
Effects of Ethanol on the Expression Level of Various BDNF mRNA Isoforms and Their Encoded Protein in the Hippocampus of Adult and Embryonic Rats



Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects of Eethanol on the Expression Pattern of Exons I, III, IV and IX of the BDNF Gene in the Hippocampus of Adult and Embryonic Rats


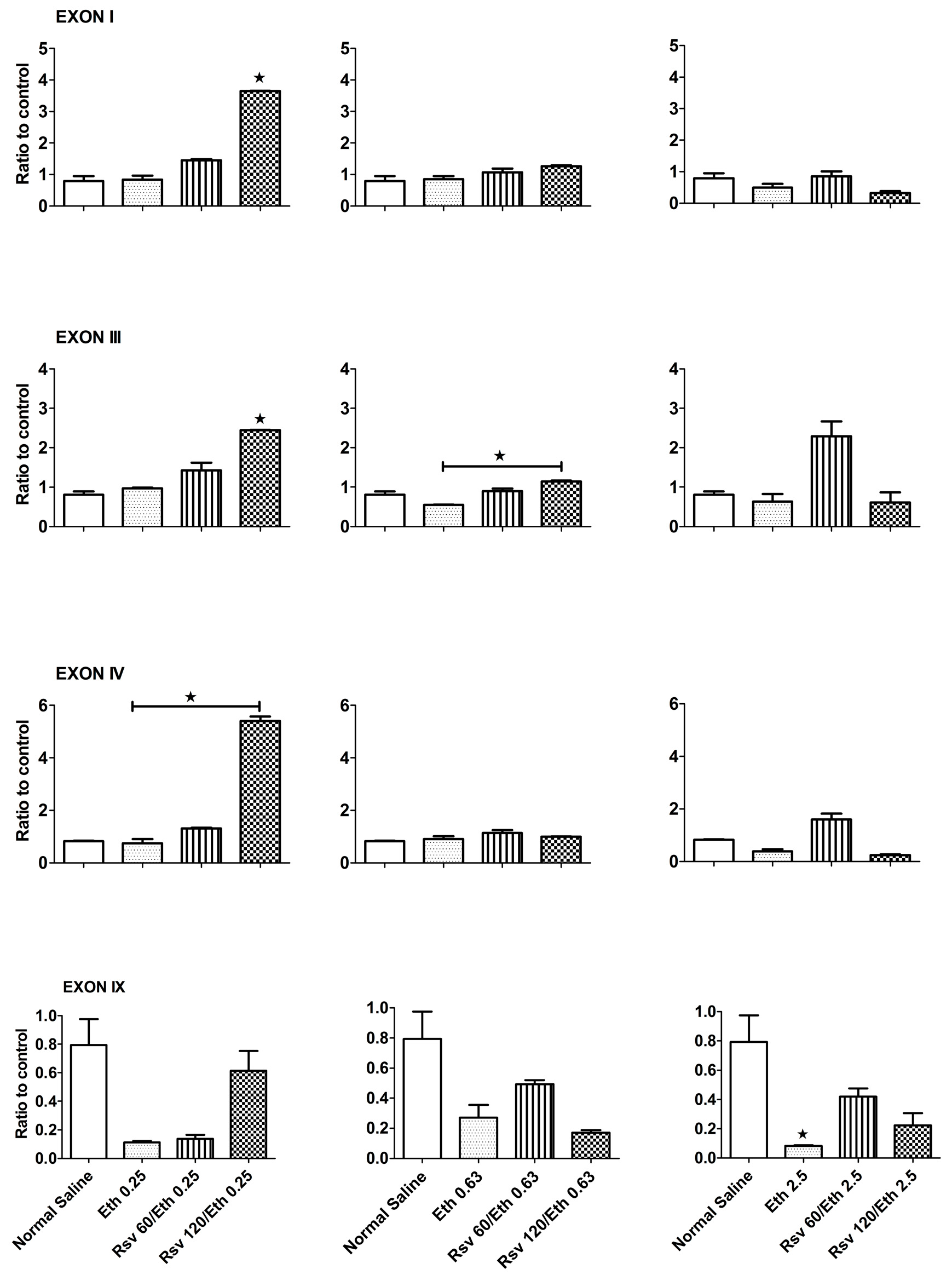{kind=link}
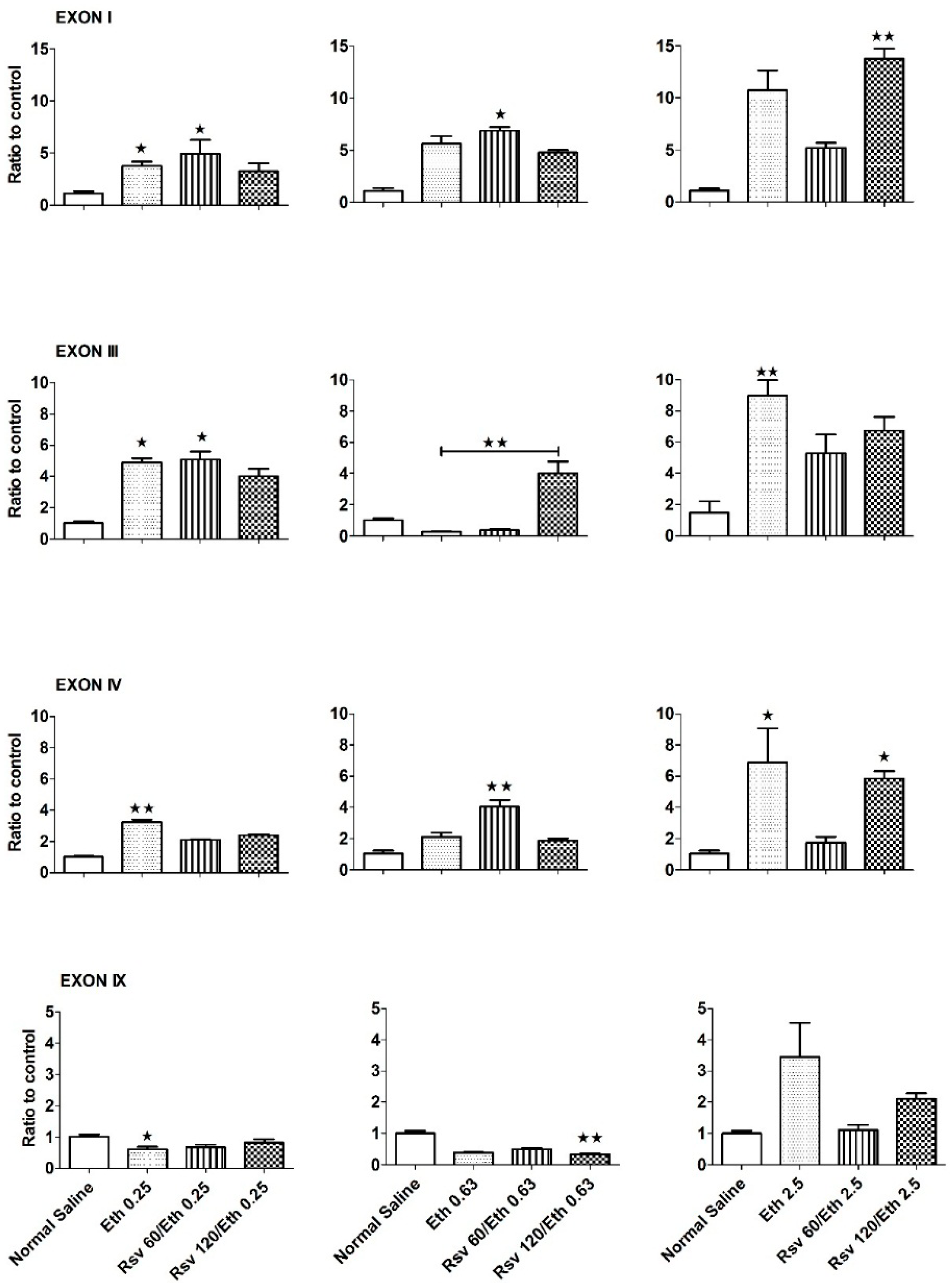{kind=link}
{kind=link}
| Treatment | Normal Saline | Ethanol 250 mg/kg·BW | Ethanol 630 mg/kg·BW | Ethanol 2500 mg/kg·BW | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CT | Eth | Rsv mg/kg·BW | Eth | Rsv mg/kg·BW | Eth | Rsv mg/kg·BW | ||||
| NS | Eth | 60/Eth | 120/Eth | Eth | 60/Eth | 120/Eth | Eth | 60/Eth | 120/Eth | |
| Group | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
2.2. Effects of Combined Eethanol and Rresveratrol on the Expression Pattern of Four Transcripts of the BDNF Gene in the Hippocampi of Adult Rats

2.3. Effects of Combined Eethanol and Rresveratrol on the Expression Pattern of Four BDNF Gene Transcripts in the Embryonic Rat Hippocampus
2.4. Effects of Eethanol and Rresveratrol on the Levels of Pro-BDNF Protein in the Hippocampus of Pregnant Rats and Their Embryos

| Treatment | Adult | Embryo | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| mRNA | Protein | mRNA | Protein | |||||||
| Exon I | Exon III | Exon IV | Exon IX | Pro-BDNF | Exon I | Exon III | Exon IV | Exon IX | Pro-BDNF | |
| Eth 0.25 | NS | NS | NS | NS | NS | ↑ | ↑ | ↑ | ↓ | NS |
| Rsv 60/Eth 0.25 | NS | NS | NS | NS | NS | ↑ | ↑ | NS | NS | NS |
| Rsv 120/Eth 0.25 | ↑ | ↑ | NS | NS | ↑ | NS | NS | NS | NS | NS |
| Eth 0.63 | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS |
| Rsv 60/Eth 0.63 | NS | NS | NS | NS | NS | ↑ | NS | ↑ | NS | NS |
| Rsv 120/Eth 0.63 | NS | NS | NS | NS | NS | NS | NS | NS | ↓ | NS |
| Eth 2.5 | NS | NS | NS | ↓ | ↑ | NS | ↑ | ↑ | NS | NS |
| Rsv 60/Eth2.5 | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS |
| Rsv 120/Eth 2.5 | NS | NS | NS | NS | NS | ↑ | NS | ↑ | NS | NS |
3. Experimental Section
3.1. Subjects and Experimental Design
3.2. Real-Time Quantitative PCR
| Gene Name | Primer Forward (5′ to 3′) | Primer Reverse (5′ to 3′) | Reference |
|---|---|---|---|
| BDNF EXON I | TGTTGGGGAGACGAGATTTT | CGTGGACGTTTGCTTCTTTC | [96] |
| BDNF EXON III | CTGAGACTGCGCTCCACTC | GTGGACGTTTGCTTCTTTCA | [96] |
| BDNF EXON IV | GAGCAGCTGCCTTGATGTTT | GTGGACGTTTGCTTCTTTCA | [96] |
| BDNF EXON IX | GTGACAGTATTAGCGAGTGGG | GGGTAGTTCGGCATTGC | [94] |
| β-actin | CCACACCCGCCACCAGTTCG | CTAGGGCGGCCCACGATGGA | [94] |
| HPRT | CCCAGCGTCGTGATTAGTGA | TGGCCTCCCATCTCCTTCAT | * |
| GAPDH | CGTGATCGAGGGCTGTTGG | CTGCTTCAGTTGGCCTTTCG | [97] |
3.4. Western Blotting
3.5. Data Analysis
Acknowledgments
Authors Contributions
Conflicts of Interest
References
- Alfonso-Loeches, S.; Guerri, C. Molecular and behavioral aspects of the actions of alcohol on the adult and developing brain. Crit. Rev. Clin. Lab. Sci. 2011, 48, 19–47. [Google Scholar] [CrossRef] [PubMed]
- Brust, J.C. Ethanol and cognition: Indirect effects, neurotoxicity and neuroprotection: A review. Int. J. Environ. Res. Public Health 2010, 7, 1540–1557. [Google Scholar] [CrossRef] [PubMed]
