
The Regulation of Nitric Oxide Synthase Isoform Expression in Mouse and Human Fallopian Tubes: Potential Insights for Ectopic Pregnancy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
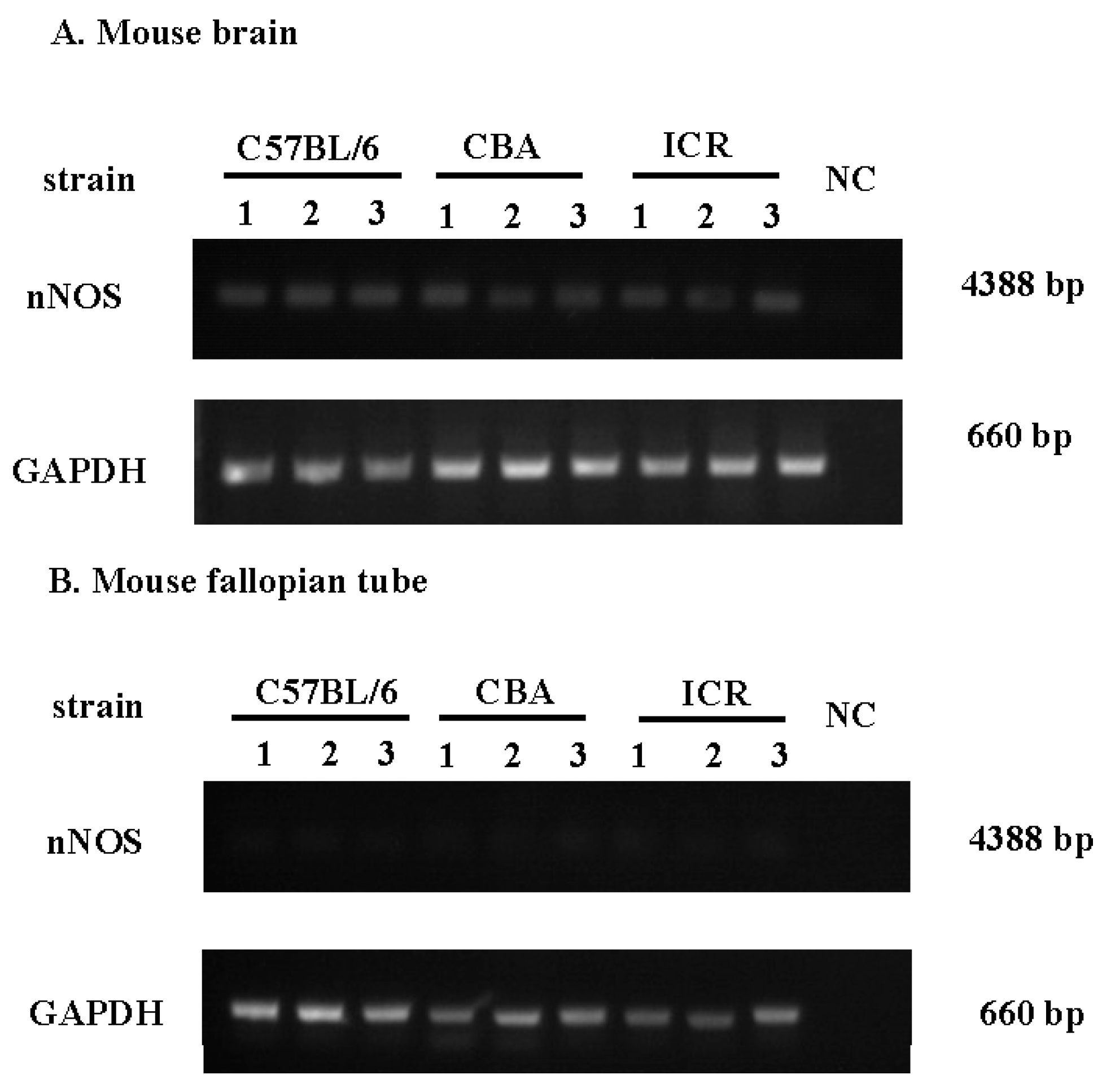
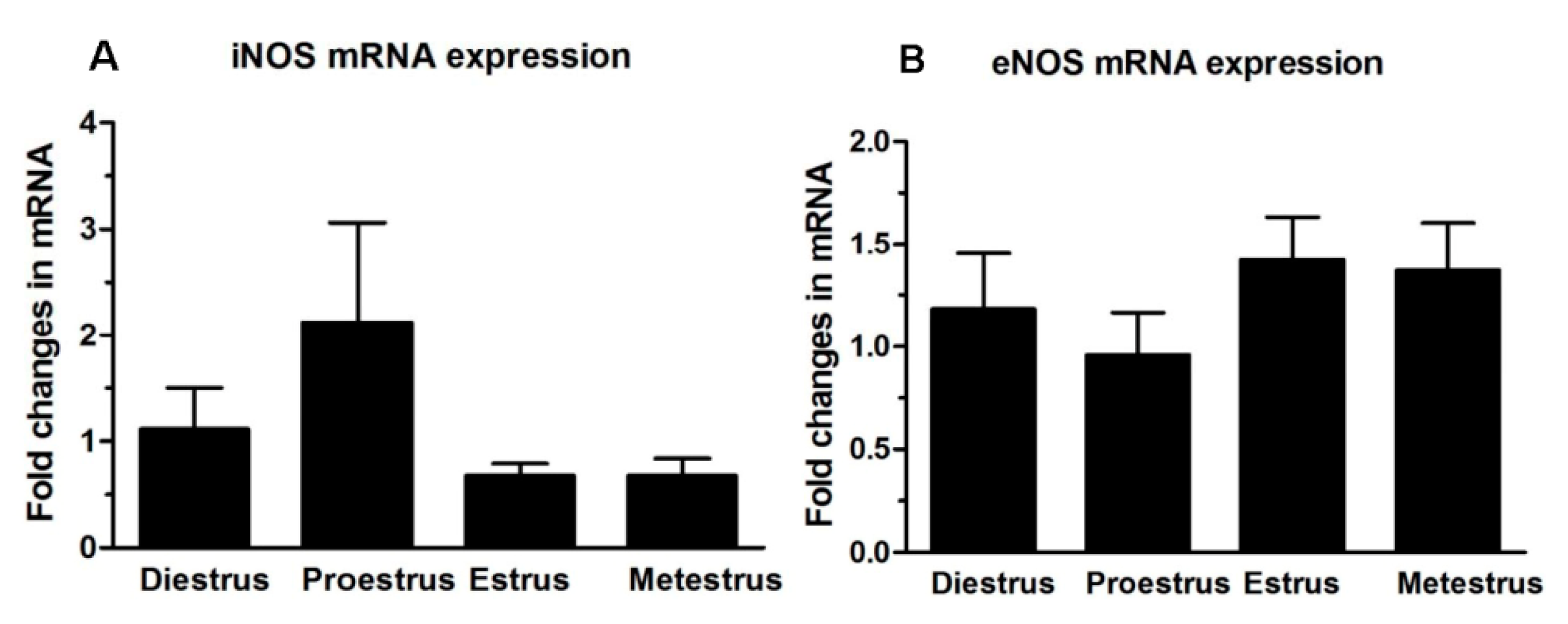
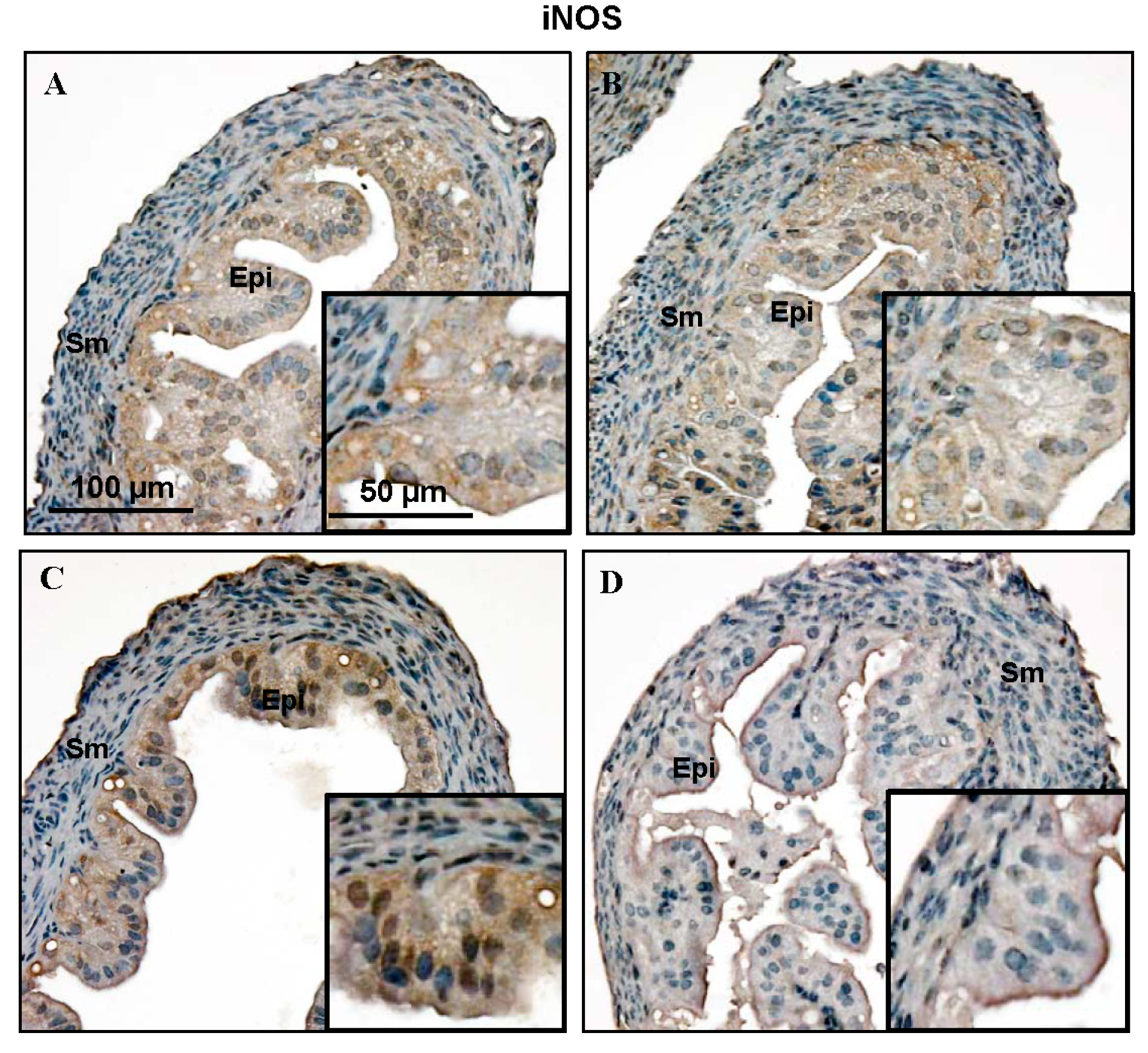
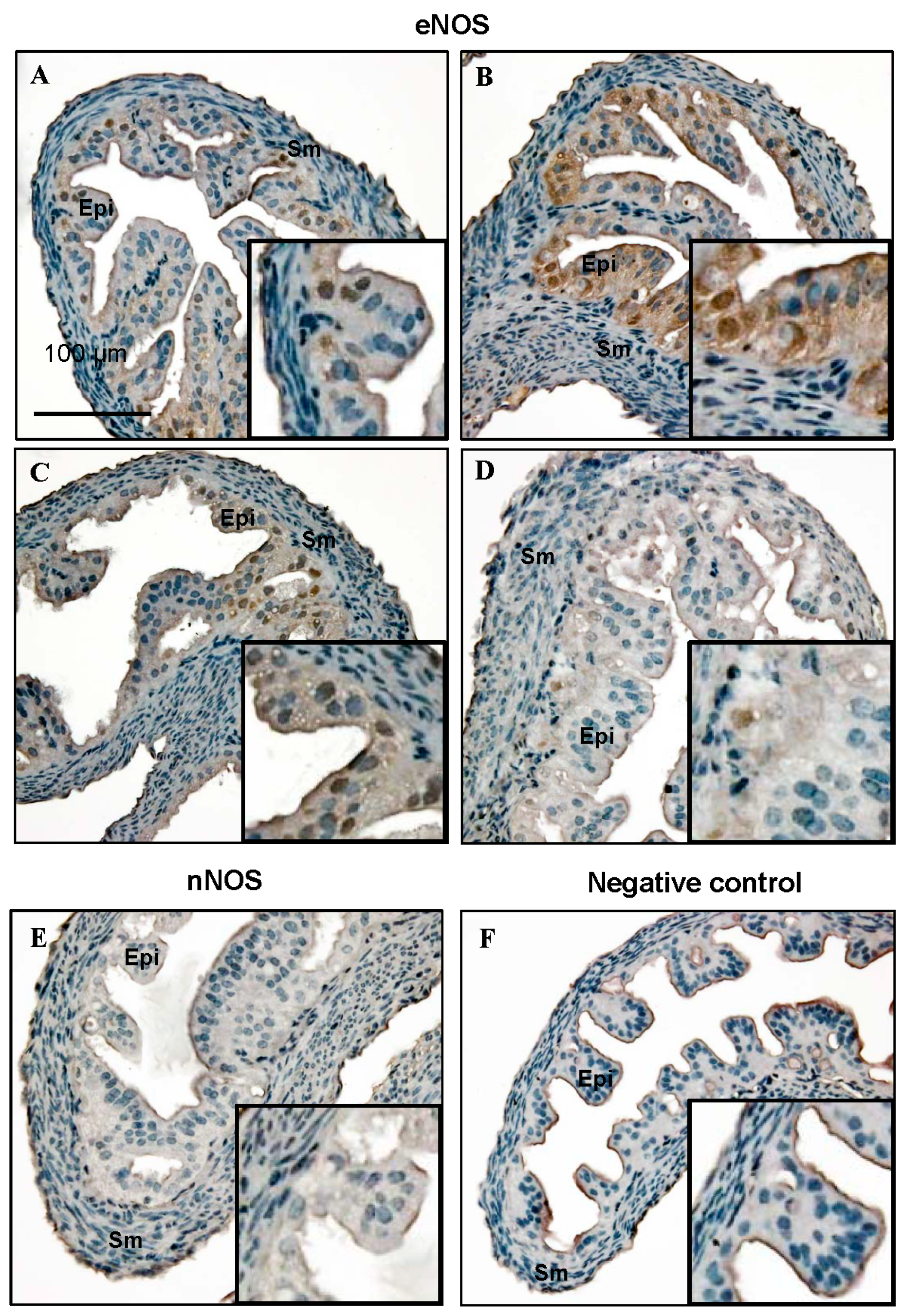
2.1. Nitric Oxide Synthase (NOS) Isoform Expression during the Estrous Cycle in Mouse Fallopian Tubes




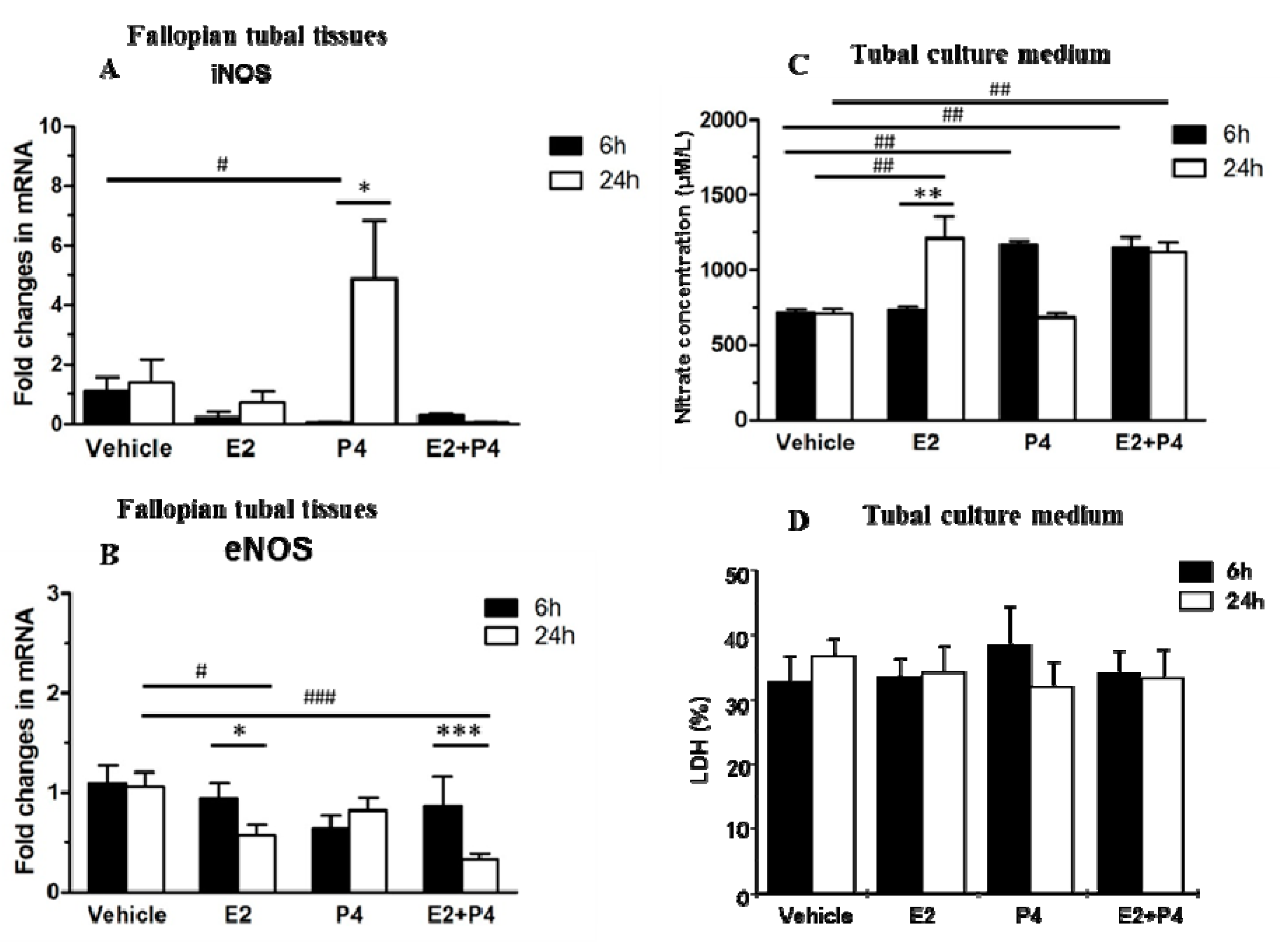
2.2. Hormones Directly Regulate NOS Isoform Expression in the Mouse Fallopian Tube
2.3. NO Levels and Lactate Dehydrogenase (LDH) Release Levels in Mouse Fallopian Tube Cultures

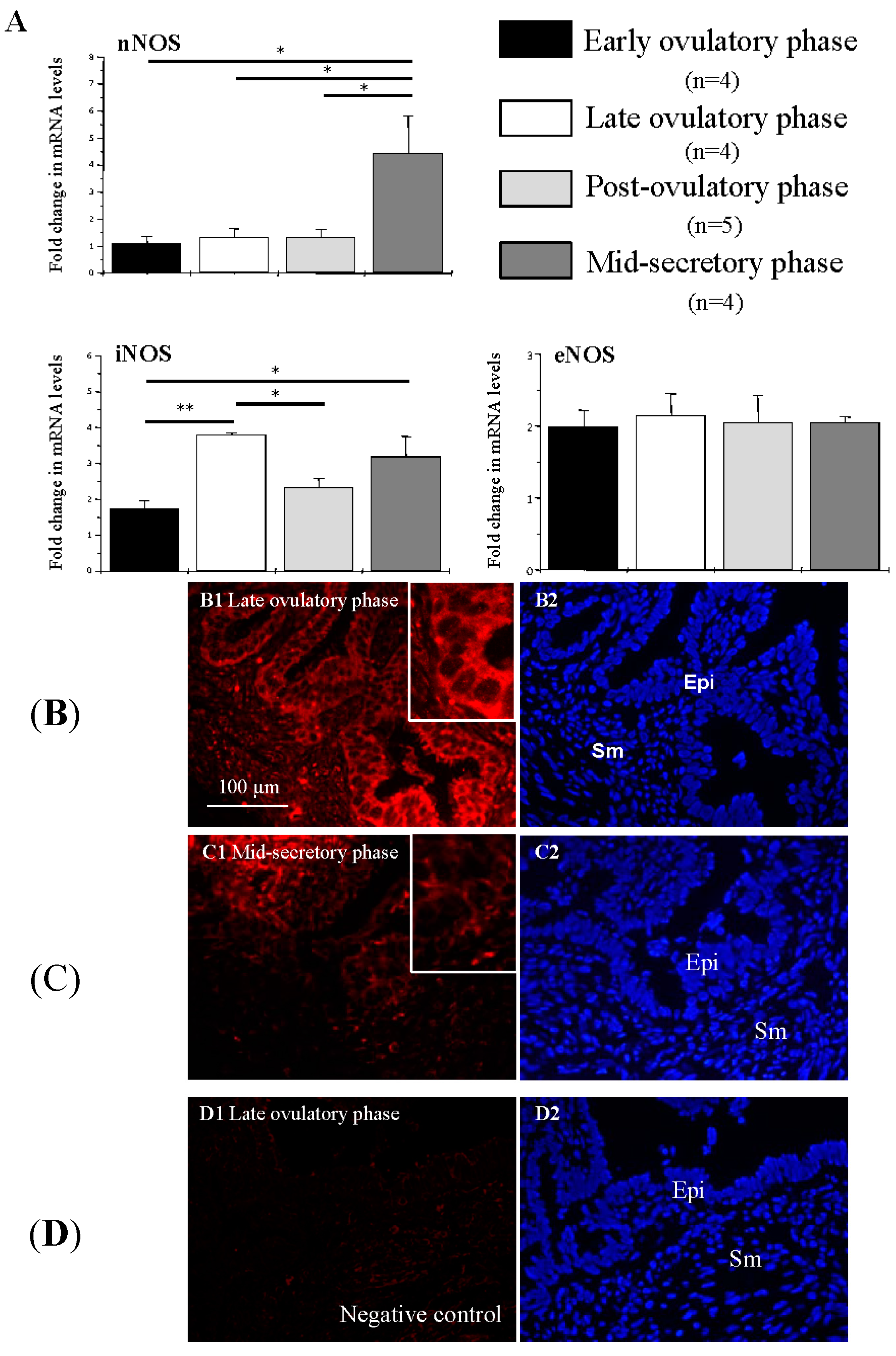
2.4. NOS Isoform Expression in Human Fallopian Tubes during the Menstrual Cycle

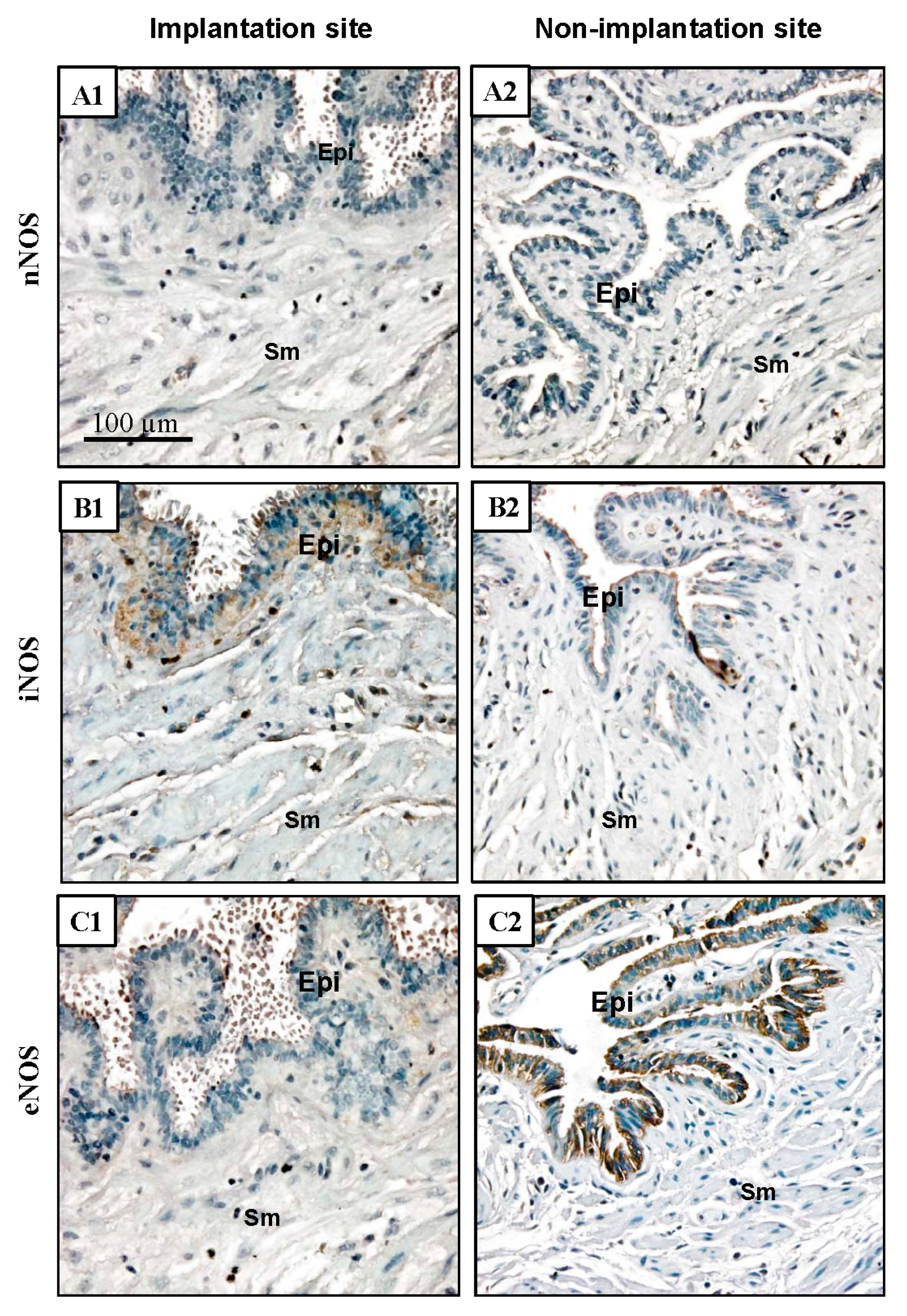
2.5. NOS Isoform Expression during Human Ectopic Pregnancy

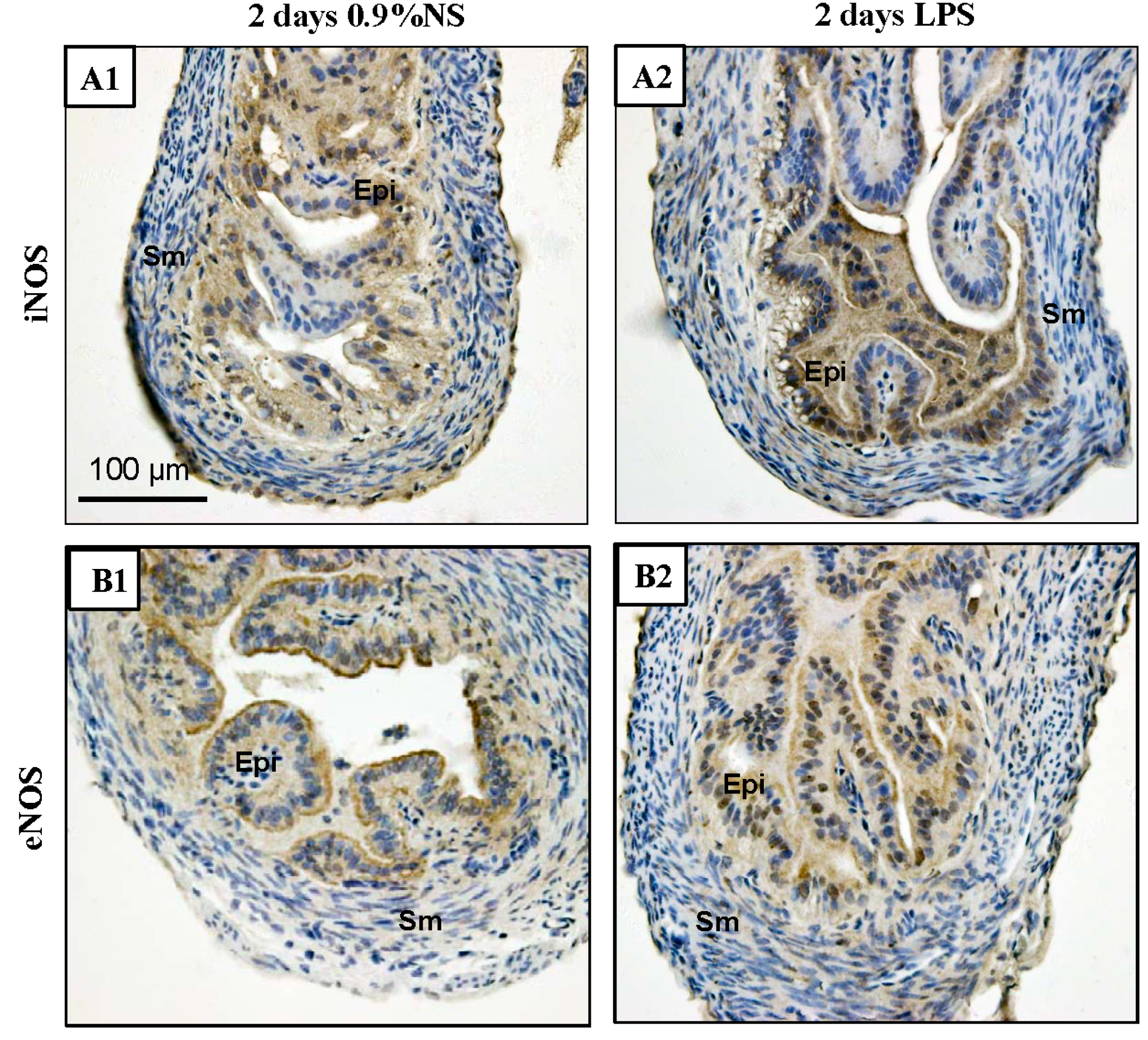
2.6. Increased iNOS Expression in Mice Treated with Lipopolysaccharide (LPS)

3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Human Tissues
4.3. Tissue Culture and Hormonal Treatment
4.4. Immunochemistry and Immunofluorescence
4.5. RNA Extraction and Quantitative Real-Time PCR Analysis
4.6. Measurement of NO Levels in Fallopian Tube Cultures
4.7. Lactate Dehydrogenase (LDH) Release Assay
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ekerhovd, E.; Brannstrom, M.; Alexandersson, M.; Norstrom, A. Evidence for nitric oxide mediation of contractile activity in isolated strips of the human Fallopian tube. Hum. Reprod. 1997, 12, 301–305. [Google Scholar] [PubMed]
- Rosselli, M.; Keller, P.J.; Dubey, R.K. Role of nitric oxide in the biology, physiology and pathophysiology of reproduction. Hum. Reprod. Update 1998, 4, 3–24. [Google Scholar] [CrossRef] [PubMed]
