Maternal High-Fat Diet Modulates Hepatic Glucose, Lipid Homeostasis and Gene Expression in the PPAR Pathway in the Early Life of Offspring


Abstract
:1. Introduction
2. Results
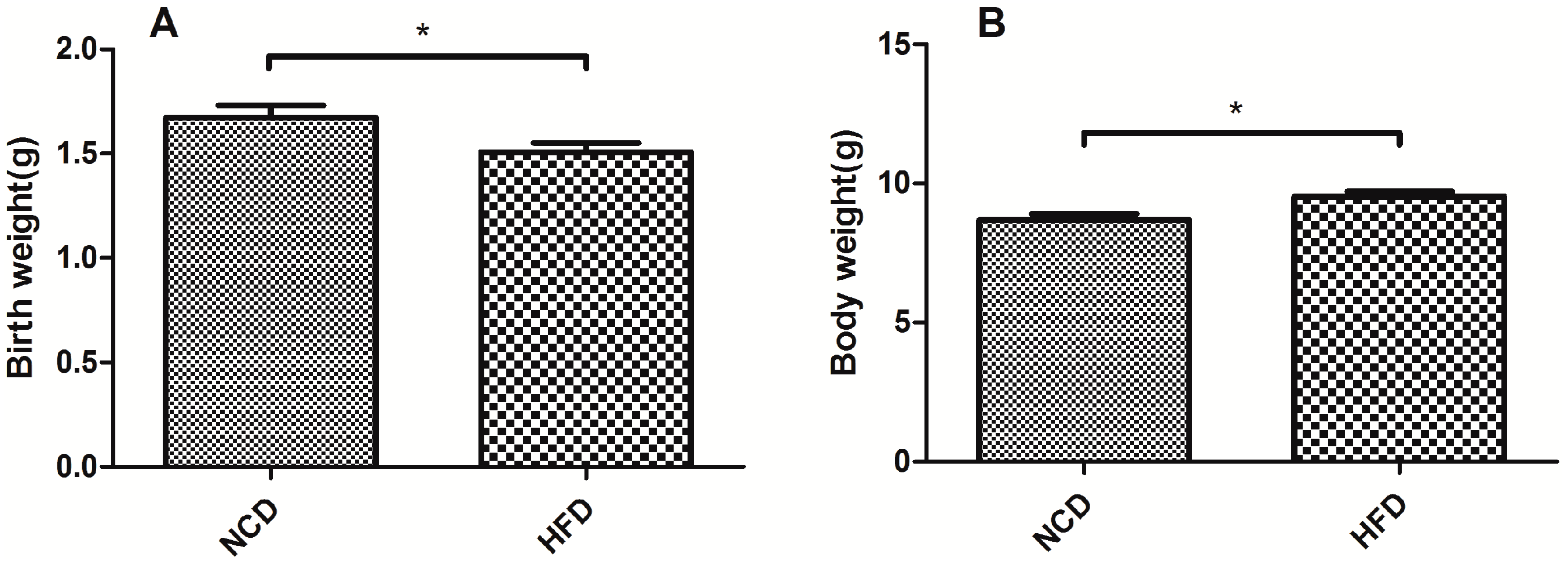
2.1. Offspring of the Dams Fed a High-Fat Diet Had a Lower Birth Weight and a Heavier Body Weight at Weaning

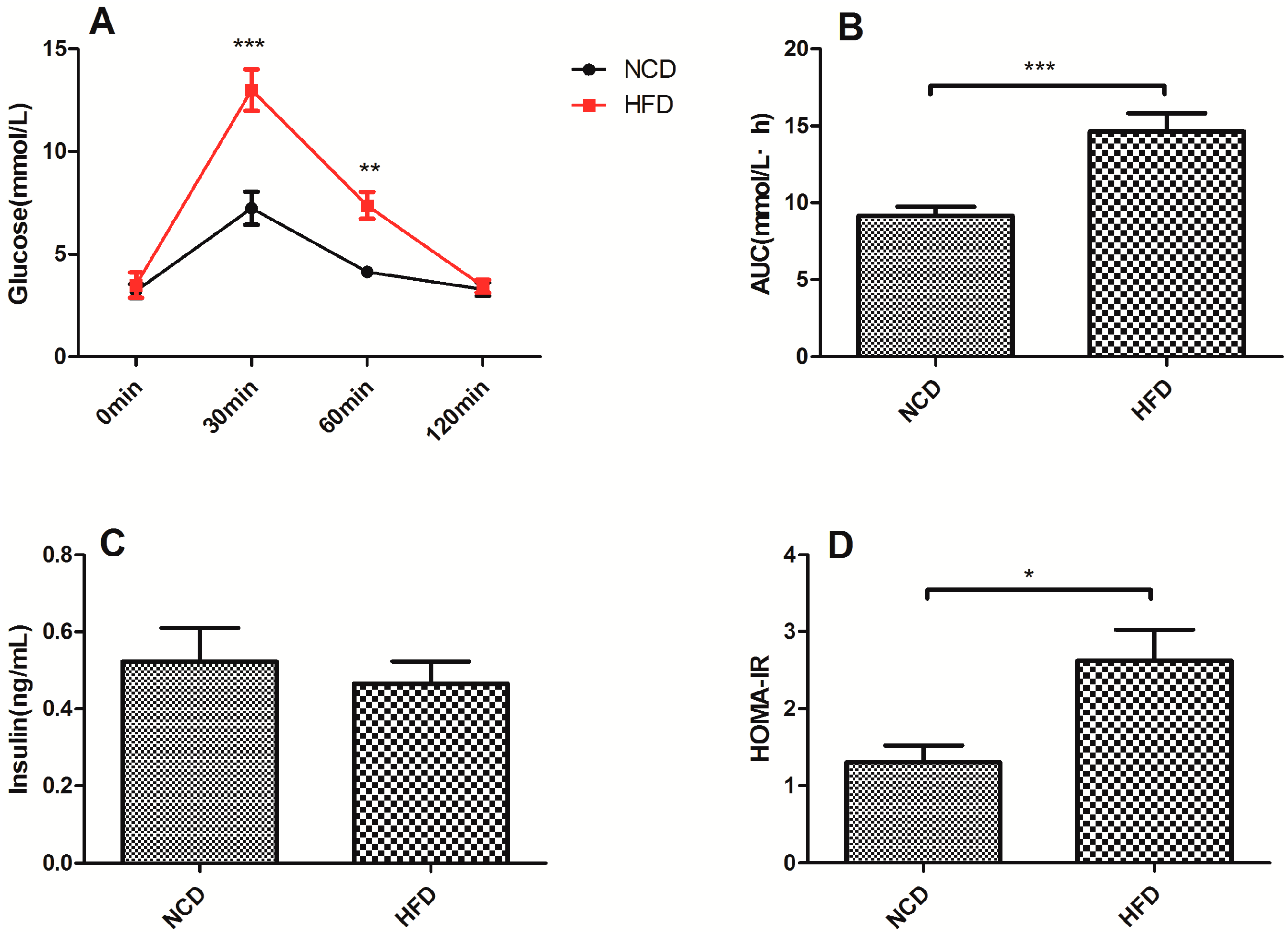
2.2. Offspring of the Dams Fed a High-Fat Diet Exhibited Impaired Glucose Tolerance and Decreased Insulin Sensitivity
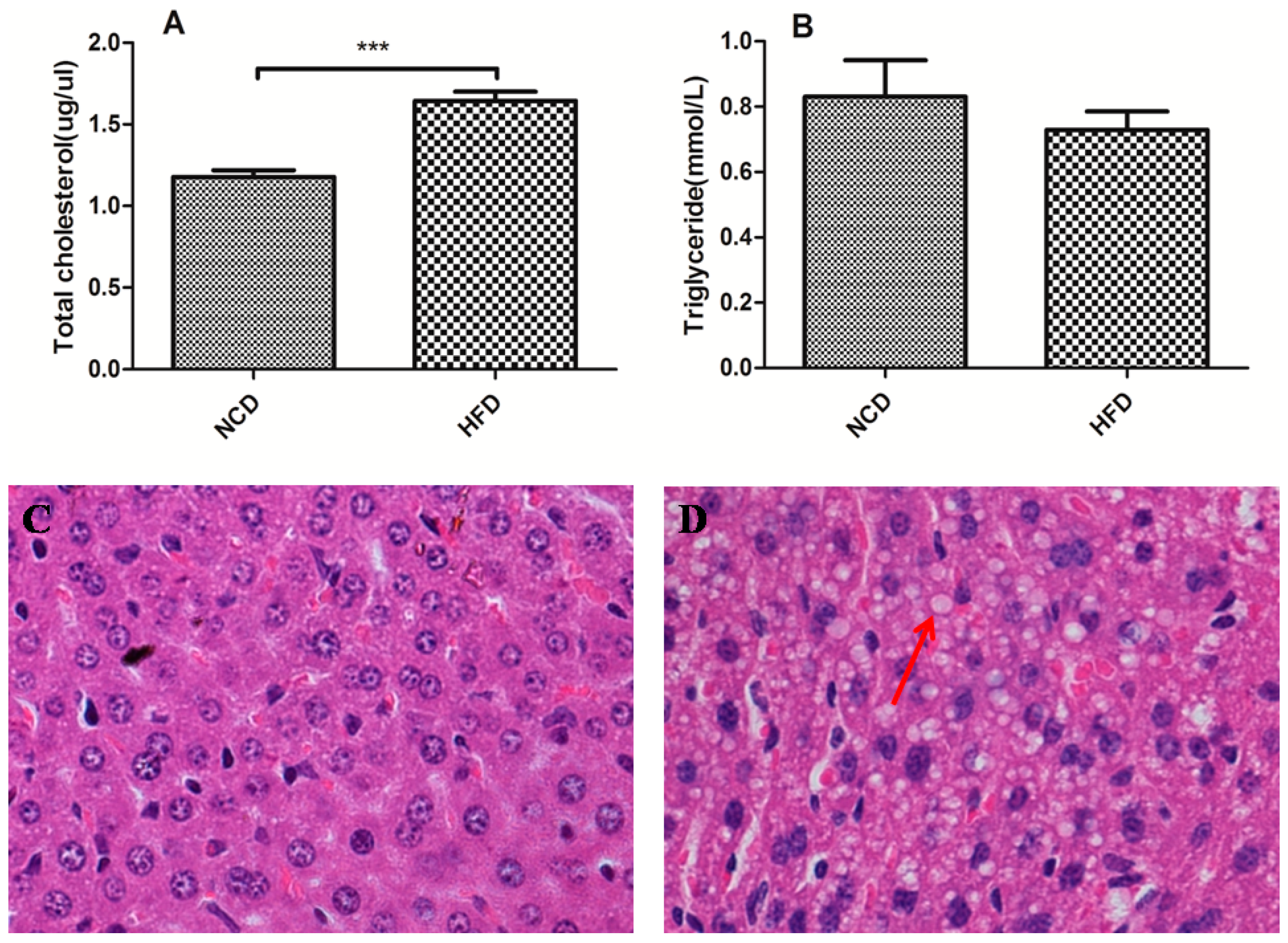
2.3. Offspring of the Dams Fed a High-Fat Diet Modulated Lipid Homeostasis


2.4. The Comparison of Biochemical Parameters of the Male and Female Offspring at Weaning

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biochemical Parameters | Male | Female | ||||
|---|---|---|---|---|---|---|
| NCD | HFD | p Value | NCD | HFD | p Value | |
| FBG (mmol/L) | 3.89 ± 1.56 | 5.19 ± 3.17 | 0.21 | 3.54 ± 1.72 | 4.89 ± 2.96 | 0.18 |
| Body weight (g) | 8.74 ± 1.14 | 9.73 ± 0.79 | 0.038 * | 8.79 ± 1.42 | 9.33 ± 0.53 | 0.32 |
| AUC of IPGTT (mmol/L·h) | 8.08 ± 1.49 | 16.45 ± 5.95 | 0.0016 ** | 10.24 ± 2.69 | 12.97 ± 4.03 | 0.12 |
| Serum insulin (µU/mL) | 0.56 ± 0.37 | 0.37 ± 0.20 | 0.55 | 0.69 ± 0.19 | 0.51 ± 0.25 | 0.35 |
| Triglyceride (mmol/L) | 1.11 ± 0.43 | 0.83 ± 0.19 | 0.15 | 1.10 ± 0.71 | 0.62 ± 0.18 | 0.11 |
| Total cholesterol (µg/uL) | 1.24 ± 0.12 | 1.77 ± 0.21 | <0.0001 *** | 1.12 ± 0.17 | 1.22 ± 0.13 | 0.054 |
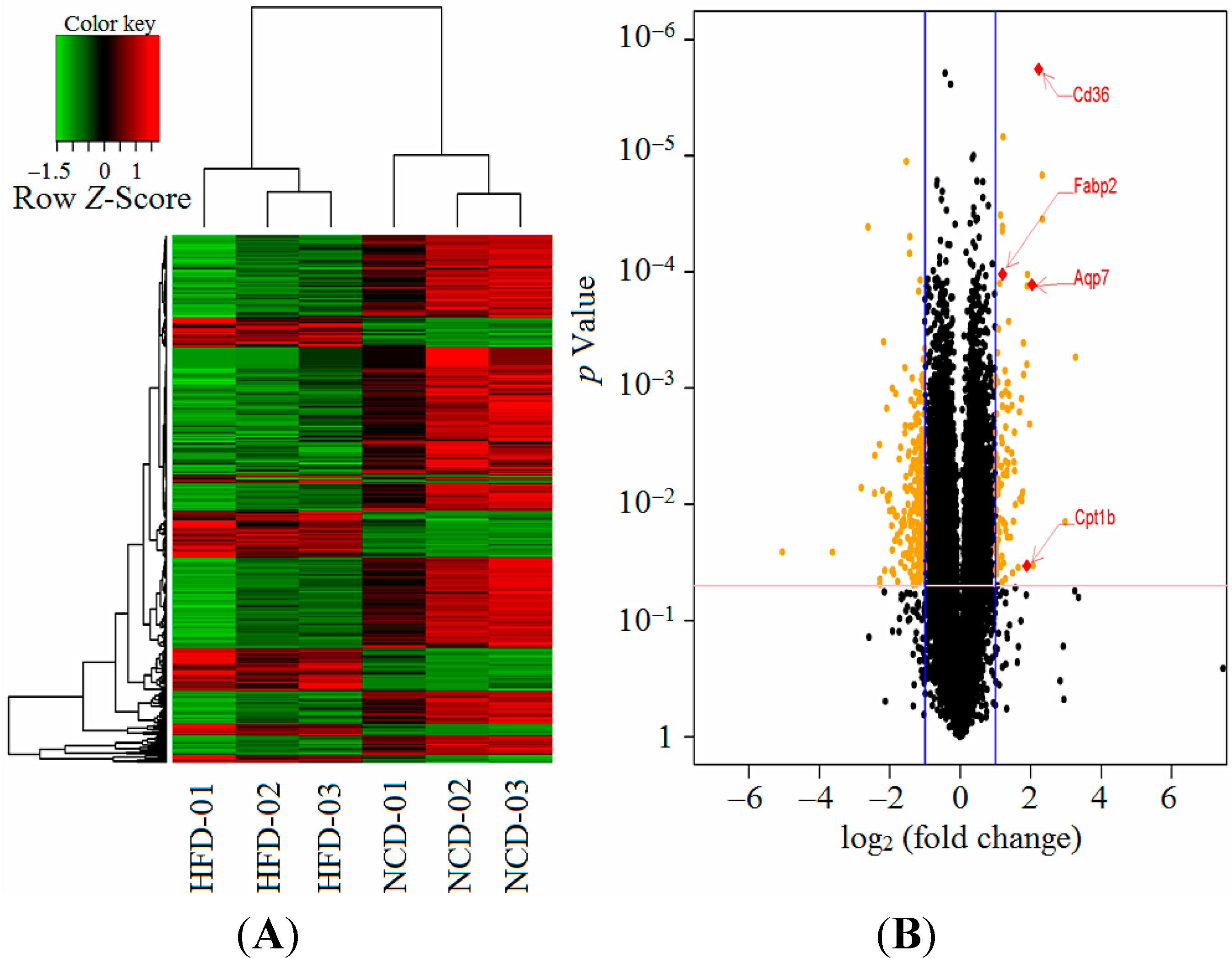
2.5. Maternal High-Fat Diet Regulated Gene Expression in the Male Offspring

2.6. Hepatic Gene Expression for the Validation of Array Analysis
| GO Classification | GO Term | GO ID | C | O | E | R | Rawp | Adjp |
|---|---|---|---|---|---|---|---|---|
| Biological process | Response to pheromone | GO:0019236 | 101 | 9 | 1.3 | 6.93 | 6.37 ×10−6 | 0.0075 |
| Biological process | G-protein coupled receptor signaling pathway | GO:0007186 | 2041 | 48 | 26.26 | 1.83 | 3.32 ×10−5 | 0.0196 |