- Gerlai, R. Embryonic alcohol exposure: Towards the development of a zebrafish model of fetal alcohol spectrum disorders. Dev. Psychobiol. 2015, 57, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Harper, C.; Matsumoto, I. Ethanol and brain damage. Curr. Opin. Pharmacol. 2005, 5, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Novier, A.; van Skike, C.E.; Diaz-Granados, J.L.; Mittleman, G.; Matthews, D.B. Acute alcohol produces ataxia and cognitive impairments in aged animals: A comparison between young adult and aged rats. Alcohol. Clin. Exp. Res. 2013, 37, 1317–1324. [Google Scholar] [CrossRef] [PubMed]
- Kodituwakku, P.W. Neurocognitive profile in children with fetal alcohol spectrum disorders. Dev. Disabil. Res. Rev. 2009, 15, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Ismail, S.; Buckley, S.; Budacki, R.; Jabbar, A.; Gallicano, G.I. Screening, diagnosing and prevention of fetal alcohol syndrome: Is this syndrome treatable? Dev. Neurosci. 2010, 32, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Ikonomidou, C.; Bittigau, P.; Ishimaru, M.J.; Wozniak, D.F.; Koch, C.; Genz, K.; Price, M.T.; Stefovska, V.; Horster, F.; Tenkova, T.; et al. Ethanol-induced apoptotic neurodegeneration and fetal alcohol syndrome. Science 2000, 287, 1056–1060. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, K.K.; Sheema, S.; Paz, R.D.; Samudio-Ruiz, S.L.; Laughlin, M.H.; Spence, N.E.; Roehlk, M.J.; Alcon, S.N.; Allan, A.M. Fetal alcohol spectrum disorder-associated depression: Evidence for reductions in the levels of brain-derived neurotrophic factor in a mouse model. Pharmacol. Biochem. Behav. 2008, 90, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Berman, R.F.; Hannigan, J.H. Effects of prenatal alcohol exposure on the hippocampus: Spatial behavior, electrophysiology, and neuroanatomy. Hippocampus 2000, 10, 94–110. [Google Scholar] [CrossRef]
- Boschen, K.E.; Criss, K.J.; Palamarchouk, V.; Roth, T.L.; Klintsova, A.Y. Effects of developmental alcohol exposure vs. intubation stress on BDNF and TrkB expression in the hippocampus and frontal cortex of neonatal rats. Int. J. Dev. Neurosci. 2015, 43, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Raivio, N.; Tiraboschi, E.; Saarikoski, S.T.; Castren, E.; Kiianmaa, K. Brain-derived neurotrophic factor expression after acute administration of ethanol. Eur. J. Pharmacol. 2012, 687, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Dias, B.G.; Banerjee, S.B.; Duman, R.S.; Vaidya, V.A. Differential regulation of brain derived neurotrophic factor transcripts by antidepressant treatments in the adult rat brain. Neuropharmacology 2003, 45, 553–563. [Google Scholar] [CrossRef]
- Mei, Y.A.; Vaudry, D.; Basille, M.; Castel, H.; Fournier, A.; Vaudry, H.; Gonzalez, B.J. PACAP inhibits delayed rectifier potassium current via a cAMP/PKA transduction pathway: Evidence for the involvement of IK in the anti-apoptotic action of PACAP. Eur. J. Neurosci. 2004, 19, 1446–1458. [Google Scholar] [CrossRef] [PubMed]
- Heaton, M.B.; Paiva, M.; Mayer, J.; Miller, R. Ethanol-mediated generation of reactive oxygen species in developing rat cerebellum. Neurosci. Lett. 2002, 334, 83–86. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, C.K.; DiPette, D.D.; Singh, U.S. Ethanol impairs activation of retinoic acid receptors in cerebellar granule cells in a rodent model of fetal alcohol spectrum disorders. Alcohol. Clin. Exp. Res. 2010, 34, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.R.; Singleton, C.K.; Hiller-Sturmhofel, S. The role of thiamine deficiency in alcoholic brain disease. Alcohol Res. Health 2003, 27, 134–142. [Google Scholar] [PubMed]
