Chromatin Structure and Dynamics in Hot Environments: Architectural Proteins and DNA Topoisomerases of Thermophilic Archaea



Abstract
:1. Introduction
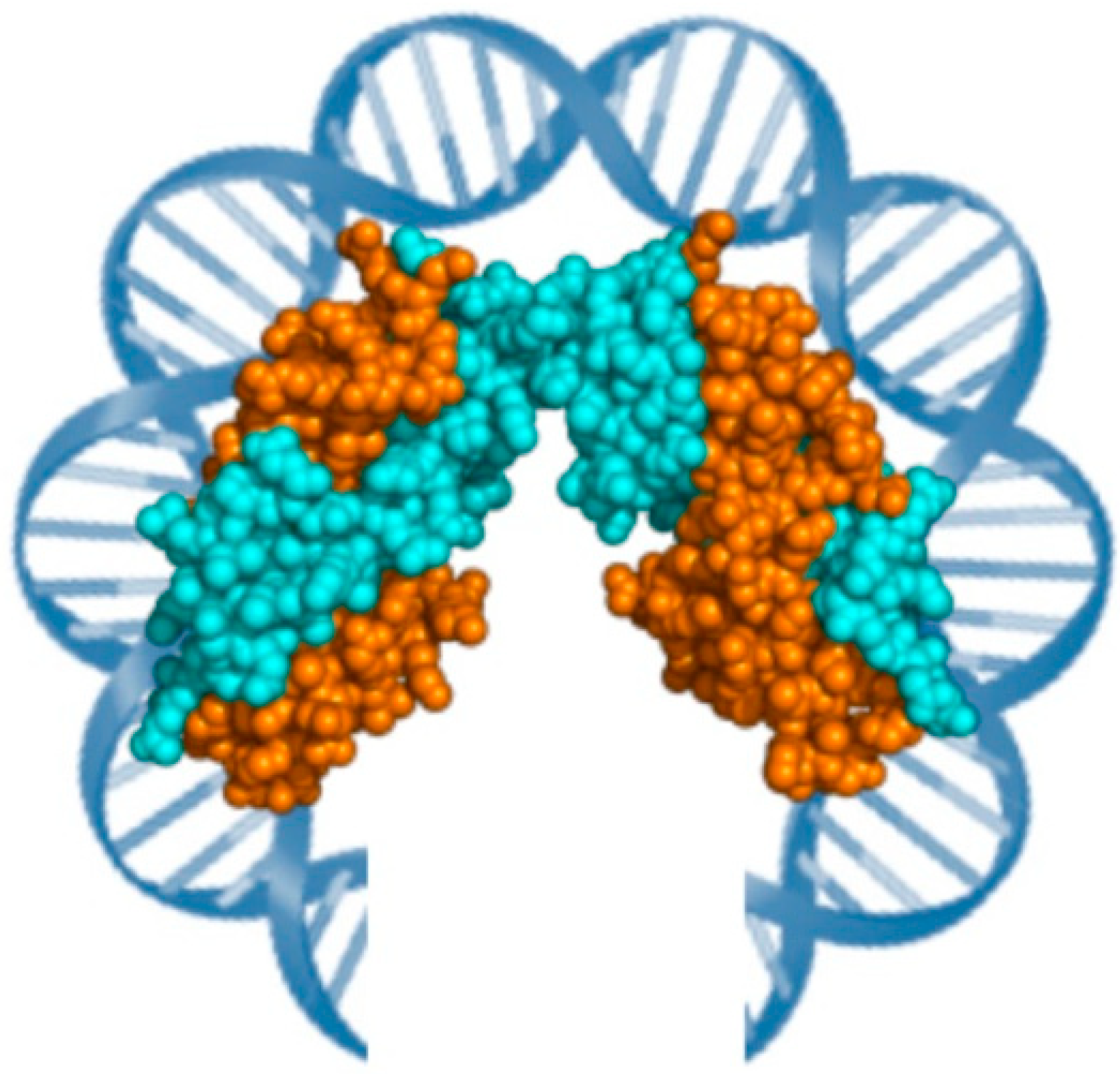
2. Architectual Proteins of Hyperthermophilic Archaea

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | HISTONE | ALBA | SUL7 | CREN7 | SMJ12 |
|---|---|---|---|---|---|
| Archaeal sub-domain | Euryarchaea and Crenarchaea | Euryarchaea and Crenarchaea | Crenarchaea (Sulfolobus) | Crenarchaea | Crenarchaea (S. solfataricus) |
| Oligomeric Structure | Dimer (7.5 kDa) | Dimer (10 kDa) | Monomer (7 kDa) | Monomer (7 kDa) | Dimer (12 kDa) |
| DNA Binding | Cooperative | Cooperative | - | - | - |
| DNA Modification | Compaction | √ | - | √ | √ |
| Bending | √ | - | √ | √ | |
| Bridging | - | √ | - | - | |
| Supercoiling | Negative | Negative | Negative | Negative | |
| Post-translational modifications | NO | Acetylation/Deacetylation | Methylation | Methylation | unknown |
2.1. Histones

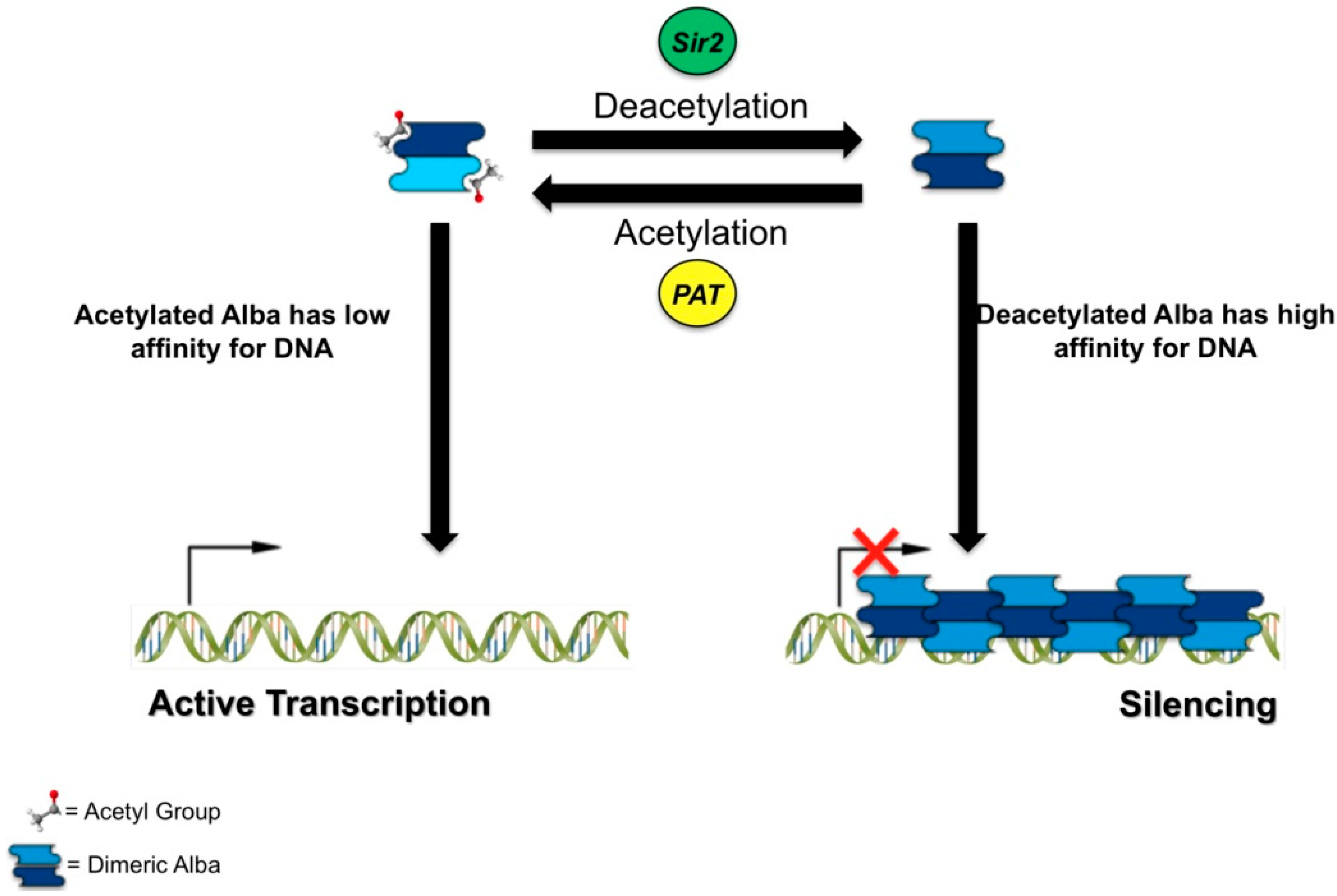
2.2. Alba

2.3. Sul7 and Cren7
2.4. Other Architectural Proteins
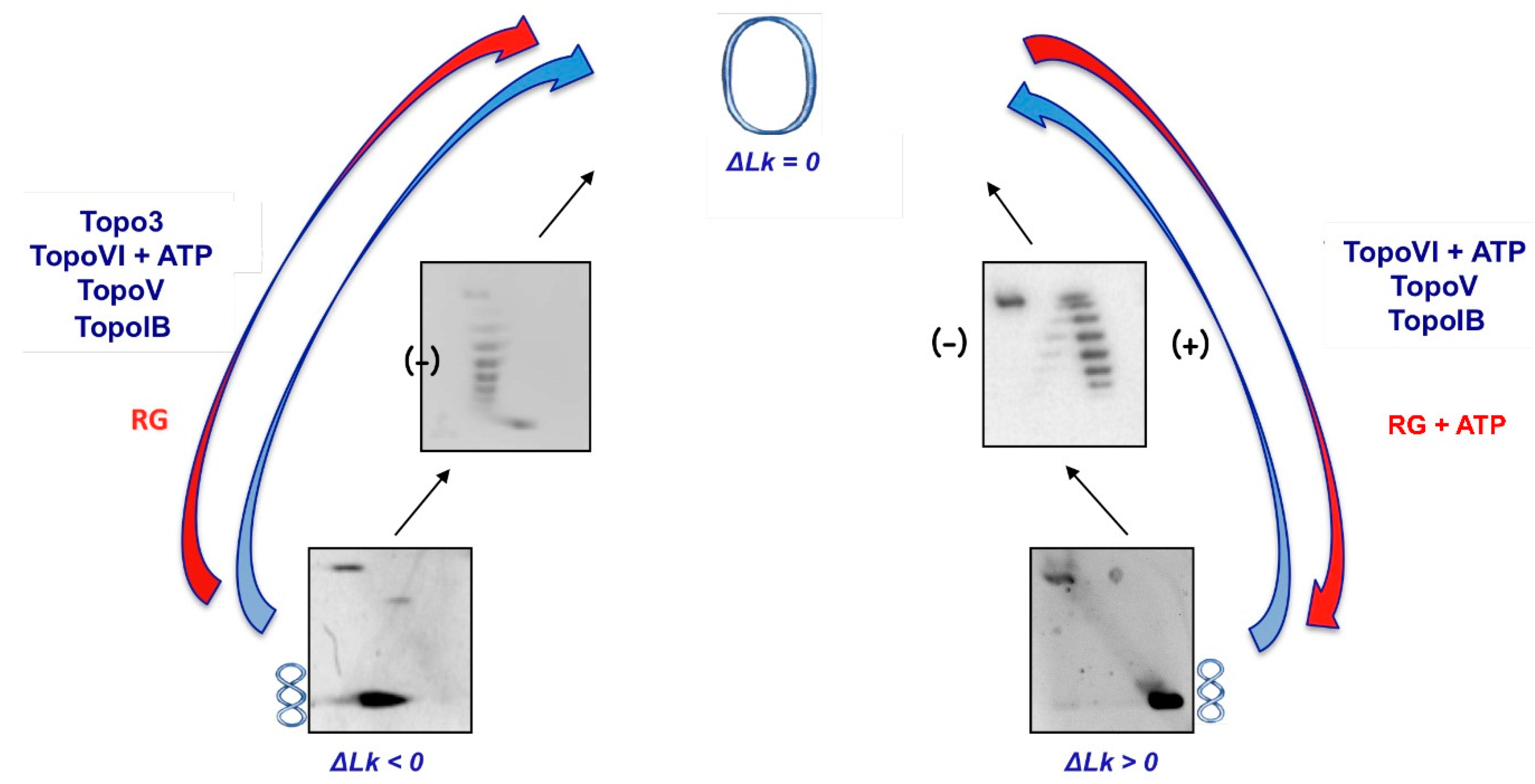
3. DNA Topoisomerases from Hyperthermophilic Archaea
| Enzyme | Topo VI | Topo 3 | Reverse Gyrase | Topo V |
|---|---|---|---|---|
| Type | II B | IA | IA | IC |
| Structure | Heterotetramer A2 + B2 (A domain containing Winged Helix Domain (Active Site)/B domain containing ATP Binding Site) | Monomer (C-terminal domain implicate in DNA binding/N-terminal domain implicate in topoisomerase activity) | Monomer (C-terminal domain like Topoisomerases type IA/N-terminal domain like SF2 helicases) | Monomer (N-terminal domain with topoisomerase activity/C-terminal domain with AP site processing activity) |
| Activity | ATP dependent positive and negative supercoiled DNA relaxation; Cut double strand | ATP independent negative supercoiled DNA relaxation; Cut single strand | ATP dependent positive supercoiled DNA; ATP independent negative supercoiled DNA relaxation; Cut single strand | Positive and negative supercoiled DNA relaxation; Cut single strand |

3.1. Topoisomerase VI
3.2. Topoisomerase 3
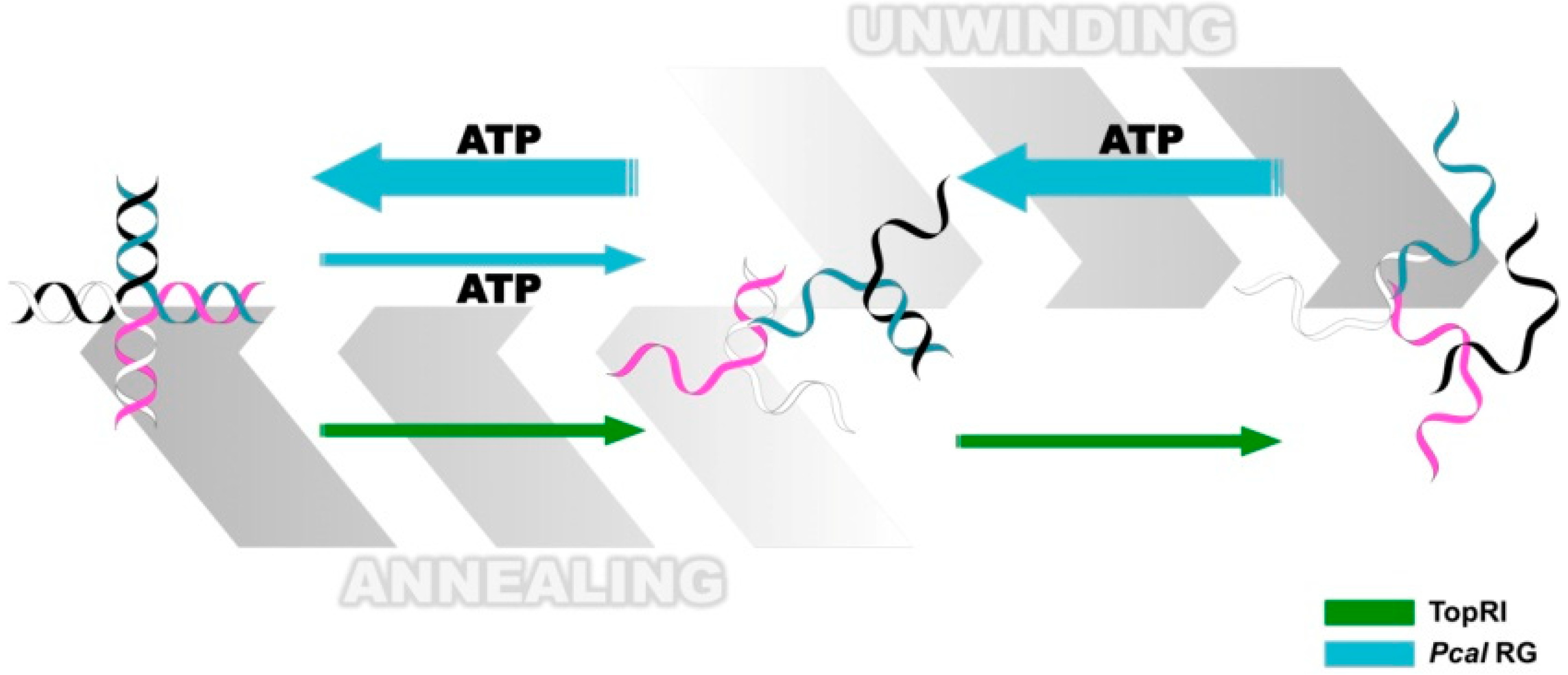
3.3. Reverse Gyrase

3.4. Topoisomerase V
3.5. Topoisomerase IB
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Luijsterburg, M.S.; White, M.F.; van Driel, R.; Dame, R.T. The major architects of chromatin: Architectural proteins in bacteria, archaea and eukaryotes. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 393–418. [Google Scholar]
- Champoux, J.J. DNA topoisomerases: Structure, function, and mechanism. Annu. Rev. Biochem. 2001, 70, 369–413. [Google Scholar]
- Wang, J.C. Cellular roles of DNA topoisomerases: A molecular perspective. Nat. Rev. Mol. Cell Biol. 2002, 3, 430–440. [Google Scholar]
- Chen, S.H.; Chan, N.L.; Hsieh, T.S. New mechanistic and functional insights into DNA topoisomerases. Annu. Rev. Biochem. 2013, 82, 139–170. [Google Scholar]
- Segal, E.; Fondufe-Mittendorf, Y.; Chen, L.; Thåström, A.; Field, Y.; Moore, I.K.; Wang, J.P.; Widom, J. A genomic code for nucleosome positioning. Nature 2006, 442, 772–778. [Google Scholar]
- Trifonov, E.N. Cracking the chromatin code: Precise rule of nucleosome positioning. Phys. Life Rev. 2011, 8, 39–50. [Google Scholar]
- Meyer, S.; Becker, N.B.; Syed, S.H.; Goutte-Gattat, D.; Shukla, M.S.; Hayes, J.J.; Angelov, D.; Bednar, J.; Dimitrov, S.; Everaers, R. From crystal and NMR structures, footprints and cryo-electron-micrographs to large and soft structures: Nanoscale modeling of the nucleosomal stem. Nucleic Acids Res. 2011, 39, 9139–9154. [Google Scholar]