- Perez Martinez, S.; Viggiano, M.; Franchi, A.M.; Herrero, M.B.; Ortiz, M.E.; Gimeno, M.F.; Villalon, M. Effect of nitric oxide synthase inhibitors on ovum transport and oviductal smooth muscle activity in the rat oviduct. J. Reprod. Fertil. 2000, 118, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Szostek, A.Z.; Siemieniuch, M.J.; Deptula, K.; Woclawek-Potocka, I.; Majewska, M.; Okuda, K.; Skarzynski, D.J. Ovarian steroids modulate tumor necrosis factor-α and nitric oxide-regulated prostaglandin secretion by cultured bovine oviductal epithelial cells. Domest. Anim. Endocrinol. 2011, 41, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Kim, C.H.; Jung, K.Y.; Jeon, B.H.; Ju, E.J.; Choo, Y.K. Involvement of nitric oxide during in vitro fertilization and early embryonic development in mice. Arch. Pharm. Res. 2004, 27, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Al-Azemi, M.; Refaat, B.; Amer, S.; Ola, B.; Chapman, N.; Ledger, W. The expression of inducible nitric oxide synthase in the human fallopian tube during the menstrual cycle and in ectopic pregnancy. Fertil. Steril. 2010, 94, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Tempfer, C.; Moreno, R.M.; O’Brien, W.E.; Gregg, A.R. Genetic contributions of the endothelial nitric oxide synthase gene to ovulation and menopause in a mouse model. Fertil. Steril. 2000, 73, 1025–1031. [Google Scholar] [CrossRef] [PubMed]
- Bryan, N.S.; Bian, K.; Murad, F. Discovery of the nitric oxide signaling pathway and targets for drug development. Front. Biosci. 2009, 14, 1–18. [Google Scholar] [CrossRef]
- Maul, H.; Longo, M.; Saade, G.R.; Garfield, R.E. Nitric oxide and its role during pregnancy: From ovulation to delivery. Curr. Pharm. Des. 2003, 9, 359–380. [Google Scholar] [CrossRef] [PubMed]
