Recent Progress on Liver Kinase B1 (LKB1): Expression, Regulation, Downstream Signaling and Cancer Suppressive Function



Abstract
:1. Introduction

{kind=link}
| Model of Knockout Mice | Main Abnormalities | References |
|---|---|---|
| LKB1−/− mice | Death at midgestation, with neural tube defects, mesenchymal cell death, and vascular abnormalities | [15] |
| LKB1−/− mice | Death in utero between 8.5 and 9.5 days postcoitum, with developmental retardation of embryo | [16] |
| LKB1+/− mice | Development of multiple gastric adenomatous polyps | [16] |
| LKB1+/− mice | Development of hamartomatous polyps in gastrointestinal tract | [17] |
| LKB1+/− mice | Development of hepatocellular carcinomas | [18] |
| LKB1+/− mice | Development of severe gastrointestinal polyposis | [19] |
| Muscle-specific LKB1−/− mice | Enhancement of insulin sensitivity in muscles | [20] |
| Endothelium-specific LKB1−/− mice | Embryonic death at E12.5, with a loss of vascular smooth muscle cells and vascular disruption | [21] |
| Embryo fibroblasts LKB1−/− mice | Defect of myofibroblast differentiation | [22] |
| LKB1(s) −/− mice | Male infertility, with abnormality at spermiation | [23] |
| LKB1+/− mice | Development of osteogenic tumours | [24] |
| Prostate-specific LKB1−/− mice | Development of prostate neoplasia | [25] |
| LKB1(s) −/− mice | Male infertility, with abnormal spermiation | [26] |
| Mullerian duct mesenchyme-derived cell-specific LKB1−/− mice | Development of oviductal adenomas and endometrial cancer | [27] |
| Somatic testicular cells-specific LKB1−/− mice | Male sub-infertility, with focal vacuolization in some of the seminiferous tubules, progressive germ cell loss and Sertoli cell only tubules | [28] |
| Adipose tissue-specific LKB1−/− mice | Reduced amount of white adipose tissue, postnatal growth retardation, and early death before weaning | [29] |
| Muscle-specific LKB1−/− mice | Reduced fatty acid oxidation during treadmill exercise | [30] |
| LKB1−/− mice | Reduce of the latency of ErbB2-mediated tumorigenesis | [31] |
2. Expression Pattern of Liver Kinase B1 (LKB1)
3. Subcellular Distribution of LKB1
4. Binding Proteins of LKB1
5. Regulation of LKB1 Expression
5.1. Epigenetic Modification
5.2. Transcriptional Regulation
5.3. Posttranslational Modification
6. LKB1 Downstream Pathways
6.1. LKB1—A Master Kinase of AMPK-Related Protein Kinases
6.2. LKB1–AMPK Signaling Pathway
6.3. LKB–MARK/Par1 Signaling Pathway
6.4. LKB1–SIK Signaling Pathway
6.5. LKB1–SNRK/NUAK2 Signaling
6.6. LKB1–BRSK/SADK Signaling
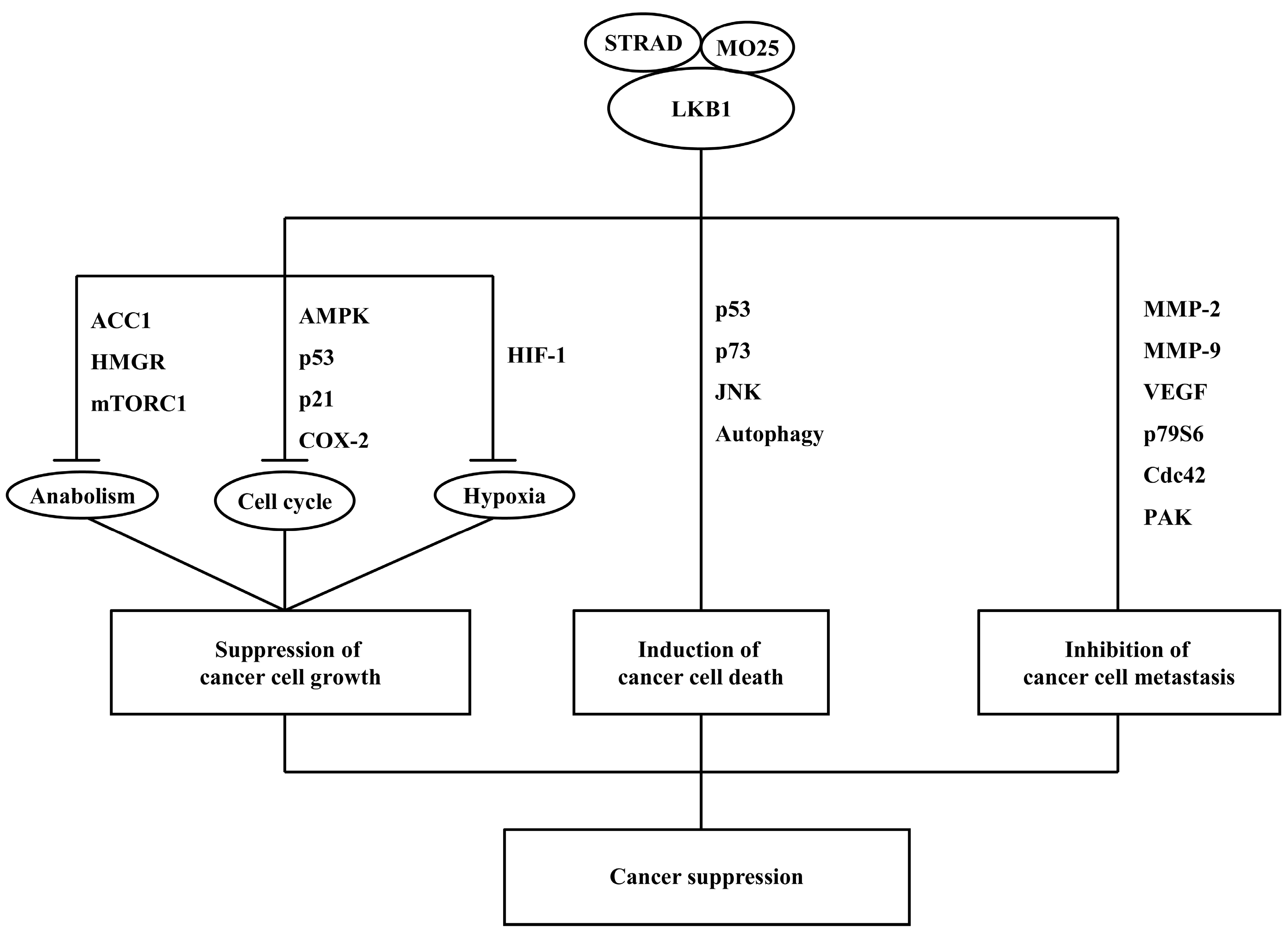
7. LKB1 Functions as a Cancer Suppressor