- Ge, Y.; Belcher, S.M.; Light, K.E. Alterations of cerebellar mRNA specific for BDNF, p75NTR, and TrkB receptor isoforms occur within hours of ethanol administration to 4-day-old rat pups. Dev. Brain Res. 2004, 151, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Luo, J. Mechanisms of ethanol-induced death of cerebellar granule cells. Cerebellum 2012, 11, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Poo, M.M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.I. Ethanol-BDNF interactions: Still more questions than answers. Pharmacol. Ther. 2008, 118, 36–57. [Google Scholar] [CrossRef] [PubMed]
- Aid, T.; Kazantseva, A.; Piirsoo, M.; Palm, K.; Timmusk, T. Mouse and rat BDNF gene structure and expression revisited. J. Neurosci. Res. 2007, 85, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Pang, P.T.; Teng, H.K.; Zaitsev, E.; Woo, N.T.; Sakata, K.; Zhen, S.; Teng, K.K.; Yung, W.H.; Hempstead, B.L.; Lu, B. Cleavage of proBDNF by tPA/plasmin is essential for long-term hippocampal plasticity. Science 2004, 306, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and synaptic plasticity, cognitive function, and dysfunction. Handb. Exp. Pharmacol. 2014, 220, 223–250. [Google Scholar] [PubMed]
- Woo, N.H.; Teng, H.K.; Siao, C.J.; Chiaruttini, C.; Pang, P.T.; Milner, T.A.; Hempstead, B.L.; Lu, B. Activation of p75NTR by proBDNF facilitates hippocampal long-term depression. Nat. Neurosci. 2005, 8, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Je, H.S.; Yang, F.; Ji, Y.; Potluri, S.; Fu, X.Q.; Luo, Z.G.; Nagappan, G.; Chan, J.P.; Hempstead, B.; Son, Y.J.; et al. ProBDNF and mature BDNF as punishment and reward signals for synapse elimination at mouse neuromuscular junctions. J. Neurosci. 2013, 33, 9957–9962. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, G.S.; Dickson, K.S.; Gray, N.K. Regulation of mRNA translation by 5′- and 3′-UTR-binding factors. Trends Biochem. Sci. 2003, 28, 182–188. [Google Scholar] [CrossRef]
- Vaghi, V.; Polacchini, A.; Baj, G.; Pinheiro, V.L.; Vicario, A.; Tongiorgi, E. Pharmacological profile of brain-derived neurotrophic factor (BDNF) splice variant translation using a novel drug screening assay: A “quantitative code”. J. Biol. Chem. 2014, 289, 27702–27713. [Google Scholar] [CrossRef] [PubMed]
- Perovic, M.; Tesic, V.; Mladenovic Djordjevic, A.; Smiljanic, K.; Loncarevic-Vasiljkovic, N.; Ruzdijic, S.; Kanazir, S. BDNF transcripts, proBDNF and proNGF, in the cortex and hippocampus throughout the life span of the rat. Age 2012, 35, 2057–2070. [Google Scholar] [CrossRef] [PubMed]
- Baj, G.; Leone, E.; Chao, M.V.; Tongiorgi, E. Spatial segregation of BDNF transcripts enables BDNF to differentially shape distinct dendritic compartments. Proc. Natl. Acad. Sci. USA 2011, 108, 16813–16818. [Google Scholar] [CrossRef] [PubMed]
- Granzotto, A.; Zatta, P. Resveratrol and Alzheimer’s disease: Message in a bottle on red wine and cognition. Front. Aging Neurosci. 2014, 6, 95. [Google Scholar] [CrossRef] [PubMed]
- Pallas, M.; Ortuno-Sahagun, D.; Benito-Andres, P.; Ponce-Regalado, M.D.; Rojas-Mayorquin, A.E. Resveratrol in epilepsy: Preventive or treatment opportunities? Front. Biosci. 2014, 19, 1057–1064. [Google Scholar] [CrossRef]