- Refaat, B.; Al-Azemi, M.; Geary, I.; Eley, A.; Ledger, W. Role of activins and inducible nitric oxide in the pathogenesis of ectopic pregnancy in patients with or without Chlamydia trachomatis infection. Clin. Vaccine Immunol. 2009, 16, 1493–1503. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.J. Ectopic pregnancy. Clin. Obstet. Gynecol. 2007, 50, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Rana, P.; Kazmi, I.; Singh, R.; Afzal, M.; Al-Abbasi, F.A.; Aseeri, A.; Singh, R.; Khan, R.; Anwar, F. Ectopic pregnancy: A review. Arch. Gynecol. Obstet. 2013, 288, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.L.; Dey, S.K.; Critchley, H.O.; Horne, A.W. Current knowledge of the aetiology of human tubal ectopic pregnancy. Hum. Reprod. Update 2010, 16, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, H.; Gervaise, A. Ectopic pregnancies after infertility treatment: Modern diagnosis and therapeutic strategy. Hum. Reprod. Update 2004, 10, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Prantner, D.; Darville, T.; Sikes, J.D.; Andrews, C.W., Jr.; Brade, H.; Rank, R.G.; Nagarajan, U.M. Critical role for interleukin-1beta (IL-1β) during Chlamydia muridarum genital infection and bacterial replication-independent secretion of IL-1β in mouse macrophages. Infect. Immun. 2009, 77, 5334–5346. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.; Hu, J.; Billig, H. Toward understanding Chlamydia infection-induced infertility caused by dysfunctional oviducts. J. Infect. Dis. 2013, 208, 707–709. [Google Scholar] [CrossRef] [PubMed]
- Shao, R. Cigarette smoking effect on oviductal ciliation and ciliogenesis. Fertil. Steril. 2013, 99, e5. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, V.; Jaiswal, M.K.; Jaiswal, Y.K. LPS-induced implantation failure: One of the causes of female infertility. Med. J. Obstet. Gynecol. 2013, 1, 1014. [Google Scholar]
- Lapointe, J.; Roy, M.; St-Pierre, I.; Kimmins, S.; Gauvreau, D.; MacLaren, L.A.; Bilodeau, J.F. Hormonal and spatial regulation of nitric oxide synthases (NOS) (neuronal NOS, inducible NOS, and endothelial NOS) in the oviducts. Endocrinology 2006, 147, 5600–5610. [Google Scholar] [CrossRef] [PubMed]