| Biological process | Response to chemical stimulus | GO:0042221 | 2194 | 49 | 28.23 | 1.74 | 9.92 ×10−5 | 0.0391 |
| Biological process | Response to organic substance | GO:0010033 | 1538 | 37 | 19.79 | 1.87 | 0.0002 | 0.0394 |
| Biological process | Response to stimulus | GO:0050896 | 7009 | 119 | 90.19 | 1.32 | 0.0002 | 0.0394 |
| Biological process | Very long-chain fatty acid metabolic process | GO:0000038 | 24 | 4 | 0.31 | 12.95 | 0.0002 | 0.0394 |
| Biological process | Regulation of mRNA processing | GO:0050684 | 46 | 5 | 0.59 | 8.45 | 0.0003 | 0.0443 |
| Molecular function | Signal transducer activity | GO:0004871 | 2351 | 56 | 29.57 | 1.89 | 2.10 ×10−6 | 0.0001 |
| Molecular function | Molecular transducer activity | GO:0060089 | 2351 | 56 | 29.57 | 1.89 | 2.10 ×10−6 | 0.0001 |
| Molecular function | Pheromone binding | GO:0005550 | 91 | 9 | 1.14 | 7.86 | 2.22 ×10−6 | 0.0001 |
| Molecular function | Pheromone receptor activity | GO:0016503 | 99 | 9 | 1.25 | 7.23 | 4.49 ×10−6 | 0.0001 |
| Molecular function | Signaling receptor activity | GO:0038023 | 2127 | 52 | 26.76 | 1.94 | 2.47 ×10−6 | 0.0001 |
| Molecular function | Transmembrane signaling receptor activity | GO:0004888 | 2039 | 49 | 25.65 | 1.91 | 8.11 ×10−6 | 0.0002 |
| Molecular function | G-protein coupled receptor activity | GO:0004930 | 1612 | 37 | 20.28 | 1.82 | 0.0003 | 0.0063 |
| Molecular function | Glutathione transferase activity | GO:0004364 | 26 | 4 | 0.33 | 12.23 | 0.0003 | 0.0063 |
| Molecular function | Palmitoyl-CoA hydrolase activity | GO:0016290 | 15 | 3 | 0.19 | 15.9 | 0.0008 | 0.0153 |
| Molecular function | Acyl-CoA hydrolase activity | GO:0047617 | 19 | 3 | 0.24 | 12.55 | 0.0016 | 0.0283 |
| Molecular function | CoA hydrolase activity | GO:0016289 | 22 | 3 | 0.28 | 10.84 | 0.0025 | 0.0411 |
| Molecular function | Carboxylesterase activity | GO:0004091 | 48 | 4 | 0.6 | 6.62 | 0.0031 | 0.0475 |
| KEGG ID | Pathway Name | Gene Symbol | C | O | E | R | Rawp | Adjp |
