
Isolation and Expression of NAC Genes during Persimmon Fruit Postharvest Astringency Removal


Abstract
:1. Introduction
2. Results and Discussion
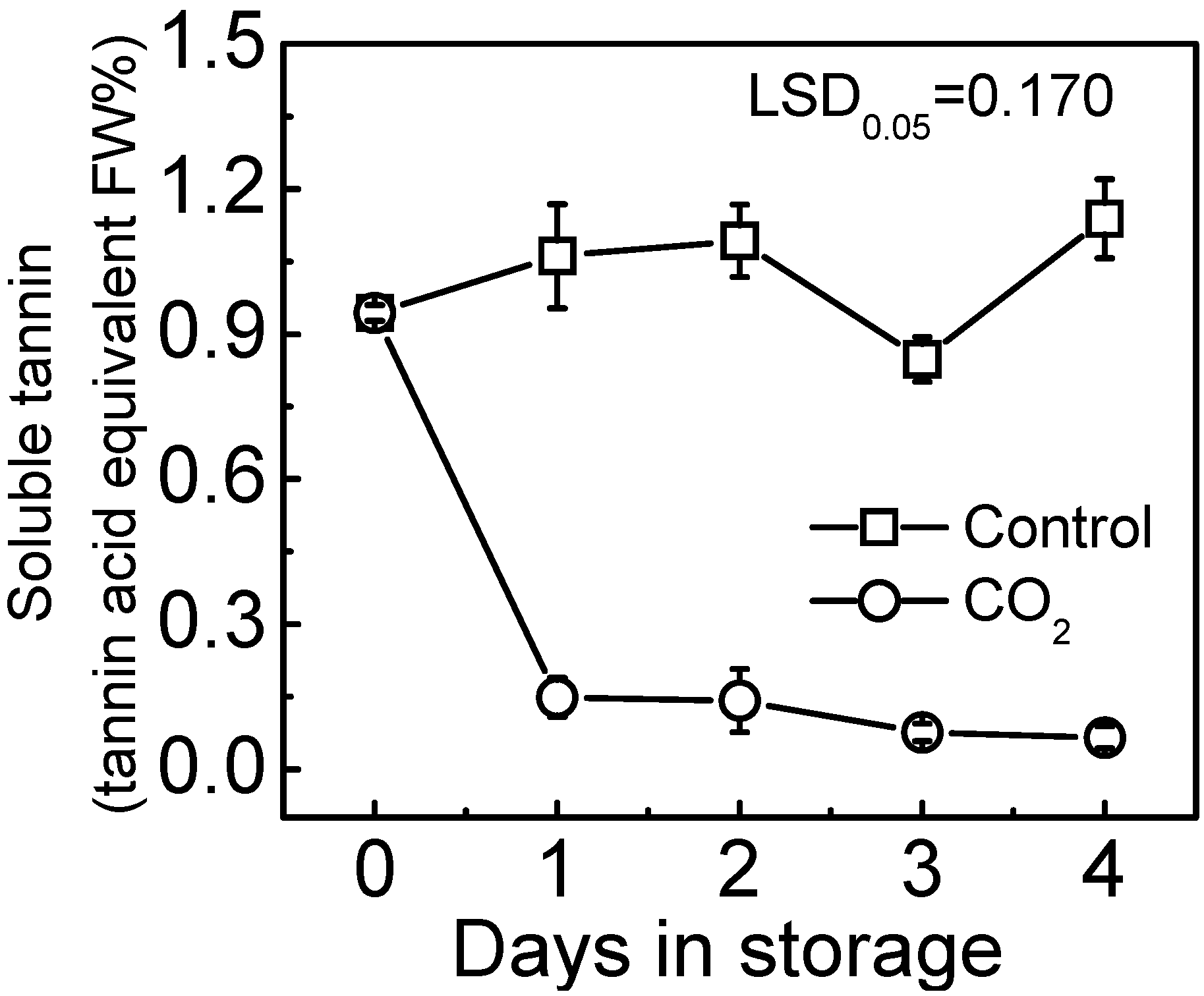
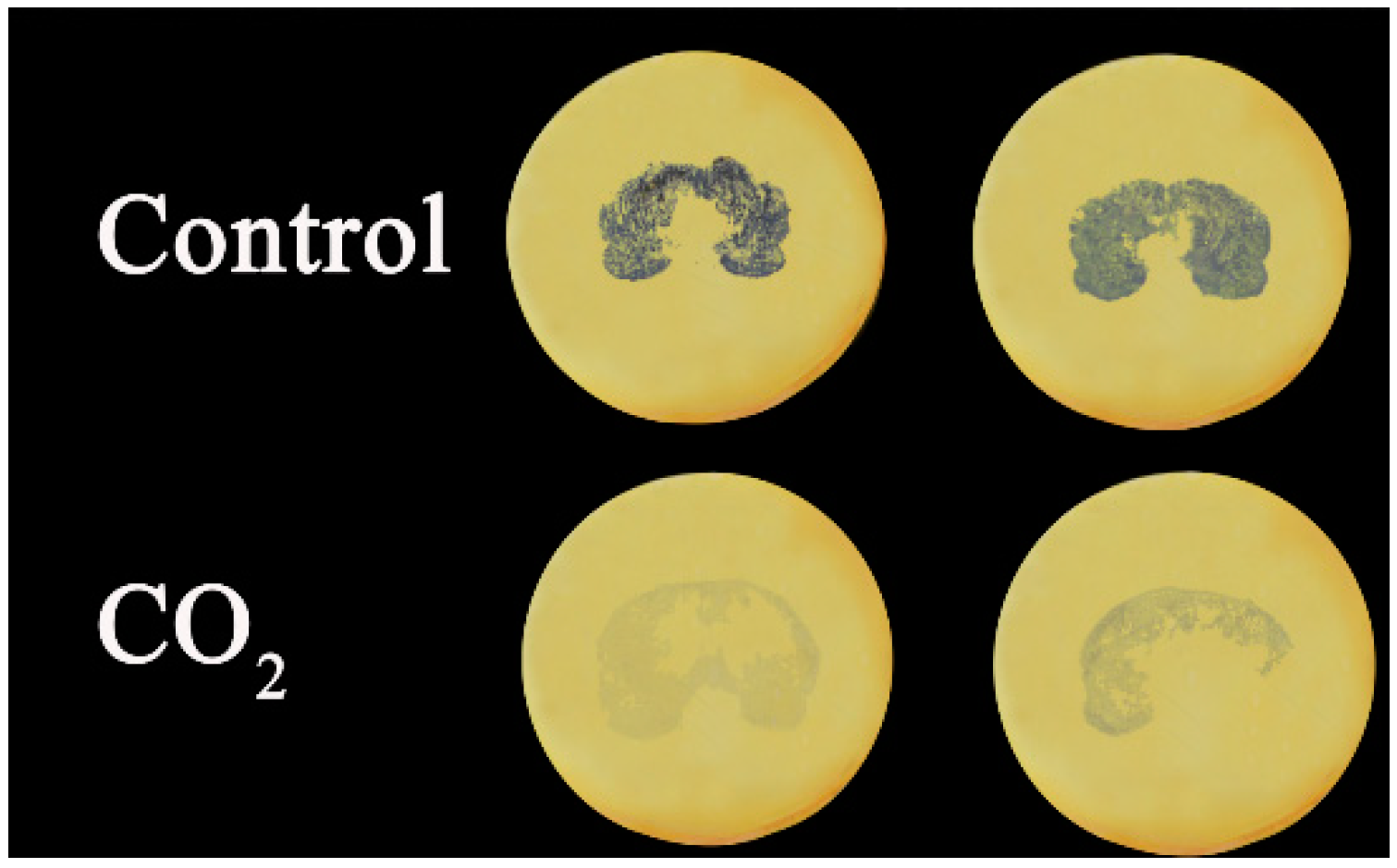
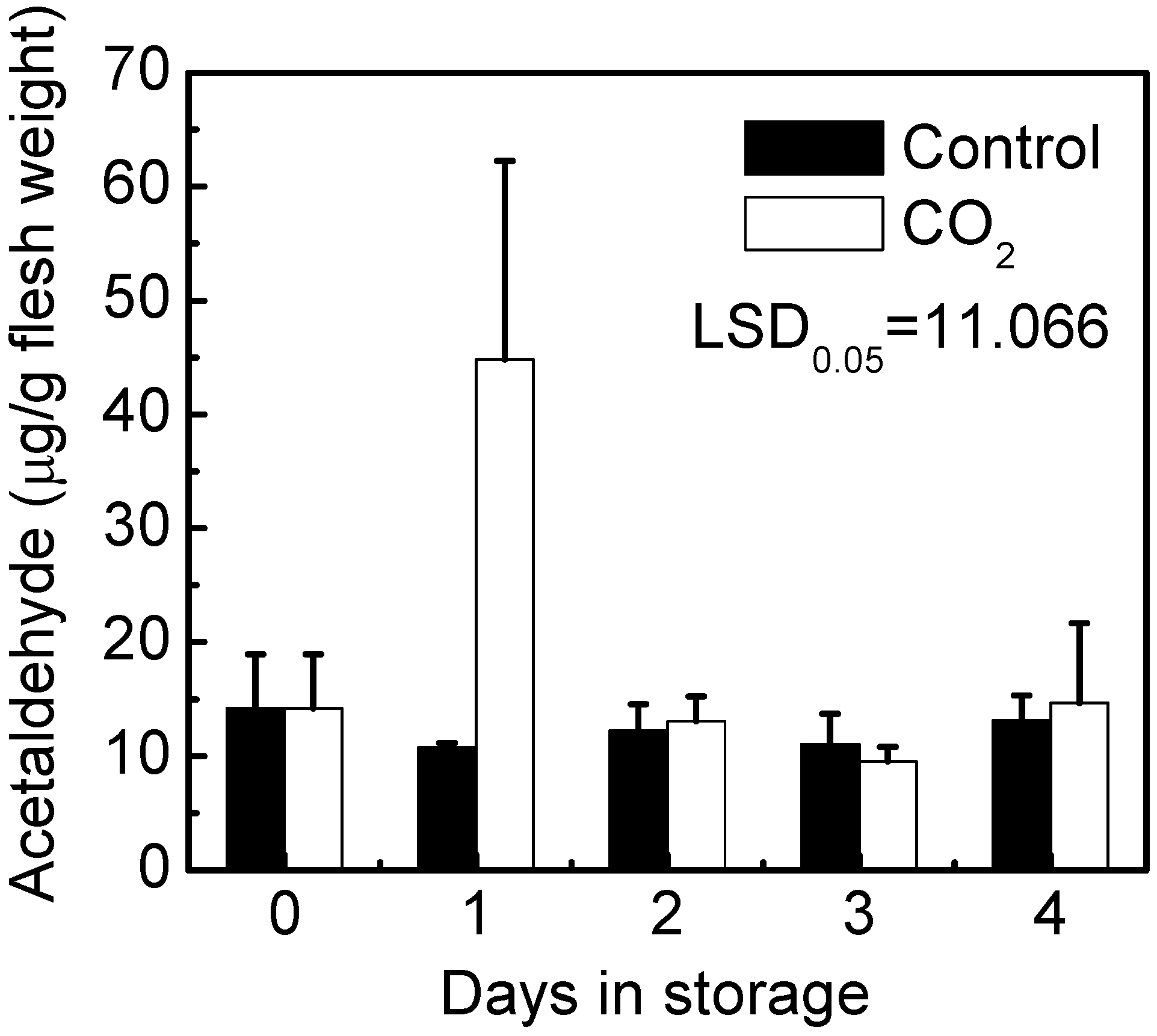
2.1. Fruit Deastringency




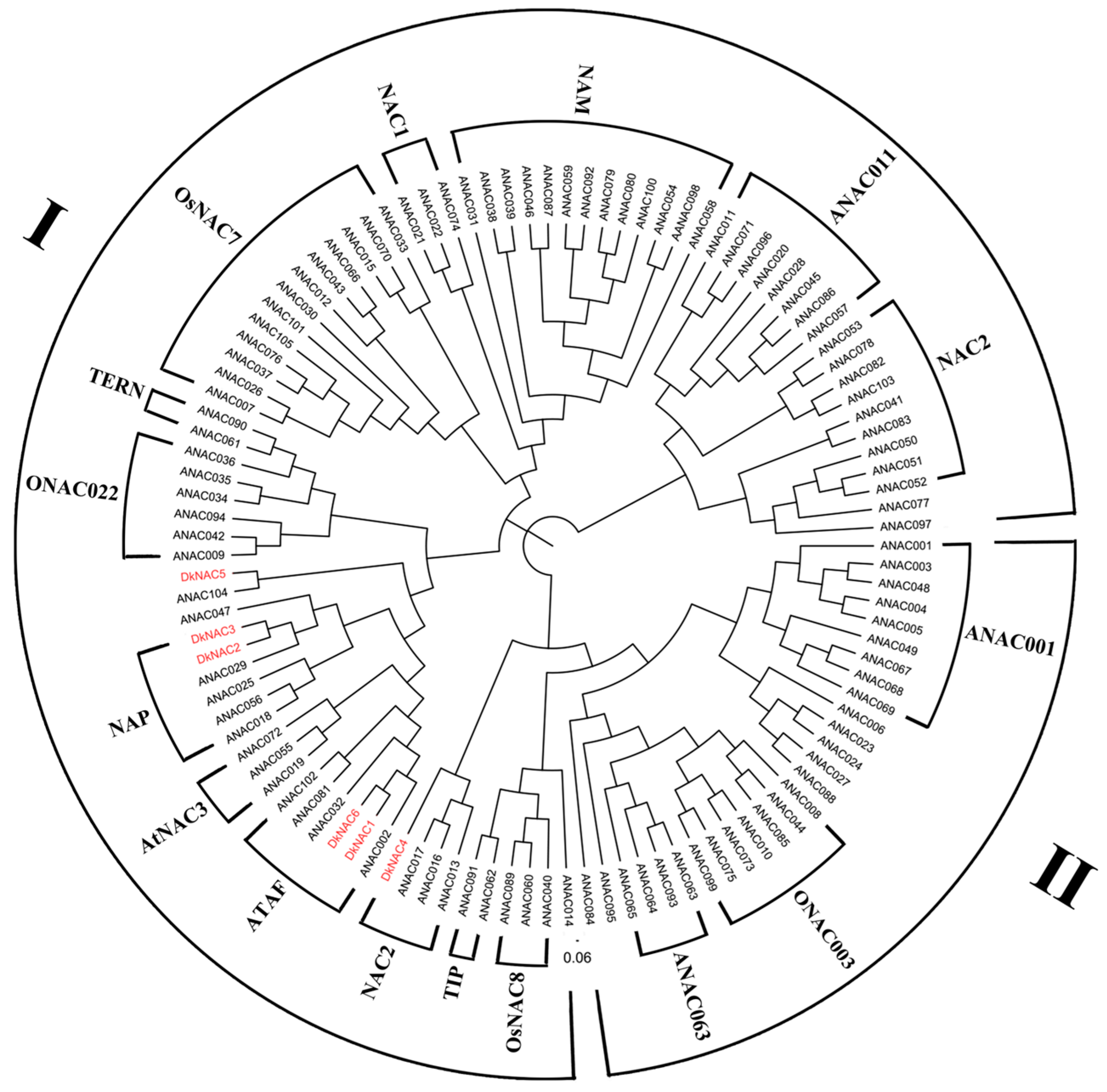
2.2. NAC Genes Isolation and Sequence Analysis

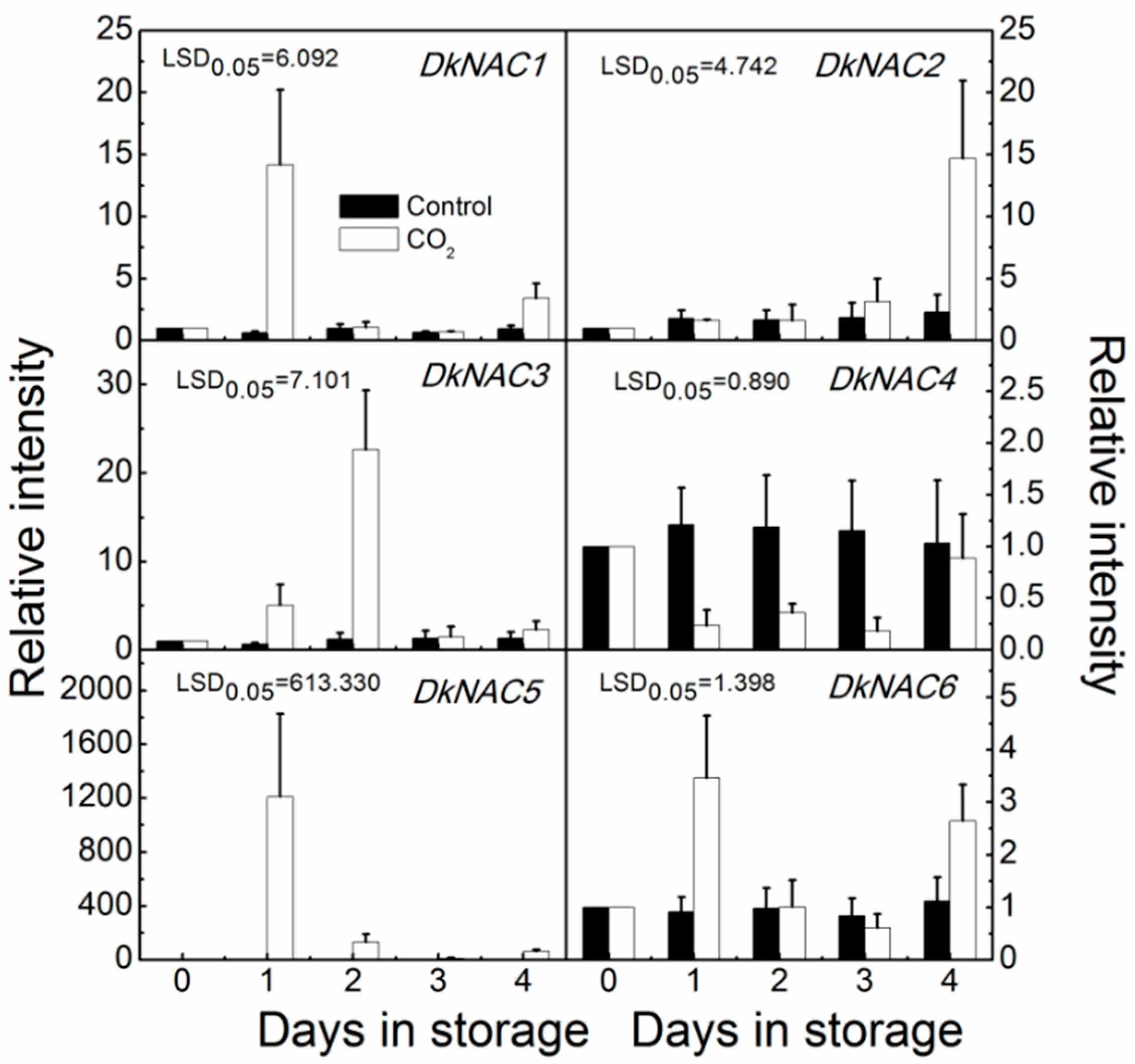
2.3. NAC Genes Expression

3. Experimental Section
3.1. Materials and Treatments
3.2. Soluble Tannins Assay and Tannin Printing
3.3. Acetaldehyde Determination
3.4. RNA Extraction and cDNA Synthesis
3.5. Gene Isolation and Sequence Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method Used | Gene | Primary PCR (5' to 3') | Secondary PCR (5' to 3') |
|---|---|---|---|
| 3'RACE | DkNAC1 | CAAGTTTGATCCGTGGCAACTTCCAG | TGGCTCTGTATGGAGAGAAAGAATGG |
| DkNAC2 | CCCAAGAGACCGGAAGTACCCGAAC | GGCGTCAAGAAAGCCCTCGTCTTCT | |
| DkNAC3 | AGAAGATTCAAGCGCCCAGATGGTG | TACCAGGACCCAGTTTGGAATGCAG | |
| DkNAC4 | CAAGAGCAGAAGTCACAAACAACTA | GAGCTGGAAGAGCTGGTTTTACC | |
| DkNAC5 | TCACGCTATCTCGGTCTTCGCTCTC | GGAGCTCATCGTCCATTTCCTCCAC | |
| DkNAC6 | AGTCCGGTGAGAGCGATATGATAG | GTGCCGAAGCTCCACACGGACTCG | |
| 5'RACE | DkNAC3 | TCCTGCATTCCAAACTGGGTCCTGGT | TGTCTTGATACCCTTTGGGGGCTTTCC |
| DkNAC4 | CCCAATCTGCTCGTTTCCCGTTTGG | CAACCCAACGATTTCCCACCCATAG | |