- Bryant, C.E.; Tomlinson, A.; Mitchell, J.A.; Thiemermann, C.; Willoughby, D.A. Nitric oxide synthase in the rat fallopian tube is regulated during the oestrous cycle. J. Endocrinol. 1995, 146, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Gangula, P.R.; Dong, Y.L.; Yallampalli, C. Immunocytochemical localization of nitric oxide synthase-III in reproductive organs of female rats during the oestrous cycle. Histochem. J. 1996, 28, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Gawronska, B.; Bodek, G.; Ziecik, A.J. Distribution of NADPH-diaphorase and nitric oxide synthase (NOS) in different regions of porcine oviduct during the estrous cycle. J. Histochem. Cytochem. 2000, 48, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Ulbrich, S.E.; Rehfeld, S.; Bauersachs, S.; Wolf, E.; Rottmayer, R.; Hiendleder, S.; Vermehren, M.; Sinowatz, F.; Meyer, H.H.; Einspanier, R. Region-specific expression of nitric oxide synthases in the bovine oviduct during the oestrous cycle and in vitro. J. Endocrinol. 2006, 188, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, O.; Calka, J.; Bukowski, R.; Zalecki, M.; Wasowicz, K.; Jaroszewski, J.J.; Markiewicz, W.; Bulbul, A.; Ucar, M. Nitric oxide in the bovine oviduct: Influence on contractile activity and nitric oxide synthase isoforms localization. Theriogenology 2012, 77, 1312–1327. [Google Scholar] [CrossRef] [PubMed]
- Rosselli, M.; Imthurn, B.; Macas, E.; Keller, P.J.; Dubey, R.K. Endogenous nitric oxide modulates endothelin-1 induced contraction of bovine oviduct. Biochem. Biophys. Res. Commun. 1994, 201, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Ekerhovd, E.; Enskog, A.; Caidahl, K.; Klintland, N.; Nilsson, L.; Brannstrom, M.; Norstrom, A. Plasma concentrations of nitrate during the menstrual cycle, ovarian stimulation and ovarian hyperstimulation syndrome. Hum. Reprod. 2001, 16, 1334–1339. [Google Scholar] [CrossRef] [PubMed]