|---|---|---|---|---|---|---|---|---|
| 04110 | Cell cycle | Cdkn1a, Ccnb1, Mad2l1, Ccnb2, Gm5593, Wee1, LOC100045924 | 127 | 7 | 0.8 | 8.74 | 2 × 10−5 | 0.0003 |
| 01040 | Biosynthesis of unsaturated fatty acids | Acot4, Acot3, Acot2 | 25 | 3 | 0.16 | 19.02 | 0.0005 | 0.0044 |
| 03320 | PPAR signaling pathway | Aqp7, Fabp2, Cpt1b, Cd36 | 80 | 4 | 0.5 | 7.93 | 0.0017 | 0.0125 |
| 04060 | Cytokine-cytokine receptor interaction | Il6ra, Il1r1, Il1r1, Cxcl13, Ccl20, Il22ra1 | 245 | 6 | 1.55 | 3.88 | 0.0049 | 0.0232 |
| 00480 | Glutathione metabolism | Gsta2, Gsta1, Mgst3 | 54 | 3 | 0.34 | 8.81 | 0.0049 | 0.0232 |
| 00562 | Inositol phosphate metabolism | Pten, Pik3c2g, Inpp4a | 57 | 3 | 0.36 | 8.34 | 0.0057 | 0.0232 |
| 04920 | Adipocytokine signaling pathway | Cpt1b, Ppargc1a, Cd36 | 68 | 3 | 0.43 | 6.99 | 0.0092 | 0.0311 |
| 04070 | Phosphatidylinositol signaling system | Pten, Pik3c2g, Inpp4a | 78 | 3 | 0.49 | 6.1 | 0.0134 | 0.0369 |
| Gene Symbol | Fold Change (Gene Array) | p Value (Gene Array) | Fold Change (qRT-PCR) | p Value (qRT-PCR) |
|---|---|---|---|---|
| Cd36 | 4.674 | 1.8048 × 10−6 | 4.9 ± 0.12 | <0.0001 |
| Aqp7 | 4.104 | 0.0001288 | 3.8 ± 0.27 | 0.0044 |
| Cpt1b | 3.705 | 0.03375443 | 3.7 ± 0.05 | 0.0292 |
| Fabp2 | 2.302 | 0.00010463 | 2.5 ± 0.18 | 0.0002 |
| PPARα | 1.318 | 0.04499 | 1.32 ± 0.21 | 0.038 |
| PPARγ | 1.562 | 0.0041 | 1.61 ± 0.18 | 0.012 |
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Protocol
4.2. Measurement of Body Weight
4.3. Glucose Tolerance Tests
4.4. Measurement of Serum Insulin, Triacylglycerol and Total Cholesterol Levels
4.5. Liver Histological Analysis
4.6. RNA Preparation and Gene Array Experiments
4.7. Gene Array Data Analysis
4.8. Quantitative RT-PCR Experiment
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Skogen, J.C.; Overland, S. The fetal origins of adult disease: A narrative review of the epidemiological literature. JRSM Short Rep. 2012, 3, 59. [Google Scholar] [CrossRef]