- Kelly, T.K.; Liu, Y.; Lay, F.D.; Liang, G.; Berman, B.P.; Jones, P.A. Genome-wide mapping of nucleosome positioning and DNA methylation within individual DNA molecules. Genome Res. 2012, 22, 2497–2506. [Google Scholar]
- Xiao, B.; Freedman, B.S.; Miller, K.E.; Heald, R.; Marko, J.F. Histone H1 compacts DNA under force and during chromatin assembly. Mol. Biol. Cell. 2012, 23, 4864–4871. [Google Scholar]
- Forterre, P. Introduction and historical perspective. In DNA Topoisomerases and Cancer; Pommier, Yves, Ed.; National Institutes of Health: Bethesda, MD, USA, 2012; pp. 1–52. [Google Scholar]
- Sandman, K.; Reeve, J.N. Archaeal chromatin proteins: Different structures but common function? Curr. Opin. Microbiol. 2005, 8, 656–661. [Google Scholar]
- Zhang, Z.; Guo, L.; Huang, L. Archaeal chromatin proteins. China Life Sci. 2012, 55, 377–385. [Google Scholar]
- Driessen, R.P.; Dame, R.T. Nucleoid-associated proteins in Crenarchaea. Biochem. Soc. Trans. 2011, 39, 116–121. [Google Scholar]
- Reeve, J.N.; Bailey, K.A.; Li, W.T.; Marc, F.; Sandman, K.; Soares, D.J. Archaeal histones: Structures, stability and DNA binding. Biochem. Soc. Trans. 2004, 32, 227–230. [Google Scholar]
- Forterre, P.; Bergerat, A.; Lopez-Garcia, P. The unique DNA topology and DNA topoisomerases of hyperthermophilic archaea. FEMS Microbiol. Rev. 1996, 18, 237–248. [Google Scholar]
- López-García, P. DNA supercoiling and temperature adaptation: A clue to early diversification of life? J. Mol. Evol. 1999, 49, 439–452. [Google Scholar]
- DeLange, R.J.; Green, G.R.; Searcy, D.G. A histone-like protein (HTa) from Thermoplasma acidophilum. I. Purification and properties. J. Biol. Chem. 1981, 256, 900–904. [Google Scholar]
- Sandman, K.; Krzychi, J.A.; Dobrinski, B.; Lurz, R.; Reeve, J.N. HMf, a DNA-binding protein isolated from the hyperthermophilic archaeon Methanothermus fervidus, is most closely related to histones. Proc. Natl. Acad. Sci. USA 1990, 87, 5788–5791. [Google Scholar]
- Musgrave, D.R.; Sandman, K.M.; Reeve, J.N. DNA binding by the archaeal histone HMf results in positive supercoiling. Proc. Natl. Acad. Sci. USA 1991, 88, 10397–10401. [Google Scholar]
- Musgrave, D.; Forterre, P.; Slesarev, A. Negative constrained DNA supercoiling in archaeal nucleosomes. Mol. Microbiol. 2000, 35, 341–349. [Google Scholar]
- Marc, F.; Sandman, K.; Lurz, R.; Reeve, J.N. Archaeal histone tetramerization determines DNA affinity and the direction of DNA supercoiling. J. Biol. Chem. 2002, 277, 30879–30886. [Google Scholar]
- Starich, M.R.; Sandman, K.; Reeve, J.N.; Summers, M.F. NMR structure of HMfB from the hyperthermophile, Methanothermus fervidus, confirms that this archaeal protein is a histone. J. Mol. Biol. 1996, 255, 187–203. [Google Scholar]
- Fahrner, R.L.; Cascio, D.; Lake, J.A.; Slesarev, A. An ancestral nuclear protein assembly: Crystal structure of the Methanopyrus kandleri histone. Protein Sci. 2001, 10, 2002–2007. [Google Scholar]
- Sandman, K.; Grayling, R.A.; Dobrinski, B.; Lurz, R.; Reeve, J.N. Growth-phase-dependent synthesis of histones in the archaeon Methanothermus fervidus. Proc. Natl. Acad. Sci. USA 1994, 91, 12624–12628. [Google Scholar]
- Dinger, M.E.; Baillie, G.J.; Musgrave, D.R. Growth phase-dependent expression and degradation of histones in the thermophilic archaeon Thermococcus zilligii. Mol. Microbiol. 2000, 36, 876–885. [Google Scholar]
- Nalabothula, N.; Xi, L.; Bhattacharyya, S.; Widom, J.; Wang, J.P.; Reeve, J.N.; Santangelo, T.J.; Fondufe-Mittendorf, Y.N. Archaeal nucleosome positioning in vivo and in vitro is directed by primary sequence motifs. BMC Genomics 2013, 14. [Google Scholar] [CrossRef]