- Robb, E.L.; Stuart, J.A. Trans-resveratrol as a neuroprotectant. Molecules 2010, 15, 1196–1212. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.A.; de Carvaho, G.S.; Oliveira, V.; Raposo, N.R.; da Silva, A.D. Resveratrol and analogues: A review of antioxidant activity and applications to human health. Recent Pat. Food Nutr. Agric. 2013, 5, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Ramis, M.R.; Esteban, S.; Miralles, A.; Tan, D.X.; Reiter, R.J. Caloric restriction, resveratrol and melatonin: Role of SIRT1 and implications for aging and related-diseases. Mech. Ageing Dev. 2015, 146–148C, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Negi, G.; Sharma, S.S. Neuroprotection by resveratrol in diabetic neuropathy: Concepts & mechanisms. Curr. Med. Chem. 2013, 20, 4640–4645. [Google Scholar] [PubMed]
- Pallas, M.; Casadesus, G.; Smith, M.A.; Coto-Montes, A.; Pelegri, C.; Vilaplana, J.; Camins, A. Resveratrol and neurodegenerative diseases: Activation of SIRT1 as the potential pathway towards neuroprotection. Curr. Neurovasc. Res. 2009, 6, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Rahvar, M.; Nikseresht, M.; Shafiee, S.M.; Naghibalhossaini, F.; Rasti, M.; Panjehshahin, M.R.; Owji, A.A. Effect of oral resveratrol on the BDNF gene expression in the hippocampus of the rat brain. Neurochem. Res. 2011, 36, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, Q.; Gu, J.; Wang, X.; Xie, K.; Xian, X.; Wang, J.; Jiang, H.; Wang, Z. Resveratrol prevents impaired cognition induced by chronic unpredictable mild stress in rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 49, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Lucio, M.; Lima, J.L.; Reis, S. Resveratrol in medicinal chemistry: A critical review of its pharmacokinetics, drug-delivery, and membrane interactions. Curr. Med. Chem. 2012, 19, 1663–1681. [Google Scholar] [CrossRef] [PubMed]
- Bourque, S.L.; Dolinsky, V.W.; Dyck, J.R.; Davidge, S.T. Maternal resveratrol treatment during pregnancy improves adverse fetal outcomes in a rat model of severe hypoxia. Placenta 2012, 33, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Russo-Neustadt, A.A.; Alejandre, H.; Garcia, C.; Ivy, A.S.; Chen, M.J. Hippocampal brain-derived neurotrophic factor expression following treatment with reboxetine, citalopram, and physical exercise. Neuropsychopharmacology 2004, 29, 2189–2199. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, S.; Liang, M.H.; Marinova, Z.; Yahyavi, A.; Chuang, D.M. The mood stabilizers lithium and valproate selectively activate the promoter IV of brain-derived neurotrophic factor in neurons. Mol. Psychiatry 2009, 14, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, F.; Molteni, R.; Maj, P.F.; Cattaneo, A.; Gennarelli, M.; Racagni, G.; Riva, M.A. Chronic duloxetine treatment induces specific changes in the expression of BDNF transcripts and in the subcellular localization of the neurotrophin protein. Neuropsychopharmacology 2007, 32, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Status Report on Alcohol and Health 2014; World Health Organization: Geneva, Switzerland, 2014; pp. 7–8. [Google Scholar]