- Painter, R.C.; Roseboom, T.J.; Bleker, O.P. Prenatal exposure to the Dutch famine and disease in later life: An overview. Reprod. Toxicol. 2005, 20, 345–352. [Google Scholar] [CrossRef]
- Elahi, M.M.; Cagampang, F.R.; Mukhtar, D.; Anthony, F.W.; Ohri, S.K.; Hanson, M.A. Long-term maternal high-fat feeding from weaning through pregnancy and lactation predisposes offspring to hypertension, raised plasma lipids and fatty liver in mice. Br. J. Nutr. 2009, 102, 514–519. [Google Scholar] [CrossRef]
- Curhan, G.C.; Willett, W.C.; Rimm, E.B.; Spiegelman, D.; Ascherio, A.L.; Stampfer, M.J. Birth weight and adult hypertension, diabetes mellitus, and obesity in US men. Circulation 1996, 94, 3246–3250. [Google Scholar] [CrossRef]
- Carone, B.R.; Fauquier, L.; Habib, N.; Shea, J.M.; Hart, C.E.; Li, R.; Bock, C.; Li, C.; Gu, H.; Zamore, P.D. Paternally induced transgenerational environmental reprogramming of metabolic gene expression in mammals. Cell 2010, 143, 1084–1096. [Google Scholar] [CrossRef]
- Jousse, C.; Parry, L.; Lambert-Langlais, S.; Maurin, A.C.; Averous, J.; Bruhat, A.; Carraro, V.; Tost, J.; Letteron, P.; Chen, P. Perinatal undernutrition affects the methylation and expression of the leptin gene in adults: Implication for the understanding of metabolic syndrome. FASEB J. 2011, 25, 3271–3278. [Google Scholar] [CrossRef]
- Guariguata, L.; Linnenkamp, U.; Beagley, J.; Whiting, D.R.; Cho, N.H. Global estimates of the prevalence of hyperglycaemia in pregnancy. Diabetes Res. Clin. Pract. 2014, 103, 176–185. [Google Scholar] [CrossRef]
- Pentinat, T.; Ramon-Krauel, M.; Cebria, J.; Diaz, R.; Jimenez-Chillaron, J.C. Transgenerational inheritance of glucose intolerance in a mouse model of neonatal overnutrition. Endocrinology 2010, 151, 5617–5623. [Google Scholar] [CrossRef]
- Bruce, K.D.; Cagampang, F.R.; Argenton, M.; Zhang, J.; Ethirajan, P.L.; Burdge, G.C.; Bateman, A.C.; Clough, G.F.; Poston, L.; Hanson, M.A. Maternal high-fat feeding primes steatohepatitis in adult mice offspring, involving mitochondrial dysfunction and altered lipogenesis gene expression. Hepatology 2009, 50, 1796–1808. [Google Scholar] [CrossRef]