- Maruyama, H.; Harwood, J.C.; Moore, K.M.; Paszkiewicz, K.; Durley, S.C.; Fukushima, H.; Atomi, H.; Takeyasu, K.; Kent, N.A. An alternative beads-on-a-string chromatin architecture in Thermococcus kodakarensis. EMBO Rep. 2013, 14, 711–777. [Google Scholar]
- Soares, D.; Dahlke, I.; Li, W.T.; Sandman, K.; Hethke, C.; Thomm, M.; Reeve, J.N. Archaeal histone stability, DNA binding, and transcription inhibition above 90 degrees C. Extremophiles 1998, 2, 75–81. [Google Scholar]
- Xie, Y.; Reeve, J.N. Transcription by an archaeal RNA polymerase is slowed but not blocked by an archaeal nucleosome. J. Bacteriol. 2004, 186, 3492–3498. [Google Scholar]
- Xuan, J.; Feng, Y. The archaeal Sac10b protein family: Conserved proteins with divergent functions. Curr. Protein Pept. Sci. 2012, 13, 258–266. [Google Scholar]
- Bell, S.D.; Botting, C.H.; Wardleworth, B.N.; Jackson, S.P.; White, M.F. The interaction of Alba, a conserved archaeal chromatin protein, with Sir2 and its regulation by acetylation. Science 2002, 296, 148–151. [Google Scholar]
- Wardleworth, B.N.; Russell, R.J.; Bell, S.D.; Taylor, G.L.; White, M.F. Structure of Alba: An archaeal chromatin protein modulated by acetylation. EMBO J. 2002, 21, 4654–4662. [Google Scholar]
- Aravind, L.; Iyer, L.M.; Anantharaman, V. The two faces of Alba: The evolutionary connection between proteins participating in chromatin structure and RNA metabolism. Genome Biol. 2003, 4, R64:1–R64:9. [Google Scholar]
- Lurz, R.; Grote, M.; Dijk, J.; Reinhardt, R.; Dobrinski, B. Electron microscopic study of DNA complexes with proteins from the Archaebacterium Sulfolobus acidocaldarius. EMBO J. 1986, 5, 3715–3721. [Google Scholar]
- Xue, H.; Guo, R.; Wen, Y.; Liu, D.; Huang, L. An abundant DNA binding protein from the hyperthermophilic archaeon Sulfolobus shibatae affects DNA supercoiling in a temperature-dependent fashion. J. Bacteriol. 2000, 182, 3929–3933. [Google Scholar]
- Chen, L.; Chen, L.R.; Zhou, X.E.; Wang, Y.; Kahsai, M.A.; Clark, A.T.; Edmondson, S.P.; Liu, Z.J.; Rose, J.P.; Wang, B.C.; et al. The hyperthermophile protein Sso10a is a dimer of winged helix DNA-binding domains linked by an antiparallel coiled coil rod. J. Mol. Biol. 2004, 341, 73–91. [Google Scholar]
- Edmondson, S.P.; Kahsai, M.A.; Gupta, R.; Shriver, J.W. Characterization of Sac10a, a hyperthermophile DNA-binding protein from Sulfolobus acidocaldarius. Biochemistry 2004, 43, 13026–13036. [Google Scholar]
- Jelinska, C.; Conroy, M.J.; Craven, C.J.; Hounslow, A.M.; Bullough, P.A.; Waltho, J.P.; Taylor, G.L.; White, M.F. Obligate heterodimerization of the archaeal Alba2 protein with Alba1 provides a mechanism for control of DNA packaging. Structure 2005, 13, 963–971. [Google Scholar]
- Jelinska, C.; Petrovic-Stojanovska, B.; Ingledew, W.J.; White, M.F. Dimer–dimer stacking interactions are important for nucleic acid binding by the archaeal chromatin protein Alba. Biochem. J. 2010, 427, 49–55. [Google Scholar] [Green Version]
- Lu, Y.W.; Huang, T.; Tsai, C.T.; Chang, Y.Y.; Li, H.W.; Hsu, C.H.; Fan, H.F. Using single-molecule approaches to study archaeal DNA-binding protein Alba1. Biochemistry 2013, 52, 7714–7722. [Google Scholar]
- Laurens, N.; Driessen, R.P.; Heller, I.; Vorselen, D.; Noom, M.C.; Hol, F.J.; White, M.F.; Dame, R.T.; Wuite, G.J. Alba shapes the archaeal genome using a delicate balance of bridging and stiffening the DNA. Nat. Commun. 2012, 3. [Google Scholar] [CrossRef]
- Marsh, V.L.; Peak-Chew, S.Y.; Bell, S.D. Sir2 and the acetyltransferase, Pat, regulate the archaeal chromatin protein, Alba. J. Biol. Chem. 2005, 280, 21122–21128. [Google Scholar]
- Brent, M.M.; Iwata, A.; Carten, J.; Zhao, K.; Marmorstein, R. Structure and biochemical characterization of protein acetyltransferase from Sulfolobus solfataricus. J. Biol. Chem. 2009, 284, 19412–19419. [Google Scholar]