7.1. Suppression of Cancer Cell Growth
7.2. Induction of Cancer Cell Death
7.3. Inhibition of Cancer Cell Metastasis
7.4. Therapeutic Approaches Targeting LKB1
| Drugs | Target of LKB1 Signaling | Effects on Cancer Cells | References |
|---|---|---|---|
| Celecoxib | Inhibition of COX-2 | Inhibition of polyps in vivo | [117] |
| Metformin | Activation of AMPK | Induction of cancer cell death | [118] |
| Rapamycin | Inhibition of mTOR | Suppression of tumor in vivo | [119] |
| Resveratrol | Activation of AMPK | Inhibition of caner cell growth | [120] |
7.5. LKB1—Still a Cancer Promoter?
8. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hemminki, A.; Markie, D.; Tomlinson, I.; Avizienyte, E.; Roth, S.; Loukola, A.; Bignell, G.; Warren, W.; Aminoff, M.; Hoglund, P.; et al. A serine/threonine kinase gene defective in Peutz-Jeghers syndrome. Nature 1998, 391, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Jenne, D.E.; Reimann, H.; Nezu, J.; Friedel, W.; Loff, S.; Jeschke, R.; Muller, O.; Back, W.; Zimmer, M. Peutz-Jeghers syndrome is caused by mutations in a novel serine threonine kinase. Nat. Genet. 1998, 18, 38–43. [Google Scholar] [CrossRef]
- Avizienyte, E.; Roth, S.; Loukola, A.; Hemminki, A.; Lothe, R.A.; Stenwig, A.E.; Fossa, S.D.; Salovaara, R.; Aaltonen, L.A. Somatic mutations in LKB1 are rare in sporadic colorectal and testicular tumors. Cancer Res. 1998, 58, 2087–2090. [Google Scholar] [PubMed]
- Park, W.S.; Moon, Y.W.; Yang, Y.M.; Kim, Y.S.; Kim, Y.D.; Fuller, B.G.; Vortmeyer, A.O.; Fogt, F.; Lubensky, I.A.; Zhuang, Z.P. Mutations of the STK11 gene in sporadic gastric carcinoma. Int. J. Oncol. 1998, 13, 601–604. [Google Scholar] [PubMed]
- Avizienyte, E.; Loukola, A.; Roth, S.; Hemminki, A.; Tarkkanen, M.; Salovaara, R.; Arola, J.; Butzow, R.; Husgafvel-Pursiainen, K.; Kokkola, A.; et al. LKB1 somatic mutations in sporadic tumors. Am. J. Pathol. 1999, 154, 677–681. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Cespedes, M.; Parrella, P.; Esteller, M.; Nomoto, S.; Trink, B.; Engles, J.M.; Westra, W.H.; Herman, J.G.; Sidransky, D. Inactivation of LKB1/STK11 is a common event in adenocarcinomas of the lung. Cancer Res. 2002, 62, 3659–3662. [Google Scholar] [PubMed]
- Bignell, G.R.; Barfoot, R.; Seal, S.; Collins, N.; Warren, W.; Stratton, M.R. Low frequency of somatic mutations in the LKB1/Peutz-Jeghers syndrome gene in sporadic breast cancer. Cancer Res. 1998, 58, 1384–1386. [Google Scholar] [PubMed]
- Baty, D.; Forster, L.F.; Defres, S.; Goudie, D.; Carey, F.A. Involvement of the Peutz-Jeghers syndrome gene LKB1 in breast and colon cancer. J. Med. Genet. 1999, 36, S46–S46. [Google Scholar]
- Hruban, R.H.; Petersen, G.M.; Goggins, M.; Tersmette, A.C.; Offerhaus, G.J.A.; Falatko, F.; Yeo, C.J.; Kern, S.E. Familial pancreatic cancer. Ann. Oncol. 1999, 10, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Su, G.H.; Hruban, R.H.; Bansal, R.K.; Bova, G.S.; Tang, D.J.; Shekher, M.C.; Westerman, A.M.; Entius, M.M.; Goggins, M.; Yeo, C.J.; et al. Germline and somatic mutations of the STK11/LKB1 Peutz-Jeghers gene in pancreatic and biliary cancers. Am. J. Pathol. 1999, 154, 1835–1840. [Google Scholar] [CrossRef]
- Guldberg, P.; Straten, P.T.; Ahrenkiel, V.; Seremet, T.; Kirkin, A.F.; Zeuthen, J. Somatic mutation of the Peutz-Jeghers syndrome gene, LKB1/STK11, in malignant melanoma. Oncogene 1999, 18, 1777–1780. [Google Scholar] [CrossRef] [PubMed]
- Rowan, A.; Bataille, V.; MacKie, R.; Healy, E.; Bicknell, D.; Bodmer, W.; Tomlinson, I. Somatic mutations in the Peutz-Jegners (LKB1/STKII) gene in sporadic malignant melanomas. J. Investig. Dermatol. 1999, 112, 509–511. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Schonleben, F.; Thaker, H.M.; Goggins, M.; Su, G.H. A novel mutation of STK11/LKB1 gene leads to the loss of cell growth inhibition in head and neck squamous cell carcinoma. Oncogene 2006, 25, 2937–2942. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.J.; Cho, Y.G.; Park, J.Y.; Kim, T.Y.; Lee, J.H.; Kim, H.S.; Lee, J.W.; Song, Y.H.; Nam, S.W.; Lee, S.H.; et al. Genetic analysis of the LKB1/STK11 gene in hepatocellular carcinomas. Eur. J. Cancer 2004, 40, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Ylikorkala, A.; Rossi, D.J.; Korsisaari, N.; Luukko, K.; Alitalo, K.; Henkemeyer, M.; Makela, T.P. Vascular abnormalities and deregulation of VEGF in LKB1-deficient mice. Science 2001, 293, 1323–1326. [Google Scholar] [CrossRef] [PubMed]
- Jishage, K.; Nezu, J.; Kawase, Y.; Iwata, T.; Watanabe, M.; Miyoshi, A.; Ose, A.; Habu, K.; Kake, T.; Kamada, N.; et al. Role of LKB1, the causative gene of Peutz-Jegher’s syndrome, in embryogenesis and polyposis. Proc. Natl. Acad. Sci. USA 2002, 99, 8903–8908. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, H.; Nakau, M.; Ishikawa, T.; Seldin, M.F.; Oshima, M.; Taketo, M.M. Gastrointestinal hamartomatous polyposis in LKB1 heterozygous knockout mice. Cancer Res. 2002, 62, 2261–2266. [Google Scholar] [PubMed]