- Rasmussen, C. Executive functioning and working memory in fetal alcohol spectrum disorder. Alcohol. Clin. Exp. Res. 2005, 29, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Roebuck, T.M.; Mattson, S.N.; Riley, E.P. A review of the neuroanatomical findings in children with fetal alcohol syndrome or prenatal exposure to alcohol. Alcohol. Clin. Exp. Res. 1998, 22, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Lindsley, T.A.; Shah, S.N.; Ruggiero, E.A. Ethanol alters BDNF-induced Rho GTPase activation in axonal growth cones. Alcohol. Clin. Exp. Res. 2011, 35, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.I.; Ju, A.; Lee, H.M.; Lee, S.S.; Song, C.H.; Won, W.Y.; Jeong, J.S.; Hong, O.K.; Kim, J.H.; Kim, D.J. Chronic ethanol ingestion, type 2 diabetes mellitus, and brain-derived neurotrophic factor (BDNF) in rats. Neurosci. Lett. 2011, 487, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Logrip, M.L.; Janak, P.H.; Ron, D. Escalating ethanol intake is associated with altered corticostriatal BDNF expression. J. Neurochem. 2009, 109, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Hauser, S.R.; Getachew, B.; Taylor, R.E.; Tizabi, Y. Alcohol induced depressive-like behavior is associated with a reduction in hippocampal BDNF. Pharmacol. Biochem. Behav. 2011, 100, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Arancibia, L.; Rage, F.; Givalois, L.; Dingeon, P.; Arancibia, S.; Beauge, F. Effects of alcohol on brain-derived neurotrophic factor mRNA expression in discrete regions of the rat hippocampus and hypothalamus. J. Neurosci. Res. 2001, 63, 200–208. [Google Scholar] [CrossRef]
- Miller, R.; King, M.A.; Heaton, M.B.; Walker, D.W. The effects of chronic ethanol consumption on neurotrophins and their receptors in the rat hippocampus and basal forebrain. Brain Res. 2002, 950, 137–147. [Google Scholar] [CrossRef]
- Okamoto, H.; Miki, T.; Lee, K.Y.; Yokoyama, T.; Kuma, H.; Gu, H.; Li, H.P.; Matsumoto, Y.; Yamaoka, I.; Fusumada, K.; et al. Effects of chronic ethanol administration on the expression levels of neurotrophic factors in the rat hippocampus. Okajimas Folia Anat. Jpn. 2006, 83, 1–6. [Google Scholar] [CrossRef] [PubMed]
- McGough, N.N.; He, D.Y.; Logrip, M.L.; Jeanblanc, J.; Phamluong, K.; Luong, K.; Kharazia, V.; Janak, P.H.; Ron, D. RACK1 and brain-derived neurotrophic factor: A homeostatic pathway that regulates alcohol addiction. J. Neurosci. 2004, 24, 10542–10552. [Google Scholar] [CrossRef] [PubMed]
- Raivio, N.; Miettinen, P.; Kiianmaa, K. Innate BDNF expression is associated with ethanol intake in alcohol-preferring AA and alcohol-avoiding ANA rats. Brain Res. 2014, 1579, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.K.; Heaton, M.B.; Walker, D.W. Up-regulation of high-affinity neurotrophin receptor, trk B-like protein on Western blots of rat cortex after chronic ethanol treatment. Mol. Brain Res. 1996, 40, 161–164. [Google Scholar] [CrossRef]
- Stragier, E.; Massart, R.; Salery, M.; Hamon, M.; Geny, D.; Martin, V.; Boulle, F.; Lanfumey, L. Ethanol-induced epigenetic regulations at the BDNF gene in C57BL/6J mice. Mol Psychiatry 2015, 20, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Kulkarny, V.V.; Wiest, N.E.; Marquez, C.P.; Nixon, S.C.; Valenzuela, C.F.; Perrone-Bizzozero, N.I. Opposite effects of acute ethanol exposure on GAP-43 and BDNF expression in the hippocampus versus the cerebellum of juvenile rats. Alcohol 2011, 45, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Baldwin, L.A. Ethanol and hormesis. Crit. Rev. Toxicol. 2003, 33, 407–424. [Google Scholar] [CrossRef] [PubMed]