- Goyal, M.; Alam, A.; Iqbal, M.S.; Dey, S.; Bindu, S.; Pal, C.; Banerjee, A.; Chakrabarti, S.; Bandyopadhyay, U. Identification and molecular characterization of an Alba-family protein from human malaria parasite Plasmodium falciparum. Nucleic Acids Res. 2012, 40, 1174–1190. [Google Scholar]
- Chou, C.C.; Lin, T.W.; Chen, C.Y.; Wang, A.H. Crystal structure of the hyperthermophilic archaeal DNA-binding protein Sso10b2 at a resolution of 1.85 Angstroms. J. Bacteriol. 2003, 185, 4066–4073. [Google Scholar]
- Zhao, K.; Chai, X.; Marmorstein, R. Structure of a Sir2 substrate, Alba, reveals a mechanism for deacetylation-induced enhancement of DNA binding. J. Biol. Chem. 2003, 278, 26071–26077. [Google Scholar]
- Biyani, K.; Kahsai, M.A.; Clark, A.T.; Armstrong, T.L.; Edmondson, S.P.; Shriver, J.W. Solution structure, stability, and nucleic acid binding of the hyperthermophile protein Sso10b2. Biochemistry 2005, 44, 14217–14230. [Google Scholar]
- Kahsai, M.A.; Vogler, B.; Clark, A.T.; Edmondson, S.P.; Shriver, J.W. Solution structure, stability, and flexibility of Sso10a: A hyperthermophile coiled-coil DNA-binding protein. Biochemistry 2005, 44, 2822–2832. [Google Scholar]
- Tanaka, T.; Padavattan, S.; Kumarevel, T. Crystal structure of archaeal chromatin protein Alba2-double-stranded DNA complex from Aeropyrum pernix K1. J. Biol. Chem. 2012, 287, 10394–10402. [Google Scholar]
- Hada, K.; Nakashima, T.; Osawa, T.; Shimada, H.; Kakuta, Y.; Kimura, M. Crystal structure and functional analysis of an archaeal chromatin protein Alba from the hyperthermophilic archaeon Pyrococcus horikoshii OT3. Biosci. Biotechnol. Biochem. 2008, 72, 749–758. [Google Scholar]
- Guo, R.; Xue, H.; Huang, L. Ssh10b, a conserved thermophilic archaeal protein, binds RNA in vivo. Mol. Microbiol. 2003, 50, 1605–1615. [Google Scholar]
- Baumann, H.; Knapp, S.; Lundbäck, T.; Ladenstein, R.; Härd, T. Solution structure and DNA-binding properties of a thermostable protein from the archaeon Sulfolobus solfataricus. Nat. Struct. Biol. 1994, 1, 808–819. [Google Scholar]
- Guagliardi, A.; Napoli, A.; Rossi, M.; Ciaramella, M. Annealing of complementary DNA strands above the melting point of the duplex promoted by an archaeal protein. J. Mol. Biol. 1997, 267, 841–848. [Google Scholar]
- Agback, P.; Baumann, H.; Knapp, S.; Ladenstein, R.; Härd, T. Architecture of nonspecific protein–DNA interactions in the Sso7d-DNA complex. Nat. Struct. Biol. 1998, 5, 579–584. [Google Scholar]
- Edmondson, S.P.; Qiu, L.; Shriver, J.W. Solution structure of the DNA-binding protein Sac7d from the hyperthermophile Sulfolobus acidocaldarius. Biochemistry 1995, 34, 13289–13304. [Google Scholar]
- Gao, Y.G.; Su, S.Y.; Robinson, H.; Padmanabhan, S.; Lim, L.; McCrary, B.S.; Edmondson, S.P. The crystal structure of the hyperthermophile chromosomal protein Sso7d bound to DNA. Nat. Struct. Biol. 1998, 5, 782–786. [Google Scholar]
- Napoli, A.; Zivanovic, Y.; Bocs, C.; Buhler, C.; Rossi, M.; Forterre, P.; Ciaramella, M. DNA bending, compaction and negative supercoiling by the architectural protein Sso7d of Sulfolobus solfataricus. Nucleic Acids Res. 2002, 30, 2656–2662. [Google Scholar]
- Napoli, A.; Valenti, A.; Salerno, V.; Nadal, M.; Garnier, F.; Rossi, M.; Ciaramella, M. Functional interaction of reverse gyrase with single-strand binding protein of the archaeon Sulfolobus. Nucleic Acids Res. 2005, 33, 564–576. [Google Scholar]
- Salerno, V.; Napoli, A.; White, M.F.; Rossi, M.; Ciaramella, M. Transcriptional response to DNA damage in the archaeon Sulfolobus solfataricus. Nucleic Acids Res. 2003, 31, 6127–6138. [Google Scholar]