- Nakau, M.; Miyoshi, H.; Seldin, M.F.; Imamura, M.; Oshima, M.; Taketo, M.M. Hepatocellular carcinoma caused by loss of heterozygosity in LKB1 gene knockout mice. Cancer Res. 2002, 62, 4549–4553. [Google Scholar] [PubMed]
- Rossi, D.J.; Ylikorkala, A.; Korsisaari, N.; Salovaara, R.; Luukko, K.; Launonen, V.; Henkemeyer, M.; Ristimaki, A.; Aaltonen, L.A.; Makela, T.P. Induction of cyclooxygenase-2 in a mouse model of Peutz-Jeghers polyposis. Proc. Natl. Acad. Sci. USA 2002, 99, 12327–12332. [Google Scholar] [CrossRef] [PubMed]
- Koh, H.J.; Arnolds, D.E.; Fujii, N.; Tran, T.T.; Rogers, M.J.; Jessen, N.; Li, Y.F.; Liew, C.W.; Ho, R.C.; Hirshman, M.F.; et al. Skeletal muscle-selective knockout of LKB1 increases insulin sensitivity, improves, glucose homeostasis, and decreases TRB3. Mol. Cell. Biol. 2006, 26, 8217–8227. [Google Scholar] [CrossRef] [PubMed]
- Londesborough, A.; Vaahtomeri, K.; Tiainen, M.; Katajisto, P.; Ekman, N.; Vallenius, T.; Makela, T.P. LKB1 in endothelial cells is required for angiogenesis and TGFβ-mediated vascular smooth muscle cell recruitment. Development 2008, 135, 2331–2338. [Google Scholar] [CrossRef] [PubMed]
- Vaahtomeri, K.; Ventela, E.; Laajanen, K.; Katajisto, P.; Wipff, P.J.; Hinz, B.; Vallenius, T.; Tiainen, M.; Makela, T.P. LKB1 is required for TGFβ-mediated myofibroblast differentiation. J. Cell Sci. 2008, 121, 3531–3540. [Google Scholar] [CrossRef] [PubMed]
- Towler, M.C.; Fogarty, S.; Hawley, S.A.; Pan, D.A.; Martin, D.M.A.; Morrice, N.A.; McCarthy, A.; Galardo, M.N.; Meroni, S.B.; Cigorraga, S.B.; et al. A novel short splice variant of the tumour suppressor LKB1 is required for spermiogenesis. Biochem. J. 2008, 416, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.; Nye, E.; Stamp, G.; Silver, A. Osteogenic tumours in LKB1-deficient mice. Exp. Mol. Pathol. 2008, 85, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Pearson, H.B.; McCarthy, A.; Collins, C.M.P.; Ashworth, A.; Clarke, A.R. LKB1 deficiency causes prostate neoplasia in the mouse. Cancer Res. 2008, 68, 2223–2232. [Google Scholar] [CrossRef] [PubMed]
- Denison, F.C.; Smith, L.B.; Muckett, P.J.; O’Hara, L.; Carling, D.; Woods, A. LKB1 is an essential regulator of spermatozoa release during spermiation in the mammalian testis. PLoS One 2011, 6, e28306. [Google Scholar] [CrossRef] [PubMed]
- Tanwar, P.S.; Kaneko-Tarui, T.; Zhang, L.; Tanaka, Y.; Crum, C.P.; Teixeira, J.M. Stromal liver kinase B1 [STK11] signaling loss induces oviductal adenomas and endometrial cancer by activating mammalian target of Rapamycin complex 1. PLoS Genet. 2012, 8, e1002906. [Google Scholar] [CrossRef] [PubMed]
- Tanwar, P.S.; Kaneko-Tarui, T.; Zhang, L.; Teixeira, J.M. Altered LKB1/AMPK/TSC1/TSC2/mTOR signaling causes disruption of Sertoli cell polarity and spermatogenesis. Hum. Mol. Genet. 2012, 21, 4394–4405. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, Q.; Song, P.; Zou, M. Liver kinase B1 is required for white adipose tissue growth and differentiation. Diabetes 2013, 62, 2347–2358. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, J.; Maarbjerg, S.J.; Jordy, A.B.; Fritzen, A.M.; Pehmoller, C.; Sylow, L.; Serup, A.K.; Jessen, N.; Thorsen, K.; Prats, C.; et al. LKB1 regulates lipid oxidation during exercise independently of AMPK. Diabetes 2013, 62, 1490–1499. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Vieira, R.; Xu, Z.; Colp, P.; Marignani, P.A. Loss of LKB1 expression reduces the latency of ErbB2-mediated mammary gland tumorigenesis, promoting changes in metabolic pathways. PLoS One 2013, 8, e56567. [Google Scholar] [CrossRef] [PubMed]
- Mehenni, H.; Gehrig, C.; Nezu, J.; Oku, A.; Shimane, M.; Rossier, C.; Guex, N.; Blouin, J.L.; Scott, H.S.; Antonarakis, S.E. Loss of LKB1 kinase activity in Peutz-Jeghers syndrome, and evidence for allelic and locus heterogeneity. Am. J. Hum. Genet. 1998, 63, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.P.; Spicer, J.; Smith, A.; Swift, S.; Ashworth, A. The mouse Peutz-Jeghers syndrome gene LKB1 encodes a nuclear protein kinase. Hum. Mol. Genet. 1999, 8, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Su, J.Y.; Erikson, E.; Maller, J.L. Cloning and characterization of a novel serine/threonine protein kinase expressed in early Xenopus embryos. J. Biol. Chem. 1996, 271, 14430–14437. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.L.; Morton, D.G.; Bestman, J.; Kemphues, K.J. The C. elegans Par-4 gene encodes a putative serine-threonine kinase required for establishing embryonic asymmetry. Development 2000, 127, 1467–1475. [Google Scholar] [PubMed]