- Miotto, P.M.; Castelli, L.M.; Amoye, F.; LeBlanc, P.J.; Peters, S.J.; Roy, B.D.; Ward, W.E. Maternal high fat feeding does not have long-lasting effects on body composition and bone health in female and male Wistar rat offspring at young adulthood. Molecules 2013, 18, 15094–15109. [Google Scholar] [CrossRef]
- McKay, J.A.; Xie, L.; Manus, C.; Langie, S.A.; Maxwell, R.J.; Ford, D.; Mathers, J.C. Metabolic effects of a high-fat diet post-weaning after low maternal dietary folate during pregnancy and lactation. Mol. Nutr. Food Res. 2014, 5, 1087–1097. [Google Scholar]
- Langley-Evans, S.C. Intrauterine programming of hypertension in the rat: Nutrient interactions. Comp. Biochem. Physiol. A 1996, 114, 327–333. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, F.; Didelot, X.; Bruce, K.D.; Cagampang, F.R.; Vatish, M.; Hanson, M.; Lehnert, H.; Ceriello, A.; Byrne, C.D. Maternal high fat diet during pregnancy and lactation alters hepatic expression of insulin like growth factor-2 and key microRNAs in the adult offspring. BMC Genomics 2009, 10, 478. [Google Scholar] [CrossRef]
- Yokomizo, H.; Inoguchi, T.; Sonoda, N.; Sakaki, Y.; Maeda, Y.; Inoue, T.; Hirata, E.; Takei, R.; Ikeda, N.; Fujii, M. Maternal high fat diet induces insulin resistance and deterioration of pancreatic beta cell function in adult offspring with gender differences in mice. Am. J. Physiol. 2014, 10, E1163–E1175. [Google Scholar]
- Ong, K.K.; Petry, C.J.; Emmett, P.M.; Sandhu, M.S.; Kiess, W.; Hales, C.N.; Ness, A.R.; Dunger, D.B. Insulin sensitivity and secretion in normal children related to size at birth, postnatal growth, and plasma insulin-like growth factor-I levels. Diabetologia 2004, 47, 1064–1070. [Google Scholar]
- Dulloo, A.G. A role for suppressed skeletal muscle thermogenesis in pathways from weight fluctuations to the insulin resistance syndrome. Acta Physiol. Scand. 2005, 184, 295–307. [Google Scholar] [CrossRef]