- Valenti, A.; Napoli, A.; Ferrara, M.C.; Nadal, M.; Rossi, M.; Ciaramella, M. Selective degradation of reverse gyrase and DNA fragmentation induced by alkylating agent in the archaeon Sulfolobus solfataricus. Nucleic Acids Res. 2006, 34, 2098–2108. [Google Scholar]
- Guo, L.; Feng, Y.; Zhang, Z.; Yao, H.; Luo, Y.; Wang, J.; Huang, L. Biochemical and structural characterization of Cren7, a novel chromatin protein conserved among Crenarchaea. Nucleic Acids Res. 2008, 36, 1129–1137. [Google Scholar]
- Chen, L.; Zhang, J.L.; Yu, L.Y.; Zheng, Q.C.; Chu, W.T.; Xue, Q.; Zhang, H.X.; Sun, C.C. Influence of hyperthermophilic protein Cren7 on the stability and conformation of DNA: Insights from molecular dynamics simulation and free energy analysis. J. Phys. Chem. B 2012, 116, 12415–12425. [Google Scholar]
- Feng, Y.; Yao, H.; Wang, J. Crystal structure of the crenarchaeal conserved chromatin protein Cren7 and double-stranded DNA complex. Protein Sci. 2010, 19, 1253–1257. [Google Scholar]
- Zhang, Z.; Gong, Y.; Guo, L.; Jiang, T.; Huang, L. Structural insights into the interaction of the crenarchaeal chromatin protein Cren7 with DNA. Mol. Microbiol. 2010, 76, 749–759. [Google Scholar]
- Driessen, R.P.; Meng, H.; Suresh, G.; Shahapure, R.; Lanzani, G.; Priyakumar, U.D.; White, M.F.; Schiessel, H.; van Noort, J.; Dame, R.T. Crenarchaeal chromatin proteins Cren7 and Sul7 compact DNA by inducing rigid bends. Nucleic Acids Res. 2013, 41, 196–205. [Google Scholar]
- Chu, Y.; Zhang, Z.; Wang, Q.; Luo, Y.; Huang, L. Identification and characterization of a highly conserved crenarchaeal protein lysine methyltransferase with broad substrate specificity. J. Bacteriol. 2012, 194, 6917–6926. [Google Scholar]
- Niu, Y.; Xia, Y.; Wang, S.; Li, J.; Niu, C.; Li, X.; Zhao, Y.; Xiong, H.; Li, Z.; Lou, H.; et al. A prototypic lysine methyltransferase 4 from archaea with degenerate sequence specificity methylates chromatin proteins Sul7d and Cren7 in different patterns. J. Biol. Chem. 2013, 288, 13728–13740. [Google Scholar]
- Sun, F.; Huang, L. Sulfolobus chromatin proteins modulate strand displacement by DNA polymerase B1. Nucleic Acids Res. 2013, 41, 8182–8195. [Google Scholar]
- Napoli, A.; Kvaratskelia, M.; White, M.F.; Rossi, M.; Ciaramella, M. A novel member of the bacterial-archaeal regulator family is a nonspecific DNA-binding protein and induces positive supercoiling. J. Biol. Chem. 2001, 276, 10745–10752. [Google Scholar]
- Napoli, A.; van der Oost, J.; Sensen, C.W.; Charlebois, R.L.; Rossi, M.; Ciaramella, M. An Lrp-likeprotein of the hyperthermophilic archaeon Sulfolobus solfataricus which binds to its own promoter. J. Bacteriol. 1999, 181, 1474–1480. [Google Scholar]
- Luo, X.; Schwarz-Linek, U.; Botting, C.H.; Hensel, R.; Siebers, B.; White, M.F. CC1, a novel crenarchaeal DNA binding protein. J. Bacteriol. 2007, 189, 403–409. [Google Scholar]
- Bergerat, A.; Gadelle, D.; Forterre, P. Purification of a DNA topoisomerase II from the hyperthermophilic archaeon Sulfolobus shibatae. A thermostable enzyme with both bacterial and eucaryal features. J. Biol. Chem. 1994, 269, 27663–27669. [Google Scholar]
- Bergerat, A.; de Massy, B.; Gadelle, D.; Varoutas, P.C.; Nicolas, A.; Forterre, P. An atypical topoisomerase II from Archaea with implications for meiotic recombination. Nature 1997, 386, 414–417. [Google Scholar]
- Buhler, C.; Lebbink, J.H.; Bocs, C.; Ladenstein, R.; Forterre, P. DNA topoisomerase VI generates ATP-dependent double-strand breaks with two-nucleotide overhangs. J. Biol. Chem. 2001, 276, 37215–37222. [Google Scholar]
- Dutta, R.; Inouye, M. GHKL, an emergent ATPase/kinase superfamily. Trends Biochem. Sci. 2000, 25, 24–28. [Google Scholar]