- Martin, S.G.; St Johnston, D. A role for Drosophila LKB1 in anterior-posterior axis formation and epithelial polarity. Nature 2003, 421, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Churchman, M.; Dowling, B.; Tomlinson, I.P.M. Identification of a novel mRNA species of the LKB1/STK11 Peutz-Jeghers serine/threonine kinase. DNA Seq. 1999, 10, 255–261. [Google Scholar] [PubMed]
- Abed, A.A.; Gunther, K.; Kraus, C.; Hohenberger, W.; Ballhausen, W.G. Mutation screening at the RNA level of the STK11/LKB-1 gene in Peutz-Jeghers syndrome reveals complex splicing abnormalities and a novel mRNA isoform (STK11 c.597 (boolean AND) 598insIVS4). Hum. Mut. 2001, 18, 397–410. [Google Scholar]
- Luukko, K.; Ylikorkala, A.; Tiainen, M.; Makela, T.P. Expression of LKB1 and PTEN tumor suppressor genes during mouse embryonic development. Mech. Dev. 1999, 83, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Rowan, A.; Churchman, M.; Jefferey, R.; Hanby, A.; Poulsom, R.; Tomlinson, I. In situ analysis of LKB1/STK11 mRNA expression in human normal tissues and tumours. J. Pathol. 2000, 192, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Ylikorkala, A.; Avizienyte, E.; Tomlinson, I.P.M.; Tiainen, M.; Roth, S.; Loukola, A.; Hemminki, A.; Johansson, M.; Sistonen, P.; Markie, D.; et al. Mutations and impaired function of LKB1 in familial and non-familial Peutz-Jeghers syndrome and a sporadic testicular cancer. Hum. Mol. Genet. 1999, 8, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.P.; Reoma, J.L.; Gamm, D.M.; Uhler, M.D. LKB1, a novel serine/threonine protein kinase and potential tumour suppressor, is phosphorylated by cAMP-dependent protein kinase (PKA) and prenylated in vivo. Biochem. J. 2000, 345, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Nezu, J.; Oku, A.; Shimane, M. Loss of cytoplasmic retention ability of mutant LKB1 found in Peutz-Jeghers syndrome patients. Biochem. Biophys. Res. Commun. 1999, 261, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Tiainen, M.; Vaahtomeri, K.; Ylikorkala, A.; Makela, T.P. Growth arrest by the LKB1 tumor suppressor: Induction of p21 (WAF1/CIP1). Hum. Mol. Genet. 2002, 11, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Karuman, P.; Gozani, O.; Odze, R.D.; Zhou, X.C.; Zhu, H.; Shaw, R.; Brien, T.P.; Bozzuto, C.D.; Ooi, D.; Cantley, L.C.; et al. The Peutz-Jegher gene product LKB1 is a mediator of p53-dependent cell death. Mol. Cell 2001, 7, 1307–1319. [Google Scholar] [CrossRef] [PubMed]
- Qanungo, S.; Haldar, S.; Basu, A. Restoration of silences Peutz-Jeghers syndrome gene, LKB1 induces apoptosis in pancreatic carinoma cells. Neoplasia 2003, 5, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Brajenovic, M.; Joberty, G.; Kuster, B.; Bouwmeester, T.; Drewes, G. Comprehensive proteomic analysis of human par protein complexes reveals an interconnected protein network. J. Biol. Chem. 2004, 279, 12804–12811. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.P.; Rayter, S.I.; Niederlander, C.; Spicer, J.; Jones, C.M.; Ashworth, A. LIP1, a cytoplasmic protein functionally linked to the Peutz-Jeghers syndrome kinase LKB1. Hum. Mol. Genet. 2001, 10, 2869–2877. [Google Scholar] [CrossRef] [PubMed]
- Baas, A.F.; Boudeau, J.; Sapkota, G.P.; Smit, L.; Medema, R.; Morrice, N.A.; Alessi, D.R.; Clevers, H.C. Activation of the tumour suppressor kinase LKB1 by the STE20-like pseudokinase STRAD. EMBO J. 2003, 22, 3062–3072. [Google Scholar] [CrossRef] [PubMed]
- Boudeau, J.; Baas, A.F.; Deak, M.; Morrice, N.A.; Kieloch, A.; Schutkowski, M.; Prescott, A.R.; Clevers, H.C.; Alessi, D.R. MO25α/β interact with STRADα/β enhancing their ability to bind, activate and localize LKB1 in the cytoplasm. EMBO J. 2003, 22, 5102–14. [Google Scholar] [CrossRef] [PubMed]
- Boudeau, J.; Scott, J.W.; Resta, N.; Deak, M.; Kieloch, A.; Komander, D.; Hardie, D.G.; Prescott, A.R.; van Aalten, D.M.F; Alessi, D.R. Analysis of the LKB1–STRAD–MO25 complex. J. Cell Sci. 2004, 117, 6365–6375. [Google Scholar]
- Zeqiraj, E.; Filippi, B.M.; Deak, M.; Alessi, D.R.; van Aalten, D.M.F. Structure of the LKB1–STRAD–MO25 complex reveals an allosteric mechanism of kinase activation. Science 2009, 326, 1707–1711. [Google Scholar] [CrossRef] [PubMed]
- Milburn, C.C.; Boudeau, J.; Deak, M.; Alessi, D.R.; van Aalten, D.M.F. Crystal structure of MO25α in complex with the C terminus of the pseudo kinase STE20-related adaptor. Nat. Struct. Mol. Biol. 2004, 11, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Nony, P.; Gaude, H.; Rossel, M.; Fournier, L.; Rouault, J.P.; Billaud, M. Stability of the Peutz-Jeghers syndrome kinase LKB1 requires its binding to the molecular chaperones Hsp90/Cdc37. Oncogene 2003, 22, 9165–9175. [Google Scholar] [CrossRef] [PubMed]