| DkNAC5 | CGTTCCCCTCCGCCATAGCTTTTCCA | GAGGGCAGCTTTGCGGTGGAGGAAA | |
| Full clone | DkNAC1 | CAGAGAGAGAGAGCTTGATCAGTG | CGCACCCGATTGAGAAAAT |
| DkNAC2 | TATAAATGGTGGCCGGAAAT | TCCGCACCTCGTACCTATCT | |
| DkNAC3 | CAAGAAAATGGCAGGAAGAAA | GCTGATTCACTGAAACCCATT | |
| DkNAC4 | CCGTCGGTACATGGAGAGAA | TGTCACATAGCCTGCTATAAAGAAGTA | |
| DkNAC5 | ACGCAGAGTACATGGGGAAT | ACGGAGAAGCCTGCATAGC |
3.6. Oligonucleotide Primers and Real-Time PCR
| Gene | Primary PCR (5' to 3') | Secondary PCR (5' to 3') |
|---|---|---|
| DkNAC1 | GGTAGCATCATAAGCGTTAATCTG | CAAGAATGACCCTATTACTACCACT |
| DkNAC2 | GGTGCGGATCGTAGAAACTA | ACAATTTTTGGGCCATAGGT |
| DkNAC3 | GATGTGGCTTGTTAGGCTTGA | CCCACAACAATACACGTTTGTTTCA |
| DkNAC4 | GCTGTATCTTTCTTGCATTGTTGAC | CAAGGGAGGAATGCCATGTA |
| DkNAC5 | GCTAGCTATGCAGGCTTCTCC | ACAAACAGCGCAACTCATTT |
| DkNAC6 | ACGCAGAGTACATGGGGAAT | ACGGAGAAGCCTGCATAGC |
3.7. Statistical Analysis
4. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Ahmad, N.; Gupta, S.; Mukhtar, H. Green tea polyphenol epigallocatechin-3-gallate differentially modulates nuclear factor κB in cancer cells versus normal cells. Arch. Biochem. Biophys. 2000, 76, 338–346. [Google Scholar] [CrossRef]
- Pataki, T.; Bak, I.; Kovacs, P.; Bagchi, D.; Das, K.; Tosaki, A. Grape seed proanthocyanidins improved cardiac recovery during reperfusion after ischemia in isolated rat hearts. Am. J. Clin. Nutr. 2002, 75, 894–895. [Google Scholar] [PubMed]
