Vivo Fluorescence and Non-Photochemical Quenching (NPQ) Measurements
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Supplementary Files
Supplementary File 1Supplementary File 2Supplementary File 3Supplementary File 4References
- Tao, N.; Hu, Z.; Liu, Q.; Xu, J.; Cheng, Y.; Guo, L.; Guo, W.; Deng, X. Expression of phytoene synthase gene (Psy) is enhanced during fruit ripening of Cara Cara navel orange (Citrus sinensis Osbeck). Plant Cell Rep. 2007, 26, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Nambara, E.; Marion-Poll, A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef] [PubMed]
- Taylor, I.B.; Sonneveld, T.; Bugg, T.D.H.; Thompson, A.J. Regulation and manipulation of the biosynthesis of abscisic acid, including the supply of xanthophyll precursors. J. Plant Growth Regul. 2005, 24, 253–273. [Google Scholar]
- Gomez-Roldan, V.; Fermas, S.; Brewer, P.B.; Puech-Pages, V.; Dun, E.A.; Pillot, J.P.; Letisse, F.; Matusova, R.; Danoun, S.; Portais, J.-C.; et al. Strigolactone inhibition of shoot branching. Nature 2008, 455, 189–194. [Google Scholar]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Takeda-Kamiya, N.; Magome, H.; Kamiya, Y.; Shirasu, K.; Yoneyama, K.; et al. Inhibition of shoot branching by new terpenoid plant hormones. Nature 2008, 455, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Matusova, R.; Rani, K.; Verstappen, F.W.; Franssen, M.C.; Beale, M.H.; Bouwmeester, H.J. The strigolactone germination stimulants of the plant-parasitic Striga and Orobanche spp. are derived from the carotenoid pathway. Plant Physiol. 2005, 139, 920–934. [Google Scholar]
- Aluru, M.; Xu, Y.; Guo, R.; Wang, Z.; Li, S.; White, W.; Wang, K.; Rodermel, S. Generation of transgenic maize with enhanced provitamin A content. J. Exp. Bot. 2008, 59, 3551–3562. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.M. Genetic modification of plant metabolism for human health benefits. Mutat. Res. 2007, 622, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Regal, P.; Amorim-Carrilho, K.T.; Cepeda, A.; Fente, C. Review of methods for analysis of carotenoids. Trends Anal. Chem. 2014, 56, 49–73. [Google Scholar] [CrossRef]
- DellaPenna, D.; Pogson, B.J. Vitamin synthesis in plants: Tocopherols and carotenoids. Annu. Rev. Plant Biol. 2006, 57, 711–738. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.J. The role of carotenoids in human health. Nutr. Clin. Care 2002, 5, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Aspects Med. 2005, 26, 459–516. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, G.; Tavazza, R.; Diretto, G.; Beyer, P.; Taylor, M.A. Metabolic engineering of carotenoid biosynthesis in plants. Trends Biotechnol. 2008, 26, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Lydiate, D.J.; Young, L.W.; Schafer, U.A.; Hannoufa, A. Enhancing the carotenoid content of Brassica napus seeds by down-regulating lycopene-ε-cyclase. Transgenic Res. 2008, 17, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Kim, E.H.; DellaPenna, D.; Brutnell, T.P. Novel lycopene-ε-cyclase activities in maize revealed through perturbation of carotenoid biosynthesis. Plant J. 2009, 59, 588–599. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.C.; Pizarro, L.; Fuentes, P.; Handford, M.; Cifuentes, V.; Stange, C.; Moreno, J.C.; Fuentes, P. Levels of lycopene-β-cyclase 1 modulate carotenoid gene expression and accumulation in daucus carota. PLoS One 2013, 8, e58144. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Musetti, V.; Kim, J.; Magallanes-Lundback, M.; DellaPenna, D. The Arabidopsis LUT1 locus encodes a member of the cytochrome P450 family that is required for carotenoid ε-ring hydroxylation activity. Proc. Natl. Acad. Sci. USA 2004, 101, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.X.; Pogson, B.; Sun, Z.R.; McDonald, K.A.; DellaPenna, D.; Gantt, E. Functional analysis of the β and ε lycopene cyclase enzymes of Arabidopsis reveals a mechanism for control of cyclic carotenoid formation. Plant Cell 1996, 8, 1613–1626. [Google Scholar] [PubMed]