- Rosselli, M.; Dubey, R.K.; Rosselli, M.A.; Macas, E.; Fink, D.; Lauper, U.; Keller, P.J.; Imthurn, B. Identification of nitric oxide synthase in human and bovine oviduct. Mol. Hum. Reprod. 1996, 2, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Ekerhovd, E.; Brannstrom, M.; Weijdegard, B.; Norstrom, A. Localization of nitric oxide synthase and effects of nitric oxide donors on the human Fallopian tube. Mol. Hum. Reprod. 1999, 5, 1040–1047. [Google Scholar] [CrossRef] [PubMed]
- Gyurko, R.; Leupen, S.; Huang, P.L. Deletion of exon 6 of the neuronal nitric oxide synthase gene in mice results in hypogonadism and infertility. Endocrinology 2002, 143, 2767–2774. [Google Scholar] [CrossRef] [PubMed]
- Burnett, T.G.; Tash, J.S.; Hunt, J.S. Investigation of the role of nitric oxide synthase 2 in pregnancy using mutant mice. Reproduction 2002, 124, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Hefler, L.A.; Tempfer, C.B.; Moreno, R.M.; O’Brien, W.E.; Gregg, A.R. Endothelial-derived nitric oxide and angiotensinogen: Blood pressure and metabolism during mouse pregnancy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R174–R182. [Google Scholar] [PubMed]
- Pallares, P.; Garcia-Fernandez, R.A.; Criado, L.M.; Letelier, C.A.; Esteban, D.; Fernandez-Toro, J.M.; Flores, J.M.; Gonzalez-Bulnes, A. Disruption of the endothelial nitric oxide synthase gene affects ovulation, fertilization and early embryo survival in a knockout mouse model. Reproduction 2008, 136, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Tranguch, S.; Huet-Hudson, Y. Decreased viability of nitric oxide synthase double knockout mice. Mol. Reprod. Dev. 2003, 65, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Liu, V.W.; Huang, P.L. Cardiovascular roles of nitric oxide: A review of insights from nitric oxide synthase gene disrupted mice. Cardiovasc. Res. 2008, 77, 19–29. [Google Scholar]