- Boudeau, J.; Deak, M.; Lawlor, M.A.; Morrice, N.A.; Alessi, D.R. Heat-shock protein 90 and Cdc37 interact with LKB1 and regulate its stability. Biochem. J. 2003, 370, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Gaude, H.; Aznar, N.; Delay, A.; Bres, A.; Buchet-Poyau, K.; Caillat, C.; Vigouroux, A.; Rogon, C.; Woods, A.; Vanacker, J.M.; et al. Molecular chaperone complexes with antagonizing activities regulate stability and activity of the tumor suppressor LKB1. Oncogene 2012, 31, 1582–1591. [Google Scholar] [CrossRef] [PubMed]
- Mehenni, H.; Lin-Marq, N.; Buchet-Poyau, K.; Reymond, A.; Collart, M.A.; Picard, D.; Antonarakis, S.E. LKB1 interacts with and phosphorylates PTEN: A functional link between two proteins involved in cancer predisposing syndromes. Hum. Mol. Genet. 2005, 14, 2209–2219. [Google Scholar] [CrossRef] [PubMed]
- Marignani, P.A.; Kanai, F.; Carpenter, C.L. LKB1 associates with Brg1 and is necessary for Brg1-induced growth arrest. J. Biol. Chem. 2001, 276, 32415–32418. [Google Scholar] [CrossRef] [PubMed]
- Blumer, J.B.; Bernard, M.L.; Peterson, Y.K.; Nezu, J.; Chung, P.; Dunican, D.J.; Knoblich, J.A.; Lanier, S.M. Interaction of activator of G-protein signaling 3 (AGS3) with LKB1, a serine/threonine kinase involved in cell polarity and cell cycle progression—Phosphorylation of the G-protein regulatory (GPR) motif as a regulatory mechanism for the interaction of GPR motifs with G(i)α. J. Biol. Chem. 2003, 278, 23217–23220. [Google Scholar] [CrossRef] [PubMed]
- Clements, W.K.; Kimelman, D. Wnt signalling gets XEEKy. Nat. Cell Biol. 2003, 5, 861–863. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, N.; Sun, Y.L.; Chen, S.J.; Paul, P.; Shaw, R.J.; Cantley, L.C.; Price, B.D. DNA damage-induced association of ATM with its target proteins requires a protein interaction domain in the N terminus of ATM. J. Biol. Chem. 2005, 280, 15158–15164. [Google Scholar] [CrossRef] [PubMed]
- Setogawa, T.; Shinozaki-Yabana, S.; Masuda, T.; Matsuura, K.; Akiyama, T. The tumor suppressor LKB1 induces p21 expression in collaboration with LMO4, GATA-6, and Ldb1. Biochem. Biophys. Res. Commun. 2006, 343, 1186–1190. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M.; Avizienyte, E.; Corn, P.G.; Lothe, R.A.; Baylin, S.B.; Aaltonen, L.A.; Herman, J.G. Epigenetic inactivation of LKB1 in primary tumors associated with the Peutz-Jeghers syndrome. Oncogene 2000, 19, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Trojan, J.; Brieger, A.; Raedle, J.; Esteller, M.; Zeuzem, S. 5'-CpG island methylation of the LKB1/STK11 promoter and allelic loss at chromosome 19p13.3 in sporadic colorectal cancer. Gut 2000, 47, 272–276. [Google Scholar]
- Brown, K.A.; McInnes, K.J.; Takagi, K.; Ono, K.; Hunger, N.I.; Wang, L.; Sasano, H.; Simpson, E.R. LKB1 expression is inhibited by estradiol-17β in MCF-7 cells. J. Steroid Biochem. Mol. Biol. 2011, 127, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Linher-Melville, K.; Zantinge, S.; Singh, G. Liver kinase B1 expression (LKB1) is repressed by estrogen receptor alpha (ERα) in MCF-7 human breast cancer cells. Biochem. Biophys. Res. Commun. 2012, 417, 1063–1068. [Google Scholar] [CrossRef]
- McInnes, K.J.; Brown, K.A.; Hunger, N.I.; Simpson, E.R. Regulation of LKB1 expression by sex hormones in adipocytes. Int. J. Obes. 2012, 36, 982–985. [Google Scholar] [CrossRef]
- Han, S.W.; Khuri, F.R.; Roman, J. Fibronectin stimulates non-small cell lung carcinoma cell growth through activation of Akt/mammalian target of rapamycin/S6 kinase and inactivation of LKB1/AMP-activated protein kinase signal pathways. Cancer Res. 2006, 66, 315–323. [Google Scholar] [CrossRef]
- Kimball, S.R.; Siegfried, B.A.; Jefferson, L.S. Glucagon represses signaling through the mammalian target of rapamycin in rat liver by activating AMP-activated protein kinase. J. Biol. Chem. 2004, 279, 54103–54109. [Google Scholar] [CrossRef] [PubMed]
- Feiger, Z.J.; Maizels, E.T.; Flynn, M.P.; Hunzicker-Dunn, M. FSH elicits C-terminal phosphorylation of the serine threonine kinase LKB1. Biol. Reprod. 2005, 2005, 245–245. [Google Scholar]