- Pogson, B.; McDonald, K.A.; Truong, M.; Britton, G.; DellaPenna, D. Arabidopsis carotenoid mutants demonstrate that lutein is not essential for photosynthesis in higher plants. Plant Cell 1996, 8, 1627–1639. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; DellaPenna, D. Defining the primary route for lutein synthesis in plants: The role of Arabidopsis carotenoid β-ring hydroxylase CYP97A3. Proc. Natl. Acad. Sci. USA 2006, 103, 3474–3479. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Kreunen, S.S.; Cuttriss, A.J.; DellaPenna, D.; Pogson, B.J. Identification of the carotenoid isomerase provides insight into carotenoid biosynthesis, prolamellar body formation and photomorphogenesis. Plant Cell 2002, 14, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Cazzonelli, C.I.; Cuttriss, A.J.; Cossetto, S.B.; Pye, W.; Crisp, P.; Whelan, J.; Finnegan, E.J.; Turnbull, C.; Pogson, B.J. Regulation of carotenoid composition and shoot branching in Arabidopsis by a chromatin modifying histone methyltransferase, SDG8. Plant Cell 2009, 21, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Harjes, C.E.; Rocheford, T.R.; Bai, L.; Brutnell, T.P.; Kandianis, C.B.; Sowinski, S.G.; Stapleton, A.E.; Vallabhaneni, R.; Williams, M.; Wurtzel, E.T.; et al. Natural genetic variation in lycopene-ε-cyclase tapped for maize biofortification. Science 2008, 319, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Pogson, B.J.; Rissler, H.M. Genetic manipulation of carotenoid biosynthesis and photoprotection. Philos. Trans. R. Soc. Lond. B 2000, 355, 1395–1403. [Google Scholar] [CrossRef]
- Kim, S.H.; Kima, Y.H.; Ahna, Y.O.; Ahnb, M.J.; Jeonga, J.C.; Lee, H.S.; Kwak, S.S. Down-regulation of the lycopene-ε-cyclase gene increases carotenoid synthesis via the β-branch-specific pathway and enhances salt-stress tolerance in sweetpotato transgenic calli. Physiol. Plantarum 2012, 147, 432–442. [Google Scholar] [CrossRef]
- Diretto, G.; Tavazza, R.; Welsch, R.; Pizzichini, D.; Mourgues, F.; Papacchioli, V.; Beyer, P.; Giuliano, G.; et al. Metabolic engineering of potato tuber carotenoids through tuber-specific silencing of lycopene-ε-cyclase. BMC Plant Biol. 2006, 6, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Schiff, M.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Martin-Hernandez, A.M.; Peart, J.R.; Malcuit, I.; Baulcombe, D.C. Virus-induced gene silencing in plants. Methods 2003, 30, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Faivre-Rampant, O.; Gilroy, E.M.; Hrubikova, K.; Hein, I.; Millam, S.; Loake, G.J.; Birch, P.; Taylor, M.; Lacomme, C. Potato virus X-induced gene silencing in leaves and tubers of potato. Plant Physiol. 2004, 134, 1308–1316. [Google Scholar]
- Jean-Baptiste, H.; Eva-Mari, A.; Kirsi, L. Dynamics of the VIGS-mediated chimeric silencing of the nicotiana benthamiana ChlH gene and of the tobacco mosaic virus vector. MPMI 2003, 16, 99–106. [Google Scholar]
- Ratcliff, F.; Martin-Hernandez, A.M.; Baulcombe, D.C. Tobacco rattle virus as a vector for analysis of gene function by silencing. Plant J. 2001, 25, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.G.; Guo, W.L.; Yin, Y.X.; Gong, Z.H. Novel F-box protein CaF-box is involved in responses to plant hormones and abiotic stress in pepper (Capsicum annuum L.). Int. J. Mol. Sci. 2014, 15, 2413–2430. [Google Scholar]