- Hayes, D.P. Nutritional hormesis. Eur. J. Clin. Nutr. 2007, 61, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.J.; Yan, S.E.; Yan, Q.S. Effects of prenatal alcohol exposure on brain-derived neurotrophic factor and its receptor tyrosine kinase B in offspring. Brain Res. 2005, 1042, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Miki, T.; Kuma, H.; Yokoyama, T.; Sumitani, K.; Matsumoto, Y.; Kusaka, T.; Warita, K.; Wang, Z.Y.; Hosomi, N.; Imagawa, T.; et al. Early postnatal ethanol exposure induces fluctuation in the expression of BDNF mRNA in the developing rat hippocampus. Acta Neurobiol. Exp. 2008, 68, 484–493. [Google Scholar]
- Hossain, A.; Hajman, K.; Charitidi, K.; Erhardt, S.; Zimmermann, U.; Knipper, M.; Canlon, B. Prenatal dexamethasone impairs behavior and the activation of the BDNF exon IV promoter in the paraventricular nucleus in adult offspring. Endocrinology 2008, 149, 6356–6365. [Google Scholar] [CrossRef] [PubMed]
- Hara, D.; Miyashita, T.; Fukuchi, M.; Suzuki, H.; Azuma, Y.; Tabuchi, A.; Tsuda, M. Persistent BDNF exon I–IX mRNA expression following the withdrawal of neuronal activity in neurons. Biochem. Biophys. Res. Commun. 2009, 390, 648–653. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Rodriguez, M.; Lotfipour, S.; Leonard, G.; Perron, M.; Richer, L.; Veillette, S.; Pausova, Z.; Paus, T. Maternal smoking during pregnancy is associated with epigenetic modifications of the brain-derived neurotrophic factor-6 exon in adolescent offspring. Am. J. Med. Genet. Part B 2010, 153B, 1350–1354. [Google Scholar] [CrossRef] [PubMed]
- Peregud, D.I.; Panchenko, L.F.; Gulyaeva, N.V. Elevation of BDNF exon I-specific transcripts in the frontal cortex and midbrain of rat during spontaneous morphine withdrawal is accompanied by enhanced pCreb1 occupancy at the corresponding promoter. Neurochem. Res. 2015, 40, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.D.; Sangrey, G.R.; Darnell, S.B.; Schassburger, R.L.; Cha, J.H.; Pierce, R.C.; Sadri-Vakili, G. Increased brain-derived neurotrophic factor (BDNF) expression in the ventral tegmental area during cocaine abstinence is associated with increased histone acetylation at BDNF exon I-containing promoters. J. Neurochem. 2012, 120, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.B.; Abel, J.M.; Lynch, W.J. Dose-dependent effects of wheel running on cocaine-seeking and prefrontal cortex BDNF exon IV expression in rats. Psychopharmacology 2014, 231, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Sadri-Vakili, G.; Kumaresan, V.; Schmidt, H.D.; Famous, K.R.; Chawla, P.; Vassoler, F.M.; Overland, R.P.; Xia, E.; Bass, C.E.; Terwilliger, E.F.; et al. Cocaine-induced chromatin remodeling increases brain-derived neurotrophic factor transcription in the rat medial prefrontal cortex, which alters the reinforcing efficacy of cocaine. J. Neurosci. 2010, 30, 11735–11744. [Google Scholar] [CrossRef] [PubMed]