- Corbett, K.D.; Berger, J.M. Structure of the topoisomerase VI-B subunit: Implications for type II topoisomerase mechanism and evolution. EMBO J. 2003, 22, 151–163. [Google Scholar]
- Nichols, M.D.; de Angelis, K.; Keck, J.L.; Berger, J.M. Structure and function of an archeal topoisomerase VI subunit with homology to the meiotic recombination factor Spo11. EMBO J. 1999, 18, 6177–6188. [Google Scholar]
- Corbett, K.D.; Benedetti, P.; Berger, J.M. Holoenzyme assembly and ATP-mediated conformational dynamics of topoisomerase VI. Nat. Struct. Mol. Biol. 2007, 14, 611–619. [Google Scholar]
- Graille, M.; Cladière, L.; Durand, D.; Lecointe, F.; Gadelle, D.; Quevillon-Cheruel, S.; Vachette, P.; Forterre, P.; van Tilbeurgh, H. Crystal structure of an intact type II DNA topoisomerase: Insights into DNA transfer mechanisms. Structure 2008, 16, 360–370. [Google Scholar]
- Forterre, P.; Gribaldo, S.; Gadelle, D.; Serre, M.C. Origin and evolution of DNA topoisomerases. Biochimie 2007, 89, 427–446. [Google Scholar]
- Dai, P.; Wang, Y.; Ye, R.; Chen, L.; Huang, L. DNA topoisomerase III from the hyperthermophilic archaeon Sulfolobus solfataricus with specific DNA cleavage activity. J. Bacteriol. 2003, 185, 5500–5507. [Google Scholar]
- Chen, L.; Huang, L. Oligonucleotide cleavage and rejoining by topoisomerase III from the hyperthermophilic archaeon Sulfolobus solfataricus: Temperature dependence and strand annealing-promoted DNA religation. Mol. Microbiol. 2006, 60, 783–794. [Google Scholar]
- Li, X.; Guo, L.; Deng, L.; Feng, D.; Ren, Y.; Chu, Y.; She, Q.; Huang, L. Deletion of the topoisomerase III gene in the hyperthermophilic archaeon Sulfolobus islandicus results in slow growth and defects in cell cycle control. J. Genet. Genomics 2011, 38, 253–259. [Google Scholar]
- Lee, S.H.; Siaw, G.E.; Willcox, S.; Griffith, J.D.; Hsieh, T.S. Synthesis and dissolution of hemicatenanes by type IA DNA topoisomerases. Proc. Natl. Acad. Sci. USA 2013, 110, E3587–E3594. [Google Scholar]
- De Felice, M.; Aria, V.; Esposito, L.; de Falco, M.; Pucci, B.; Rossi, M.; Pisani, F.M. A novel DNA helicase with strand-annealing activity from the crenarchaeon Sulfolobus solfataricus. Biochem. J. 2007, 408, 87–95. [Google Scholar]
- Valenti, A.; de Felice, M.; Perugino, G.; Bizard, A.; Nadal, M.; Rossi, M.; Ciaramella, M. Synergic and opposing activities of thermophilic RecQ-like helicase and topoisomerase 3 proteins in Holliday junction processing and replication fork stabilization. J. Biol. Chem. 2012, 287, 30282–30295. [Google Scholar]
- D’Amaro, A.; Rossi, M.; Ciaramella, M. Reverse gyrase: An unusual DNA manipulator of hyperthermophilic organisms. Ital. J. Biochem. 2007, 56, 103–109. [Google Scholar]
- Nadal, M. Reverse gyrase: An insight into the role of DNA-topoisomerases. Biochimie 2007, 89, 447–455. [Google Scholar]
- Perugino, G.; Valenti, A.; D’Amaro, A.; Rossi, M.; Ciaramella, M. Reverse gyrase and genome stability in hyperthermophilic organisms. Biochem. Soc. Trans. 2009, 37, 69–73. [Google Scholar]
- Valenti, A.; Perugino, G.; Rossi, M.; Ciaramella, M. Positive supercoiling in thermophiles and mesophiles: Of the good and evil. Biochem. Soc. Trans. 2011, 39, 58–63. [Google Scholar]
- Lulchev, P.; Klostermeier, D. Reverse gyrase-recent advances and current mechanistic understanding of positive DNA supercoiling. Nucleic Acids Res. 2014, 42, 8200–8213. [Google Scholar]
- Vettone, A.; Perugino, G.; Rossi, M.; Valenti, A.; Ciaramella, M. Genome stability: Recent insights in the topoisomerase reverse gyrase and thermophilic DNA-alkyltransferase. Extremophiles 2014, 18, 895–904. [Google Scholar]
- Forterre, P. A hot story from comparative genomics: Reverse gyrase is the only hyperthermophile-specific protein. Trends Genet. 2002, 18, 236–237. [Google Scholar]