- Crescenzo, R.; Samec, S.; Antic, V.; Rohner-Jeanrenaud, F.; Seydoux, J.; Montani, J.P.; Dulloo, A.G. A role for suppressed thermogenesis favoring catch-up fat in the pathophysiology of catch-up growth. Diabetes 2003, 52, 1090–1097. [Google Scholar] [CrossRef]
- Purcell, R.H.; Sun, B.; Pass, L.L.; Power, M.L.; Moran, T.H.; Tamashiro, K.L. Maternal stress and high-fat diet effect on maternal behavior, milk composition, and pup ingestive behavior. Physiol. Behav. 2011, 104, 474–479. [Google Scholar] [CrossRef]
- Sun, B.; Purcell, R.H.; Terrillion, C.E.; Yan, J.; Moran, T.H.; Tamashiro, K.L. Maternal high-fat diet during gestation or suckling differentially affects offspring leptin sensitivity and obesity. Diabetes 2012, 61, 2833–2841. [Google Scholar] [CrossRef]
- Guo, F.; Jen, K.L. High-fat feeding during pregnancy and lactation affects offspring metabolism in rats. Physiol. Behav. 1995, 57, 681–686. [Google Scholar] [CrossRef]
- Chen, S.H.; He, F.; Zhou, H.L.; Wu, H.R.; Xia, C.; Li, Y.M. Relationship between nonalcoholic fatty liver disease and metabolic syndrome. J. Dig. Dis. 2011, 12, 125–130. [Google Scholar] [CrossRef]
- Yamaguchi, R.; Nakagawa, Y.; Liu, Y.J.; Fujisawa, Y.; Sai, S.; Nagata, E.; Sano, S.; Satake, E.; Matsushita, R.; Nakanishi, T. Effects of maternal high-fat diet on serum lipid concentration and expression of peroxisomal proliferator-activated receptors in the early life of rat offspring. Horm. Metab. Res. 2010, 42, 821–825. [Google Scholar] [CrossRef]
- Feige, J.N.; Gelman, L.; Michalik, L.; Desvergne, B.; Wahli, W. From molecular action to physiological outputs: Peroxisome proliferator-activated receptors are nuclear receptors at the crossroads of key cellular functions. Prog. Lipid Res. 2006, 45, 120–159. [Google Scholar] [CrossRef]
- Christodoulides, C.; Vidal-Puig, A. PPARs and adipocyte function. Mol. Cell. Endocrinol. 2010, 318, 61–68. [Google Scholar] [CrossRef]