- Serafini, M.; Bugianesi, R.; Maiani, G.; Valtuena, S.; de Santis, S.; Crozier, A. Plasma antioxidants from chocolate. Nature 2003, 424, 1013. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.F.; Liang, J.; Li, C.M.; Zhang, J.; Dou, H.L. Study on inhibitory effects of persimmon tannins on activities of major enzymes from several snake venoms and its mechanism. Sci. Agric. Sin. 2008, 41, 910–917. [Google Scholar]
- Inoue, K.; Kawakita, H.; Ohio, K.; Oshima, T. Adsorptive removal of uranium and thorium with a crosslinked persimmon peel gel. J. Radioanal. Nucl. Chem. 2006, 267, 435–442. [Google Scholar] [CrossRef]
- Parajuli, D.; Kawakita, H.; Inoue, K.; Ohto, K.; Kajiyama, K. Persimmon peel gel for the selective recovery of gold. Hydrometallurgy 2007, 87, 133–139. [Google Scholar] [CrossRef]
- Xiong, Y.; Adhikari, C.R.; Kawakita, H.; Ohto, K.; Inoue, K.; Harada, H. Selective recovery of precious metals by persimmon waste chemically modified with dimethylamine. Bioresour. Technol. 2009, 100, 4083–4089. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Zhang, Q.L.; Luo, Z.R. Genome-wide transcriptome analysis of Chinese pollination-constant nonastringent persimmon fruit treated with ethanol. BMC Genomics 2014, 15, 112. [Google Scholar] [CrossRef] [PubMed]