- Brochier-Armanet, C.; Forterre, P. Widespread distribution of archaeal reverse gyrase in thermophilic bacteria suggests a complex history of vertical inheritance and lateral gene transfers. Archaea 2007, 2, 83–93. [Google Scholar]
- Heine, M.; Chandra, S.B. The linkage between reverse gyrase and hyperthermophiles: A review of their invariable association. J. Microbiol. 2009, 47, 229–234. [Google Scholar]
- Kampmann, M.; Stock, D. Reverse gyrase has heat-protective DNA chaperone activity independent of supercoiling. Nucleic Acids Res. 2004, 32, 3537–3545. [Google Scholar]
- Atomi, H.; Fukui, T.; Kanai, T.; Morikawa, M.; Imanaka, T. Description of Thermococcus kodakaraensis sp. nov., a well studied hyperthermophilic archaeon previously reported as Pyrococcus sp. KOD1. Archaea 2004, 1, 263–267. [Google Scholar]
- Zhang, C.; Tian, B.; Li, S.; Ao, X.; Dalgaard, K.; Gökce, S.; Liang, Y.; She, Q. Genetic manipulation in Sulfolobus islandicus and functional analysis of DNA repair genes. Biochem. Soc. Trans. 2013, 41, 405–410. [Google Scholar]
- Rodríguez, A.C.; Stock, D. Crystal structure of reverse gyrase: Insights into the positive supercoiling of DNA. EMBO J. 2002, 21, 418–426. [Google Scholar]
- Rudolph, M.G.; del Toro Duany, Y.; Jungblut, S.P.; Ganguly, A.; Klostermeier, D. Crystal structures of Thermotoga maritima reverse gyrase: Inferences for the mechanism of positive DNA supercoiling. Nucleic Acids Res. 2013, 41, 1058–1070. [Google Scholar]
- Déclais, A.C.; Marsault, J.; Confalonieri, F.; de La Tour, C.B.; Duguet, M. Reverse gyrase, the two domains intimately cooperate to promote positive supercoiling. J. Biol. Chem. 2000, 275, 19498–19504. [Google Scholar]
- Valenti, A.; Perugino, G.; D’Amaro, A.; Cacace, A.; Napoli, A.; Rossi, M.; Ciaramella, M. Dissection of reverse gyrase activities: Insight into the evolution of a thermostable molecular machine. Nucleic Acids Res. 2008, 36, 4587–4597. [Google Scholar]
- Li, J.; Liu, J.; Zhou, J.; Xiang, H. Functional evaluation of four putative DNA-binding regions in Thermoanaerobacter tengcongensis reverse gyrase. Extremophiles 2011, 15, 281–291. [Google Scholar]
- Jamroze, A.; Perugino, G.; Valenti, A.; Rashid, N.; Rossi, M.; Akhtar, M.; Ciaramella, M. The reverse gyrase from Pyrobaculum calidifontis, a novel extremely thermophilic DNA topoisomerase endowed with DNA unwinding and annealing activities. J. Biol. Chem. 2014, 289, 3231–3243. [Google Scholar]
- Rodriguez, A.C. Studies of a positive supercoiling machine. Nucleotide hydrolysis and a multifunctional “latch” in the mechanism of reverse gyrase. J. Biol. Chem. 2002, 277, 29865–29873. [Google Scholar]
- Rodríguez, A.C. Investigating the role of the latch in the positive supercoiling mechanism of reverse gyrase. Biochemistry 2003, 42, 5993–6004. [Google Scholar]
- Bizard, A.; Garnier, F.; Nadal, M. TopR2, the second reverse gyrase of Sulfolobus solfataricus, exhibits unusual properties. J. Mol. Biol. 2011, 408, 839–849. [Google Scholar]
- Larsen, N.B.; Hickson, I.D. RecQ helicases: Conserved guardians of genomic integrity. Adv. Exp. Med. Biol. 2013, 767, 161–184. [Google Scholar]
- Plank, J.L.; Wu, J.; Hsieh, T.S. Topoisomerase IIIα and Bloom’s helicase can resolve a mobile double Holliday junction substrate through convergent branch migration. Proc. Natl. Acad. Sci. USA 2006, 103, 11118–11123. [Google Scholar]
- Bussen, W.; Raynard, S.; Busygina, V.; Singh, A.K.; Sung, P. Holliday junction processing activity of the BLM-Topo IIIα-BLAP75 complex. J. Biol. Chem. 2007, 282, 31484–31492. [Google Scholar]
- Valenti, A.; Ciaramella, M.; National Research Council of Italy, Naples, Italy. Unpublished work. 2009.