- Morioka, N.; Yoshida, Y.; Nakamura, Y.; Hidaka, N.; Hisaoka-Nakashima, K.; Nakata, Y. The regulation of exon-specific brain-derived neurotrophic factor mRNA expression by protein kinase c in rat cultured dorsal root ganglion neurons. Brain Res. 2013, 1509, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Obata, N.; Mizobuchi, S.; Itano, Y.; Matsuoka, Y.; Kaku, R.; Tomotsuka, N.; Morita, K.; Kanzaki, H.; Ouchida, M.; Yokoyama, M. Decoy strategy targeting the brain-derived neurotrophic factor exon I to attenuate tactile allodynia in the neuropathic pain model of rats. Biochem. Biophys. Res. Commun. 2011, 408, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Rana, D.G.; Patel, A.K.; Joshi, C.G.; Jhala, M.K.; Goyal, R.K. Alteration in the expression of exon IIC transcripts of brain-derived neurotrophic factor gene by simvastain in chronic mild stress in mice: A possible link with dopaminergic pathway. Can. J. Physiol. Pharmacol. 2014, 92, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Salerno, K.M.; Jing, X.; Diges, C.M.; Cornuet, P.K.; Glorioso, J.C.; Albers, K.M. Sox11 modulates brain-derived neurotrophic factor expression in an exon promoter-specific manner. J. Neurosci. Res. 2012, 90, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Ranney, A.; Petro, M.S. Resveratrol protects spatial learning in middle-aged C57BL/6 mice from effects of ethanol. Behav. Pharmacol. 2009, 20, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yang, Y.; Gao, L.; Zhao, J.; Cai, Y.; Huang, J.; Jing, S.; Bao, X.; Wang, Y.; Gao, J.; et al. Protective effects of resveratrol on the inhibition of hippocampal neurogenesis induced by ethanol during early postnatal life. Biochim. Biophys. Acta 2015, 1852, 1298–1310. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Zhang, J.; Liu, H.; Li, Z. The protective effects of resveratrol on schwann cells with toxicity induced by ethanol in vitro. Neurochem. Int. 2013, 63, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Quincozes-Santos, A.; Bobermin, L.D.; Tramontina, A.C.; Wartchow, K.M.; Tagliari, B.; Souza, D.O.; Wyse, A.T.; Goncalves, C.A. Oxidative stress mediated by NMDA, AMPA/KA channels in acute hippocampal slices: Neuroprotective effect of resveratrol. Toxicol. In Vitro 2014, 28, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Xu, H.; Sun, B.; Zhu, Z.; Zheng, D.; Li, X. Enhanced neuroprotective effects of resveratrol delivered by nanoparticles on hydrogen peroxide-induced oxidative stress in rat cortical cells culture. Mol. Pharm. 2013, 10, 2045–2053. [Google Scholar] [CrossRef] [PubMed]
- Porquet, D.; Grinan-Ferre, C.; Ferrer, I.; Camins, A.; Sanfeliu, C.; del Valle, J.; Pallas, M. Neuroprotective role of trans-resveratrol in a murine model of familial Alzheimer’s disease. J. Alzheimer’s Dis. JAD 2014, 42, 1209–1220. [Google Scholar]
- Wang, L.M.; Wang, Y.J.; Cui, M.; Luo, W.J.; Wang, X.J.; Barber, P.A.; Chen, Z.Y. A dietary polyphenol resveratrol acts to provide neuroprotection in recurrent stroke models by regulating AMPK and SIRT1 signaling, thereby reducing energy requirements during ischemia. Eur. J. Neurosci. 2013, 37, 1669–1681. [Google Scholar] [CrossRef] [PubMed]
- Pang, C.; Cao, L.; Wu, F.; Wang, L.; Wang, G.; Yu, Y.; Zhang, M.; Chen, L.; Wang, W.; Chen, L.; et al. The effect of trans-resveratrol on post-stroke depression via regulation of hypothalamus-pituitary-adrenal axis. Neuropharmacology 2015, 97, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.H.; Madhana, R.M.; Athira, K.V.; Kasala, E.R.; Bodduluru, L.N.; Pitta, S.; Mahareddy, J.R.; Lahkar, M. Resveratrol ameliorates depressive-like behavior in repeated corticosterone-induced depression in mice. Steroids 2015, 101, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Cheon, S.Y.; Jung, W.; Lee, W.T.; Lee, J.E. Resveratrol induces the expression of interleukin-10 and brain-derived neurotrophic factor in BV 2 microglia under hypoxia. Int. J. Mol. Sci. 2014, 15, 15512–15529. [Google Scholar] [CrossRef] [PubMed]