- Akagi., T.; Ikegami, A.; Tsujimoto, T.; Kobayashi, S.; Sato, A.; Kono, A.; Yonemori, K. DkMyb4 is a Myb transcription factor involved in proanthocyanidin biosynthesis in persimmon fruit. Plant Physiol. 2009, 151, 2028–2045. [Google Scholar]
- Yonemori, K.; Suzuki, Y. Differences in three-dimensional distribution of tannin cells in flesh tissue between astringent and non-astringent type persimmon. Acta Hortic. 2008, 833, 119–124. [Google Scholar]
- Arnal, L.; Del-Río, M.A. Removing astringency by carbon dioxide and nitrogen-enriched atmospheres in persimmon fruit cv. “Rojo brillante”. J. Food Sci. 2003, 68, 1516–1518. [Google Scholar] [CrossRef]
- Salvador, A.; Arnal, L.; Besada, C.; Larrea, V.; Quiles, A.; Pérez-Munuera, I. Physiological and structural changes during ripening and deastringency treatment of persimmon fruit cv. “Rojo Brillante”. Postharvest Biol. Technol. 2007, 46, 181–188. [Google Scholar] [CrossRef]
- Yamada, M.; Taira, S.; Ohtsuki, M.; Sato, A.; Iwanami, H.; Yakushiji, H.; Wang, R.Z.; Yang, Y.; Li, G.C. Varietal differences in the ease of astringency removal by carbon dioxide gas and ethanol vapor treatments among oriental astringent persimmons of Japanese and Chinese origin. Sci. Hortic. 2002, 94, 63–72. [Google Scholar] [CrossRef]
- Yin, X.R.; Shi, Y.N.; Min, T.; Luo, Z.R.; Yao, Y.C.; Xu, Q.; Ferguson, I.B.; Chen, K.S. Expression of ethylene response genes during persimmon fruit astringency removal. Planta 2012, 235, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Taira, S.; Ono, M.; Otsuki, M. Effects of freezing rate on astringency reduction in persimmon during and after thawing. Postharvest Biol. Technol. 1998, 14, 317–324. [Google Scholar] [CrossRef]