- Xie, Z.L.; Dong, Y.Z.; Zhang, M.; Cui, M.Z.; Cohen, R.A.; Riek, U.; Neumann, D.; Schlattner, U.; Zou, M.H. Activation of protein kinase Cζ by peroxynitrite regulates LKB1-dependent AMP-activated protein kinase in cultured endothelial cells. J. Biol. Chem. 2006, 281, 6366–6375. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, G.P.; Boudeau, J.; Deak, M.; Kieloch, A.; Morrice, N.; Alessi, D.R. Identification and characterization of four novel phosphorylation sites (Ser(31), Ser(325), Thr(336) and Thr(366)) on LKB1/STK11, the protein kinase mutated in Peutz-Jeghers cancer syndrome. Biochem. J. 2002, 362, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, G.P.; Deak, M.; Kieloch, A.; Morrice, N.; Goodarzi, A.A.; Smythe, C.; Shiloh, Y.; Lees-Miller, S.P.; Alessi, D.R. Ionizing radiation induces ataxia telangiectasia mutated kinase (ATM)-mediated phosphorylation of LKB1/STK11 at Thr-366. Biochem. J. 2002, 368, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, G.P.; Kieloch, A.; Lizcano, J.M.; Lain, S.; Arthur, J.S.C.; Williams, M.R.; Morrice, N.; Deak, M.; Alessi, D.R. Phosphorylation of the protein kinase mutated in Peutz-Jeghers cancer syndrome, LKB1/STK11, at Ser(431) by p90(RSK) and cAMP-dependent protein kinase, but not its farnesylation at Cys(433), is essential for LKB1 to suppress cell growth. J. Biol. Chem. 2001, 276, 19469–19482. [Google Scholar] [CrossRef] [PubMed]
- Zu, Y.; Liu, L.; Lee, M.Y.K.; Xu, C.; Liang, Y.; Man, R.Y.; Vanhoutte, P.M.; Wang, Y. SIRT1 promotes proliferation and prevents senescence through targeting LKB1 in primary porcine aortic endothelial cells. Circ. Res. 2010, 106, 1384–1393. [Google Scholar] [CrossRef] [PubMed]
- Lan, F.; Cacicedo, J.M.; Ruderman, N.; Ido, Y. SIRT1 modulation of the acetylation status, cytosolic localization, and activity of LKB1—Possible role in AMP-activated protein kinase activation. J. Biol. Chem. 2008, 283, 27628–27635. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.Y.; Xu, S.Q.; Maitland-Toolan, K.A.; Sato, K.; Jiang, B.B.; Ido, Y.S.; Lan, F.; Walsh, K.; Wierzbicki, M.; Verbeuren, T.J.; et al. SIRT1 regulates hepatocyte lipid metabolism through activating AMP-activated protein kinase. J. Biol. Chem. 2008, 283, 20015–20026. [Google Scholar] [CrossRef] [PubMed]
- Lizcano, J.M.; Goransson, O.; Toth, R.; Deak, M.; Morrice, N.A.; Boudeau, J.; Hawley, S.A.; Udd, L.; Makela, T.P.; Hardie, D.G.; et al. LKB1 is a master kinase that activates 13 kinases of the AMPK subfamily, including MARK/PAR-1. EMBO J. 2004, 23, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, M.; Villa, F.; Deak, M.; Toth, R.; Prescott, A.R.; van Aalten, D.M.F.; Alessi, D.R. The ubiquitin-associated domain of AMPK-related kinases regulates conformation and LKB1-mediated phosphorylation and activation. Biochem. J. 2006, 394, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.A.; Lodish, H.F. A revised model for AMP-activated protein kinase structure—The α-subunit binds to both the β- and γ-subunits although there is no direct binding between the β- and γ-subunits. J. Biol. Chem. 2006, 281, 36434–36442. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.P.; Leiper, F.C.; Woods, A.; Carling, D.; Carlson, M. Activation of yeast SNF1 and mammalian AMP-activated protein kinase by upstream kinases. Proc. Natl. Acad. Sci. USA 2003, 100, 8839–8843. [Google Scholar] [CrossRef] [PubMed]
- Shaw, R.J.; Kosmatka, M.; Bardeesy, N.; Hurley, R.L.; Witters, L.A.; DePinho, R.A.; Cantley, L.C. The tumor suppressor LKB1 kinase directly activates AMP-activated kinase and regulates apoptosis in response to energy stress. Proc. Natl. Acad. Sci. USA 2004, 101, 3329–3335. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.; Johnstone, S.R.; Dickerson, K.; Leiper, F.C.; Fryer, L.G.D.; Neumann, D.; Schlattner, U.; Wallimann, T.; Carlson, M.; Carling, D. LKB1 is the upstream kinase in the AMP-activated protein kinase cascade. Curr. Biol. 2003, 13, 2004–2008. [Google Scholar] [CrossRef] [PubMed]
- Drewes, G.; Ebneth, A.; Preuss, U.; Mandelkow, E.M.; Mandelkow, E. MARK, a novel family of protein kinases that phosphorylate microtubule-associated proteins and trigger microtubule disruption. Cell 1997, 89, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Goransson, O.; Deak, M.; Wullschleger, S.; Morrice, N.A.; Prescott, A.R.; Alessi, D.R. Regulation of the polarity kinases PAR-1/MARK by 14-3-3 interaction and phosphorylation. J. Cell Sci. 2006, 119, 4059–4070. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Imai, Y.; Lu, B.W. Activation of PAR-1 kinase and stimulation of tau phosphorylation by diverse signals require the tumor suppressor protein LKB1. J. Neurosci. 2007, 27, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Kojima, Y.; Miyoshi, H.; Clevers, H.C.; Oshima, M.; Aoki, M.; Taketo, M.M. Suppression of tubulin polymerization by the LKB1-microtubule-associated protein/microtubule affinity-regulating kinase signaling. J. Biol. Chem. 2007, 282, 23532–23540. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, Y.K.; Satoh, T.; Okamoto, M.; Takemori, H. Importance of autophosphorylation at Ser186 in the A-loop of salt inducible kinase 1 for its sustained kinase activity. J. Cell. Biochem. 2008, 104, 1724–1739. [Google Scholar] [CrossRef] [PubMed]