- Grygiel-Gorniak, B. Peroxisome proliferator-activated receptors and their ligands: Nutritional and clinical implications—A review. Nutr. J. 2014, 13, 17. [Google Scholar] [CrossRef]
- Hyatt, M.A.; Gardner, D.S.; Sebert, S.; Wilson, V.; Davidson, N.; Nigmatullina, Y.; Chan, L.L.; Budge, H.; Symonds, M.E. Suboptimal maternal nutrition, during early fetal liver development, promotes lipid accumulation in the liver of obese offspring. Reproduction 2011, 141, 119–126. [Google Scholar] [CrossRef]
- Erhuma, A.; Salter, A.M.; Sculley, D.V.; Langley-Evans, S.C.; Bennett, A.J. Prenatal exposure to a low-protein diet programs disordered regulation of lipid metabolism in the aging rat. Am. J. Physiol. 2007, 292, E1702–E1714. [Google Scholar]
- Kalanderian, A.; Abate, N.; Patrikeev, I.; Wei, J.; Vincent, K.L.; Motamedi, M.; Saade, G.R.; Bytautiene, E. Pioglitazone therapy in mouse offspring exposed to maternal obesity. Am. J. Obstet. Gynecol. 2013, 208, 308. [Google Scholar]
- Magliano, D.C.; Bargut, T.C.; de Carvalho, S.N.; Aguila, M.B.; Mandarim-de-Lacerda, C.A.; Souza-Mello, V. Peroxisome proliferator-activated receptors-α and γ are targets to treat offspring from maternal diet-induced obesity in mice. PLoS One 2013, 8, e64258. [Google Scholar] [CrossRef]
- Fruchart, J.C.; Staels, B.; Duriez, P. The role of fibric acids in atherosclerosis. Curr. Atheroscler. Rep. 2001, 3, 83–92. [Google Scholar] [CrossRef]
- Krishnaswami, A.; Ravi-Kumar, S.; Lewis, J.M. Thiazolidinediones: A 2010 perspective. Perm. J. 2010, 14, 64–72. [Google Scholar]
- Pershadsingh, H.A. Dual peroxisome proliferator-activated receptor-α/γ agonists: In the treatment of type 2 diabetes mellitus and the metabolic syndrome. Treat. Endocrinol. 2006, 5, 89–99. [Google Scholar] [CrossRef]
- Winzell, M.S.; Ahren, B. The high-fat diet-fed mouse: A model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes 2004, 53, S215–S219. [Google Scholar] [CrossRef]