- Matsuo, T.; Ito, S. On mechanisms of removing astringency in persimmon fruits by carbon dioxide treatment I. Some properties of the two processes in the de-astringency. Plant Cell Physiol. 1997, 18, 17–25. [Google Scholar]
- Tamura, F.; Tanabe, K.; Itai, A.; Hasegawa, M. Characteristics of acetaldehyde accumulation and removal of astringency with ethanol and carbon dioxide treatments in “Saijo” persimmon fruit. J. Jpn. Soc. Hortic. Sci. 1999, 68, 1178–1183. [Google Scholar] [CrossRef]
- Min, T.; Yin, X.R.; Shi, Y.N.; Luo, Z.R.; Yao, Y.C.; Grierson, D.; Ferguson, I.B.; Chen, K.S. Ethylene-responsive transcription factors interact with promoters of ADH and PDC involved in persimmon (Diospyros kaki) fruit de-astringency. J. Exp. Bot. 2012, 63, 6393–6405. [Google Scholar] [CrossRef] [PubMed]
- Christianson, J.A.; Wilson, I.W.; Llewellyn, D.J.; Dennis, E.S. The low-oxygen-induced NAC domain transcription factor ANAC102 affects viability of Arabidopsis seeds following low-oxygen treatment. Plant Physiol. 2009, 149, 1724–1738. [Google Scholar] [CrossRef] [PubMed]