- Screaton, R.A.; Conkright, M.D.; Katoh, Y.; Best, J.L.; Canettieri, G.; Jeffries, S.; Guzman, E.; Niessen, S.; Yates, J.R.; Takemori, H.; et al. The CREB coactivator TORC2 functions as a calcium- and cAMP-sensitive coincidence detector. Cell 2004, 119, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Katoh, Y.; Takemori, H.; Lin, X.; Tamura, M.; Muraoka, M.; Satoh, T.; Tsuchiya, Y.; Min, L.; Doi, J.; Miyauchi, A.; et al. Silencing the constitutive active transcription factor CREB by the LKB1–SIK signaling cascade. FEBS J. 2006, 273, 2730–2748. [Google Scholar] [CrossRef] [PubMed]
- Shaw, R.J.; Lamia, K.A.; Vasquez, D.; Koo, S.H.; Bardeesy, N.; DePinho, R.A.; Montminy, M.; Cantley, L.C. The kinase LKB1 mediates glucose homeostasis in liver and therapeutic effects of metformin. Science 2005, 310, 1642–1646. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, M.; McBride, A.; Lizcano, J.M.; Deak, M.; Toth, R.; Morrice, N.A.; Alessi, D.R. Identification of the sucrose non-fermenting related kinase SNRK, as a novel LKB1 substrate. FEBS Lett. 2005, 579, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, D.L.; Rosen, C.F. Regulation of SNARK activity in response to cellular stresses. Biochim. Biophys. Acta 2005, 1724, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Kuga, W.; Tsuchihara, K.; Ogura, T.; Kanehara, S.; Saito, M.; Suzuki, A.; Esumi, H. Nuclear localization of SNARK; its impact on gene expression. Biochem. Biophys. Res. Commun. 2008, 377, 1062–1066. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Takashima, S.; Shintani, Y.; Yamazaki, S.; Seguchi, O.; Nakano, A.; Higo, S.; Kato, H.; Liao, Y.; Asano, Y.; et al. Identification of a novel substrate for TNFα-induced kinase NUAK2. Biochem. Biophys. Res. Commun. 2008, 365, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Bright, N.J.; Carling, D.; Thornton, C. Investigating the regulation of brain-specific kinases 1 and 2 by phosphorylation. J. Biol. Chem. 2008, 283, 14946–14954. [Google Scholar] [CrossRef] [PubMed]
- Barnes, A.P.; Lilley, B.N.; Pan, Y.A.; Plummer, L.J.; Powell, A.W.; Raines, A.N.; Sanes, J.R.; Polleux, F. LKB1 and SAD kinases define a pathway required for the polarization of cortical neurons. Cell 2007, 129, 549–563. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G.; Alessi, D.R. LKB1 and AMPK and the cancer-metabolism link—Ten years after. BMC Biol. 2013, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.Q.; Saha, A.K.; Wen, R.; Ruderman, N.B.; Luo, Z.J. AMP-activated protein kinase activators can inhibit the growth of prostate cancer cells by multiple mechanisms. Biochem. Biophys. Res. Commun. 2004, 321, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. New roles for the LKB1–AMPK pathway. Curr. Opin. Cell Biol. 2005, 17, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Zakikhani, M.; Dowling, R.; Fantus, I.G.; Sonenberg, N.; Pollak, M. Metformin is an AMP kinase-dependent growth inhibitor for breast cancer cells. Cancer Res. 2006, 66, 10269–10273. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.G.; Plas, D.R.; Kubek, S.; Buzzai, M.; Mu, J.; Xu, Y.; Birnbaum, M.J.; Thompson, C.B. AMP-activated protein kinase induces a p53-dependent metabolic checkpoint. Mol. Cell 2005, 18, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.T.; Kim, Y.M.; Surh, Y.J.; Baik, H.W.; Lee, S.Y.; Ha, J.; Park, O.J. Selenium regulates cyclooxygenase-2 and extracellular signal-regulated kinase signaling pathways by activating AMP-activated protein kinase in colon cancer cells. Cancer Res. 2006, 66, 10057–10063. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.; Liu, C.Y.; Chatterjee, A.; Hoque, M.O.; Kim, M.S.; Engles, J.; Westra, W.; Trink, B.; Ratovitski, E.; Sidransky, D. LKB1/STK11 suppresses cyclooxygenase-2 induction and cellular invasion through PEA3 in lung cancer. Cancer Res. 2006, 66, 7870–7879. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.M.; Mullally, J.E.; Fitzpatrick, F.A. Reactive lipid species from cyclooxygenase-2 inactivate tumor suppressor LKB1/STK11—Cyclopentenone prostaglandins and 4-hydroxy-2-nonenal covalently modify and inhibit the AMP-kinase kinase that modulates cellular energy homeostasis and protein translation. J. Biol. Chem. 2006, 281, 2598–2604. [Google Scholar] [CrossRef] [PubMed]
- Zeng, P.Y.; Berger, S.L. LKB1 is recruited to the p21/WAF1 promoter by p53 to mediate transcriptional activation. Cancer Res. 2006, 66, 10701–10708. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.R.; Hay, M.P. Targeting hypoxia in cancer therapy. Nat. Rev. Cancer 2011, 11, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Powis, G.; Kirkpatrick, L. Hypoxia inducible factor-1α as a cancer drug target. Mol. Cancer Ther. 2004, 3, 647–654. [Google Scholar] [PubMed]