- MacMicking, J.D.; Nathan, C.; Hom, G.; Chartrain, N.; Fletcher, D.S.; Trumbauer, M.; Stevens, K.; Xie, Q.W.; Sokol, K.; Hutchinson, N.; et al. Altered responses to bacterial infection and endotoxic shock in mice lacking inducible nitric oxide synthase. Cell 1995, 81, 641–650. [Google Scholar]
- Wei, X.Q.; Charles, I.G.; Smith, A.; Ure, J.; Feng, G.J.; Huang, F.P.; Xu, D.; Muller, W.; Moncada, S.; Liew, F.Y. Altered immune responses in mice lacking inducible nitric oxide synthase. Nature 1995, 375, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Hillis, S.D.; Owens, L.M.; Marchbanks, P.A.; Amsterdam, L.F.; Mac Kenzie, W.R. Recurrent chlamydial infections increase the risks of hospitalization for ectopic pregnancy and pelvic inflammatory disease. Am. J. Obstet. Gynecol. 1997, 176, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Wira, C.R.; Fahey, J.V.; Sentman, C.L.; Pioli, P.A.; Shen, L. Innate and adaptive immunity in female genital tract: Cellular responses and interactions. Immunol. Rev. 2005, 206, 306–335. [Google Scholar] [CrossRef] [PubMed]
- Igietseme, J.U.; Perry, L.L.; Ananaba, G.A.; Uriri, I.M.; Ojior, O.O.; Kumar, S.N.; Caldwell, H.D. Chlamydial infection in inducible nitric oxide synthase knockout mice. Infect. Immun. 1998, 66, 1282–1286. [Google Scholar] [PubMed]
- Dixon, R.E.; Hwang, S.J.; Hennig, G.W.; Ramsey, K.H.; Schripsema, J.H.; Sanders, K.M.; Ward, S.M. Chlamydia infection causes loss of pacemaker cells and inhibits oocyte transport in the mouse oviduct. Biol. Reprod. 2009, 80, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Stephens, R.S. The cellular paradigm of chlamydial pathogenesis. Trends Microbiol. 