- Min, T.; Fang, F.; Ge, H.; Shi, Y.N.; Luo, Z.R.; Yao, Y.C.; Grierson, D.; Yin, X.R.; Chen, K.S. Two novel anoxia-induced ethylene response factors that interact with promoters of deastringency-related genes from persimmon. PLoS One 2014, e97043. [Google Scholar] [CrossRef]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Nuruzzaman, M.; Manimekalai, R.; Sharon, A.M.; Satoh, K.; Kondoh, H.; Ooka, H.; Kikuchi, S. Genome-wide analysis of NAC transcription factor family in rice. Gene 2010, 465, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.S.P.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [PubMed]
- He, X.J.; Mu, R.L.; Cao, W.H.; Zhang, Z.G.; Zhang, J.S.; Chen, S.Y. AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development. Plant J. 2005, 44, 903–916. [Google Scholar] [CrossRef] [PubMed]
- Salvador, A.; Cuquerella, J.; Martínez-Jávega, J.M.; Monterde, A.; Navarro, P. 1-MCP preserves the firmness of stored persimmon “Rojo Brillante”. J. Food Sci. 2004, 69, 69–73. [Google Scholar]
- Hribar, J.; Zavrtanik, M.; Simčič, M.; Vidrih, R. Changes during storing and astringency removal of persimmon fruit (Diospyros Kaki L.). Acta Aliment. 2000, 29, 123–136. [Google Scholar] [CrossRef]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.F.; Gan, S.S. AtNAP, a NAC family transcription factor, has an important role in leaf senescence. Plant J. 2006, 46, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Clercq, I.D.; Vermeirssen, V.; van