- Capp, C.; Qian, Y.; Sage, H.; Huber, H.; Hsieh, T.S. Separate and combined biochemical activities of the subunits of a naturally split reverse gyrase. J. Biol. Chem. 2010, 285, 39637–39645. [Google Scholar]
- Valenti, A.; Perugino, G.; Varriale, A.; D’Auria, S.; Rossi, M.; Ciaramella, M. The archaeal topoisomerase reverse gyrase is a helix-destabilizing protein that unwinds four-way DNA junctions. J. Biol. Chem. 2010, 285, 36532–36541. [Google Scholar]
- Ganguly, A.; del Toro Duany, Y.; Klostermeier, D. Reverse gyrase transiently unwinds double-stranded DNA in an ATP-dependent reaction. J. Mol. Biol. 2013, 425, 32–40. [Google Scholar]
- Jaxel, C.; Bouthier de la Tour, C.; Duguet, M.; Nadal, M. Reverse gyrase gene from Sulfolobus shibatae B12: Gene structure, transcription unit and comparative sequence analysis of the two domains. Nucleic Acids Res. 1996, 24, 4668–4675. [Google Scholar]
- Plank, J.; Hsieh, T.S. Helicase-appended topoisomerases: New insight into the mechanism of directional strand transfer. J. Biol. Chem. 2009, 284, 30737–30741. [Google Scholar]
- Napoli, A.; Valenti, A.; Salerno, V.; Nadal, M.; Garnier, F.; Rossi, M.; Ciaramella, M. Reverse gyrase recruitment to DNA after UV light irradiation in Sulfolobus solfataricus. J. Biol. Chem. 2004, 279, 33192–33198. [Google Scholar]
- Valenti, A.; Perugino, G.; Nohmi, T.; Rossi, M.; Ciaramella, M. Inhibition of translesion DNA polymerase by archaeal reverse gyrase. Nucleic Acids Res. 2009, 37, 4287–4295. [Google Scholar]
- Slesarev, A.I.; Stetter, K.O.; Lake, J.A.; Gellert, M.; Krah, R.; Kozyavkin, S.A. DNA topoisomerase V is a relative of eukaryotic topoisomerase I from a hyperthermophilic prokaryote. Nature 1993, 364, 735–737. [Google Scholar]
- Kozyavkin, S.A.; Pushkin, A.V.; Eiserling, F.A.; Stetter, K.O.; Lake, J.A.; Slesarev, A.I. DNA enzymology above 100 degrees C. Topoisomerase V unlinks circular DNA at 80–122 degrees C. J. Biol. Chem. 1995, 270, 13593–13595. [Google Scholar]
- Belova, G.I.; Prasad, R.; Nazimov, I.V.; Wilson, S.H.; Slesarev, A.I. The domain organization and properties of individual domains of DNA topoisomerase V, a type 1B topoisomerase with DNA repair activities. J. Biol. Chem. 2002, 277, 4959–4965. [Google Scholar]
- Pavlov, A.R.; Belova, G.I.; Kozyavkin, S.A.; Slesarev, A.I. Helix-hairpin-helix motifs confer salt resistance and processivity on chimeric DNA polymerases. Proc. Natl. Acad. Sci. USA 2002, 99, 13510–13515. [Google Scholar]
- Forterre, P. DNA topoisomerase V: A new fold of mysterious origin. Trends Biotechnol. 2006, 24, 245–247. [Google Scholar]
- Taneja, B.; Patel, A.; Slesarev, A.; Mondragón, A. Structure of the N-terminal fragment of topoisomerase V reveals a new family of topoisomerases. EMBO J. 2006, 25, 398–408. [Google Scholar]
- Taneja, B.; Schnurr, B.; Slesarev, A.; Marko, J.F.; Mondragón, A. Topoisomerase V relaxes supercoiled DNA by a constrained swiveling mechanism. Proc. Natl. Acad. Sci. USA 2007, 104, 14670–14675. [Google Scholar]
- Rajan, R.; Prasad, R.; Taneja, B.; Wilson, S.H.; Mondragón, A. Identification of one of the apurinic/apyrimidinic lyase active sites of topoisomerase V by structural and functional studies. Nucleic Acids Res. 2013, 41, 657–666. [Google Scholar]
- rochier-Armanet, C.; Gribaldo, S.; Forterre, P. A DNA topoisomerase IB in Thaumarchaeota testifies for the presence of this enzyme in the last common ancestor of Archaea and Eucarya. Biol. Direct. 2008, 3. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Mazzanti, M.; Mistrik, M.; Kosar, M.; Beznoussenko, G.V.; Mironov, A.A.; Garrè, M.; Parazzoli, D.; Shivashankar, G.V.; Scita, G.; et al. ATR mediates a checkpoint at the nuclear envelope in response to mechanical stress. Cell 2014, 158, 633–646. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Visone, V.; Vettone, A.; Serpe, M.; Valenti, A.; Perugino, G.; Rossi, M.; Ciaramella, M. Chromatin Structure and Dynamics in Hot Environments: Architectural Proteins and DNA Topoisomerases of Thermophilic Archaea. Int. J. Mol. Sci. 2014, 15, 17162-17187. https://doi.org/10.3390/ijms150917162
Visone V, Vettone A, Serpe M, Valenti A, Perugino G, Rossi M, Ciaramella M. Chromatin Structure and Dynamics in Hot Environments: Architectural Proteins and DNA Topoisomerases of Thermophilic Archaea. International Journal of Molecular Sciences. 2014; 15(9):17162-17187. https://doi.org/10.3390/ijms150917162
Chicago/Turabian StyleVisone, Valeria, Antonella Vettone, Mario Serpe, Anna Valenti, Giuseppe Perugino, Mosè Rossi, and Maria Ciaramella. 2014. "Chromatin Structure and Dynamics in Hot Environments: Architectural Proteins and DNA Topoisomerases of Thermophilic Archaea" International Journal of Molecular Sciences 15, no. 9: 17162-17187. https://doi.org/10.3390/ijms150917162