- Macfarlane, S. Tobraviruses-plant pathogens and tools for biotechnology. Mol. Plant Pathol. 2010, 11, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Zhu, Y.; Li, Q.; Liu, J.; Tian, Y.; Wu, J.; Liu, Y. Development of agrobacterium-mediated virus-induced gene silencing and performance evaluation of four marker genes in gossypium barbadense. PLoS One 2013, 8, e73211. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.Y.; Matyasek, R.; Kovarik, A.; Leitch, A.R. Genome evolution in allotetraploid Nicotiana. Biol. J. Linn. Soc. 2004, 82, 599–606. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P. Tobacco Rar1, EDS1 and NPR1/NIM1 like genes are required for N-mediated resistance to tobacco mosaic virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Kapila, J.; DeRycke, R.; van Montagu, M.; Angenon, G. An agrobacterium-mediated transient gene expression system for intact leaves. Plant Sci. 1997, 122, 101–108. [Google Scholar] [CrossRef]
- Cuningham, F.X. Regulation of carotenoid synthesis and accumulation in plants. Pure Appl. Chem. 2002, 74, 1409–1417. [Google Scholar]
- Hirschberg, J. Carotenoid biosynthesis in flowering plants. Curr. Opin. Plant Biol. 2001, 4, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Farré, G.; Bai, C.; Twyman, R.M.; Capell, T.; Christou, P.; Zhu, C. Nutritious crops producing multiple carotenoids—A metabolic balancing act. Trends Plant Sci. 2011, 16, 532–540. [Google Scholar]
- Chen, X.Y.; Han, H.P.; Jiang, P.; Nie, L.L.; Bao, H.X.; Fan, P.X.; Lv, S.; Feng, J.; Li, Y. Transformation of β-lycopene cyclase genes from Salicornia europaea and Arabidopsis conferred salt tolerance in Arabidopsis and tobacco. Plant Cell Physiol. 2011, 52, 909–921. [Google Scholar]
- Dall’Osto, L.; Fiore, A.; Cazzaniga, S.; Giuliano, G.; Bassi, R. Different roles of α- and β-branch xanthophylls in photosystem assembly and photoprotection. J. Biol. Chem. 2007, 282, 35056–35068. [Google Scholar]
- Dall’Osto, L.; Lico, C.; Alric, J.; Giuliano, G.; Havaux, M.; Bassi, R. Lutein is needed for efficient chlorophyll triplet quenching in the major LHCII antenna complex of higher plants and effective photoprotection in vivo under strong light. BMC Plant Biol. 2006, 6, 32–51. [Google Scholar]
- Kalituho, L.; Rech, J.; Jahns, P. The roles of specific xanthophylls in light utilization. Planta 2007, 225, 423–439. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, J.; Zhao, X.Q.; Wang, J.; Wong, G.K.; Yu, J. KaKs_Calculator: Calculating Ka and Ks through model selection and model averaging. Genomics Proteomics Bioinform. 2006, 4, 259–263. [Google Scholar] [CrossRef]
- Sanderson, M.J. Molecular Systematics of Plants II: DNA Sequencing; Springer US: London, UK, 1998; pp. 242–264. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shi, Y.; Wang, R.; Luo, Z.; Jin, L.; Liu, P.; Chen, Q.; Li, Z.; Li, F.; Wei, C.; Wu, M.; et al. Molecular Cloning and Functional Characterization of the Lycopene ?-Cyclase Gene via Virus-Induced Gene Silencing and Its Expression Pattern in Nicotiana tabacum. Int. J. Mol. Sci. 2014, 15, 14766-14785. https://doi.org/10.3390/ijms150814766
Shi Y, Wang R, Luo Z, Jin L, Liu P, Chen Q, Li Z, Li F, Wei C, Wu M, et al. Molecular Cloning and Functional Characterization of the Lycopene ?-Cyclase Gene via Virus-Induced Gene Silencing and Its Expression Pattern in Nicotiana tabacum. International Journal of Molecular Sciences. 2014; 15(8):14766-14785. https://doi.org/10.3390/ijms150814766
Chicago/Turabian StyleShi, Yanmei, Ran Wang, Zhaopeng Luo, Lifeng Jin, Pingping Liu, Qiansi Chen, Zefeng Li, Feng Li, Chunyang Wei, Mingzhu Wu, and et al. 2014. "Molecular Cloning and Functional Characterization of the Lycopene ?-Cyclase Gene via Virus-Induced Gene Silencing and Its Expression Pattern in Nicotiana tabacum" International Journal of Molecular Sciences 15, no. 8: 14766-14785. https://doi.org/10.3390/ijms150814766
APA StyleShi, Y., Wang, R., Luo, Z., Jin, L., Liu, P., Chen, Q., Li, Z., Li, F., Wei, C., Wu, M., Wei, P., Xie, H., Qu, L., Lin, F., & Yang, J. (2014). Molecular Cloning and Functional Characterization of the Lycopene ?-Cyclase Gene via Virus-Induced Gene Silencing and Its Expression Pattern in Nicotiana tabacum. International Journal of Molecular Sciences, 15(8), 14766-14785. https://doi.org/10.3390/ijms150814766