Aken, O.; Vandepoele, K.; Murcha, M.W.; Law, S.R.; Inzé, A.; Ng, S.; Ivanova, A.; Rombaut, D.; et al. The membrane-bound NAC transcription factor ANAC013 functions in mitochondrial retrograde regulation of the oxidative stress response in Arabidopsis. Plant Cell 2013, 25, 3472–3490. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Cai, Z.; Gan, S. Transcriptome of Arabidopsis leaf senescence. Plant Cell Environ. 2004, 27, 521–549. [Google Scholar] [CrossRef]
- Shan, W.; Kuang, J.F.; Chen, L.; Xie, H.; Peng, H.H.; Xiao, Y.Y.; Li, X.P.; Chen, W.X.; He, Q.G.; Chen, J.Y.; et al. Molecular characterization of banana NAC transcription factors and their interactions with ethylene signalling component EIL during fruit ripening. J. Exp. Bot. 2012, 63, 5171–5187. [Google Scholar] [CrossRef] [PubMed]
- Kou, X.H.; Watkins, C.B.; Gan, S.-S. Arabidopsis AtNAP regulates fruit senescence. J. Exp. Bot. 2012, 63, 6139–6147. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Luo, C.; Zhang, Q.L.; Luo, Z.R. Isolation and characterization of a laccase gene potentially involved in proanthocyanidin polymerization in oriental persimmon (Diospyros kaki Thunb.) fruit. Mol. Biol. Rep. 2013, 40, 2809–2820. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The ClustalX windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 24, 4876–4882. [Google Scholar] [CrossRef]