- Shaw, R.J.; Bardeesy, N.; Manning, B.D.; Lopez, L.; Kasmatka, M.; de Pinho, R.A.; Cantley, L.C. The LKB1 tumor suppressor negatively regulates mTOR signaling. Cancer Cell 2004, 6, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.V.; Tran, C.; Mellinghoff, I.K.; Welsbie, D.S.; Chan, E.; Fueger, B.; Czernin, J.; Sawyers, C.L. Hypoxia-inducible factor determines sensitivity to inhibitors of mTOR in kidney cancer. Nat. Med. 2006, 12, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Shackelford, D.B.; Vasquez, D.S.; Corbeil, J.; Wu, S.L.; Leblanc, M.; Wu, C.L.; Vera, D.R.; Shaw, R.J. mTOR and HIF-1α-mediated tumor metabolism in an LKB1 mouse model of Peutz-Jeghers syndrome. Proc. Natl. Acad. Sci. USA 2009, 106, 11137–11142. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Koh, H.; Kim, M.; Park, J.; Lee, S.Y.; Lee, S.; Chung, J. JNK pathway mediates apoptotic cell death induced by tumor suppressor LKB1 in Drosophila. Cell Death Differ. 2006, 13, 1110–1122. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Iwai, A.; Nakashima, M.; Fujikura, D.; Chiba, S.; Li, H.M.; Uehara, J.; Kawaguchi, S.; Kaya, M.; Nagoya, S.; et al. LKB1 is crucial for TRAIL-mediated apoptosis induction in osteosarcoma. Anticancer Res. 2007, 27, 761–768. [Google Scholar] [PubMed]
- Shaw, R.J. LKB1 and AMP-activated protein kinase control of mTOR signalling and growth. Acta Physiol. 2009, 196, 65–80. [Google Scholar] [CrossRef]
- Zhuang, Z.G.; Di, G.H.; Shen, Z.Z.; Ding, J.; Shao, Z.M. Enhanced expression of LKB1 in breast cancer cells attenuates angiogenesis, invasion, and metastatic potential. Mol. Cancer Res. 2006, 4, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Taliaferro-Smith, L.; Nagalingam, A.; Zhong, D.; Zhou, W.; Saxena, N.K.; Sharma, D. LKB1 is required for adiponectin-mediated modulation of AMPK-S6K axis and inhibition of migration and invasion of breast cancer cells. Oncogene 2009, 28, 2621–2633. [Google Scholar] [CrossRef] [PubMed]
- Udd, L.; Katajisto, P.; Rossi, D.J.; Lepisto, A.; Lahesmaa, A.M.; Ylikorkala, A.; Jarvinen, H.J.; Ristimaki, A.P.; Makela, T.P. Suppression of Peutz-Jeghers polyposis by inhibition of cyclooxygenase-2. Gastroenterology 2004, 127, 1030–1037. [Google Scholar] [CrossRef]
- Xiao, X.X.; He, Q.Q.; Lu, C.M.; Werle, K.D.; Zhao, R.X.; Chen, J.F.; Davis, B.C.; Cui, R.T.; Liang, J.Y.; Xu, Z.X. Metformin impairs the growth of liver kinase B1-intact cervical cancer cells. Gynecol. Oncol. 2012, 127, 249–255. [Google Scholar] [CrossRef]
- Contreras, C.M.; Akbay, E.A.; Gallardo, T.D.; Haynie, J.M.; Sharma, S.; Tagao, O.; Bardeesy, N.; Takahashi, M.; Settleman, J.; Wong, K.K.; et al. LKB1 inactivation is sufficient to drive endometrial cancers that are aggressive yet highly responsive to mTOR inhibitor monotherapy. Dis. Models Mech. 2010, 3, 181–193. [Google Scholar] [CrossRef]
- Park, S.Y.; Park, O.J. Modulation of cancer cell proliferation by cell survival signal Akt and tumor suppressive energy sensor AMP-activated Protein kinase in colon cancer cells treated with resveratrol. Food Sci. Biotechnol. 2010, 19, 1537–1541. [Google Scholar] [CrossRef]
- Jeon, S.M.; Chandel, N.S.; Hay, N. AMPK regulates NADPH homeostasis to promote tumour cell survival during energy stress. Nature 2012, 485, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ulbrich, J.; Muller, J.; Wustefeld, T.; Aeberhard, L.; Kress, T.R.; Muthalagu, N.; Rycak, L.; Rudalska, R.; Moll, R.; et al. Deregulated MYC expression induces dependence upon AMPK-related kinase 5. Nature 2012, 483, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Shackelford, D.B.; Abt, E.; Gerken, L.; Vasquez, D.S.; Seki, A.; Leblanc, M.; Wei, L.; Fishbein, M.C.; Czernin, J.; Mischel, P.S.; et al. LKB1 Inactivation dictates therapeutic response of non-small cell lung cancer to the metabolism drug phenformin. Cancer Cell 2013, 23, 143–158. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gan, R.-Y.; Li, H.-B. Recent Progress on Liver Kinase B1 (LKB1): Expression, Regulation, Downstream Signaling and Cancer Suppressive Function. Int. J. Mol. Sci. 2014, 15, 16698-16718. https://doi.org/10.3390/ijms150916698
Gan R-Y, Li H-B. Recent Progress on Liver Kinase B1 (LKB1): Expression, Regulation, Downstream Signaling and Cancer Suppressive Function. International Journal of Molecular Sciences. 2014; 15(9):16698-16718. https://doi.org/10.3390/ijms150916698
Chicago/Turabian StyleGan, Ren-You, and Hua-Bin Li. 2014. "Recent Progress on Liver Kinase B1 (LKB1): Expression, Regulation, Downstream Signaling and Cancer Suppressive Function" International Journal of Molecular Sciences 15, no. 9: 16698-16718. https://doi.org/10.3390/ijms150916698
APA StyleGan, R.-Y., & Li, H.-B. (2014). Recent Progress on Liver Kinase B1 (LKB1): Expression, Regulation, Downstream Signaling and Cancer Suppressive Function. International Journal of Molecular Sciences, 15(9), 16698-16718. https://doi.org/10.3390/ijms150916698