2003, 11, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Entrican, G.; Wattegedera, S.; Rocchi, M.; Fleming, D.C.; Kelly, R.W.; Wathne, G.; Magdalenic, V.; Howie, S.E. Induction of inflammatory host immune responses by organisms belonging to the genera Chlamydia/Chlamydophila. Vet. Immunol. Immunopathol. 2004, 100, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Marconi, C.; Santos-Greatti, M.M.; Parada, C.M.; Pontes, A.; Pontes, A.G.; Giraldo, P.C.; Donders, G.G.; da Silva, M.G. Cervicovaginal levels of proinflammatory cytokines are increased during chlamydial infection in bacterial vaginosis but not in lactobacilli-dominated flora. J. Low. Genit. Tract Dis. 2014, 18, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Bayati, L.F. Association between increased expression of endothelial isoform of nitric oxide synthase in the human fallopian tube and tubal ectopic pregnancy. Iran J. Reprod. Med. 2014, 12, 19–28. [Google Scholar] [PubMed]
- Shao, R.; Norstrom, A.; Weijdegard, B.; Egecioglu, E.; Fernandez-Rodriguez, J.; Feng, Y.; Stener-Victorin, E.; Brannstrom, M.; Billig, H. Distinct expression pattern of Dicer1 correlates with ovarian-derived steroid hormone receptor expression in human Fallopian tubes during ovulation and the midsecretory phase. J. Clin. Endocrinol. Metab. 2011, 96, E869–E877. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.; Wang, X.; Weijdegard, B.; Norstrom, A.; Fernandez-Rodriguez, J.; Brannstrom, M.; Billig, H. Coordinate regulation of heterogeneous nuclear ribonucleoprotein dynamics by steroid hormones in the human fallopian tube and endometrium in vivo and in vitro. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1269–E1282. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.; Weijdegard, B.; Fernandez-Rodriguez, J.; Egecioglu, E.; Zhu, C.; Andersson, N.; Thurin-Kjellberg, A.; Bergh, C.; Billig, H. Ciliated epithelial-specific and regional-specific expression and regulation of the estrogen receptor-β2 in the fallopian tubes of immature rats: A possible mechanism for estrogen-mediated transport process in vivo. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E147–E158. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.; Nutu, M.; Weijdegard, B.; Egecioglu, E.; Fernandez-Rodriguez, J.; Karlsson-Lindahl, L.; Gemzell-Danielsson, K.; Bergh, C.; Billig, H. Clomiphene citrate causes aberrant tubal apoptosis and estrogen receptor activation in rat fallopian tube: implications for tubal ectopic pregnancy. Biol. Reprod. 2009, 80, 1262–1271. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Ma, S.; Zou, S.; Li, X.; Cui, P.; Weijdegård, B.; Wu, G.; Shao, R.; Billig, H.; Feng, Y. The Regulation of Nitric Oxide Synthase Isoform Expression in Mouse and Human Fallopian Tubes: Potential Insights for Ectopic Pregnancy. Int. J. Mol. Sci. 2015, 16, 49-67. https://doi.org/10.3390/ijms16010049
Hu J, Ma S, Zou S, Li X, Cui P, Weijdegård B, Wu G, Shao R, Billig H, Feng Y. The Regulation of Nitric Oxide Synthase Isoform Expression in Mouse and Human Fallopian Tubes: Potential Insights for Ectopic Pregnancy. International Journal of Molecular Sciences. 2015; 16(1):49-67. https://doi.org/10.3390/ijms16010049
Chicago/Turabian StyleHu, Junting, Shulan Ma, Sien Zou, Xin Li, Peng Cui, Birgitta Weijdegård, Gencheng Wu, Ruijin Shao, Håkan Billig, and Yi Feng. 2015. "The Regulation of Nitric Oxide Synthase Isoform Expression in Mouse and Human Fallopian Tubes: Potential Insights for Ectopic Pregnancy" International Journal of Molecular Sciences 16, no. 1: 49-67. https://doi.org/10.3390/ijms16010049
APA StyleHu, J., Ma, S., Zou, S., Li, X., Cui, P., Weijdegård, B., Wu, G., Shao, R., Billig, H., & Feng, Y. (2015). The Regulation of Nitric Oxide Synthase Isoform Expression in Mouse and Human Fallopian Tubes: Potential Insights for Ectopic Pregnancy. International Journal of Molecular Sciences, 16(1), 49-67. https://doi.org/10.3390/ijms16010049