- Yin, X.R.; Chen, K.S.; Allan, A.C.; Wu, R.M.; Zhang, B.; Lallu, N.; Ferguson, I.B. Ethylene-induced modulation of genes associated with the ethylene signalling pathway in ripening kiwifruit. J. Exp. Bot. 2008, 59, 2097–2108. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Min, T.; Wang, M.-M.; Wang, H.; Liu, X.; Fang, F.; Grierson, D.; Yin, X.-R.; Chen, K.-S. Isolation and Expression of NAC Genes during Persimmon Fruit Postharvest Astringency Removal. Int. J. Mol. Sci. 2015, 16, 1894-1906. https://doi.org/10.3390/ijms16011894
Min T, Wang M-M, Wang H, Liu X, Fang F, Grierson D, Yin X-R, Chen K-S. Isolation and Expression of NAC Genes during Persimmon Fruit Postharvest Astringency Removal. International Journal of Molecular Sciences. 2015; 16(1):1894-1906. https://doi.org/10.3390/ijms16011894
Chicago/Turabian StyleMin, Ting, Miao-Miao Wang, Hongxun Wang, Xiaofen Liu, Fang Fang, Donald Grierson, Xue-Ren Yin, and Kun-Song Chen. 2015. "Isolation and Expression of NAC Genes during Persimmon Fruit Postharvest Astringency Removal" International Journal of Molecular Sciences 16, no. 1: 1894-1906. https://doi.org/10.3390/ijms16011894
APA StyleMin, T., Wang, M. -M., Wang, H., Liu, X., Fang, F., Grierson, D., Yin, X. -R., & Chen, K. -S. (2015). Isolation and Expression of NAC Genes during Persimmon Fruit Postharvest Astringency Removal. International Journal of Molecular Sciences, 16(1), 1894-1906. https://doi.org/10.3390/ijms16011894