- Principles of Laboratory Animal Care, National Institutes of Health Publication No. 85–23. Available online: http://grants1.nih.gov/grants/olaw/references/phspol.htm (accessed on 20 May 2014).
- Purves, R.D. Optimum numerical integration methods for estimation of area-under-the-curve (AUC) and area-under-the-moment-curve (AUMC). J. Pharm. Biopharm. 1992, 20, 211–226. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- WEB-Based GEne SeT AnaLysis Toolkit. Available online: http://bioinfo.vanderbilt.edu/webgestalt/ (accessed on 20 May 2014).
- Zhang, Q.; Xiao, X.; Li, M.; Li, W.; Yu, M.; Zhang, H.; Sun, X.; Mao, L.; Xiang, H. Telmisartan improves kidney function through inhibition of the oxidative phosphorylation pathway in diabetic rats. J. Mol. Endocrinol. 2012, 49, 35–46. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zheng, J.; Xiao, X.; Zhang, Q.; Yu, M.; Xu, J.; Wang, Z. Maternal High-Fat Diet Modulates Hepatic Glucose, Lipid Homeostasis and Gene Expression in the PPAR Pathway in the Early Life of Offspring. Int. J. Mol. Sci. 2014, 15, 14967-14983. https://doi.org/10.3390/ijms150914967
Zheng J, Xiao X, Zhang Q, Yu M, Xu J, Wang Z. Maternal High-Fat Diet Modulates Hepatic Glucose, Lipid Homeostasis and Gene Expression in the PPAR Pathway in the Early Life of Offspring. International Journal of Molecular Sciences. 2014; 15(9):14967-14983. https://doi.org/10.3390/ijms150914967
Chicago/Turabian StyleZheng, Jia, Xinhua Xiao, Qian Zhang, Miao Yu, Jianping Xu, and Zhixin Wang. 2014. "Maternal High-Fat Diet Modulates Hepatic Glucose, Lipid Homeostasis and Gene Expression in the PPAR Pathway in the Early Life of Offspring" International Journal of Molecular Sciences 15, no. 9: 14967-14983. https://doi.org/10.3390/ijms150914967
APA StyleZheng, J., Xiao, X., Zhang, Q., Yu, M., Xu, J., & Wang, Z. (2014). Maternal High-Fat Diet Modulates Hepatic Glucose, Lipid Homeostasis and Gene Expression in the PPAR Pathway in the Early Life of Offspring. International Journal of Molecular Sciences, 15(9), 14967-14983. https://doi.org/10.3390/ijms150914967