- Park, H.R.; Kong, K.H.; Yu, B.P.; Mattson, M.P.; Lee, J. Resveratrol inhibits the proliferation of neural progenitor cells and hippocampal neurogenesis. J. Biol. Chem. 2012, 287, 42588–42600. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Zhang, W.; Li, H.; Chen, C.; Liu, H.; Li, Z. Neuroprotective effects of resveratrol on embryonic dorsal root ganglion neurons with neurotoxicity induced by ethanol. Food Chem. Toxicol. 2013, 55, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, C.K.; Lavoie, H.A.; Dipette, D.J.; Singh, U.S. Resveratrol restores Nrf2 level and prevents ethanol-induced toxic effects in the cerebellum of a rodent model of fetal alcohol spectrum disorders. Mol. Pharmacol. 2011, 80, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.W.; Mooney, S.M. Chronic exposure to ethanol alters neurotrophin content in the basal forebrain-cortex system in the mature rat: Effects on autocrine-paracrine mechanisms. J. Neurobiol. 2004, 60, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Ceccanti, M.; Mancinelli, R.; Tirassa, P.; Laviola, G.; Rossi, S.; Romeo, M.; Fiore, M. Early exposure to ethanol or red wine and long-lasting effects in aged mice. A study on nerve growth factor, brain-derived neurotrophic factor, hepatocyte growth factor, and vascular endothelial growth factor. Neurobiol. Aging 2012, 33, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Heaton, M.B.; Mitchell, J.J.; Paiva, M.; Walker, D.W. Ethanol-induced alterations in the expression of neurotrophic factors in the developing rat central nervous system. Dev. Brain Res. 2000, 121, 97–107. [Google Scholar] [CrossRef]
- Madhyastha, S.; Sekhar, S.; Rao, G. Resveratrol improves postnatal hippocampal neurogenesis and brain derived neurotrophic factor in prenatally stressed rats. Int. J. Dev. Neurosci. 2013, 31, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gu, J.; Wang, X.; Xie, K.; Luan, Q.; Wan, N.; Zhang, Q.; Jiang, H.; Liu, D. Antidepressant-like activity of resveratrol treatment in the forced swim test and tail suspension test in mice: The HPA axis, BDNF expression and phosphorylation of ERK. Pharmacol. Biochem. Behav. 2013, 112, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Fleige, S.; Pfaffl, M.W. RNA integrity and the effect on the real-time qRT-PCR performance. Mol. Asp. Med. 2006, 27, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Mashayekhi, F.J.; Rasti, M.; Rahvar, M.; Mokarram, P.; Namavar, M.R.; Owji, A.A. Expression levels of the BDNF gene and histone modifications around its promoters in the ventral tegmental area and locus ceruleus of rats during forced abstinence from morphine. Neurochem. Res. 2012, 37, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. Quantification Strategies in Real-Time PCR. In a–z of Quantitative PCR; Bustin, S.A., Ed.; International University Line: La Jolla, CA, USA, 2004; pp. 87–112. [Google Scholar]
- Kobayashi, H.; Yokoyama, M.; Matsuoka, Y.; Omori, M.; Itano, Y.; Kaku, R.; Morita, K.; Ichikawa, H. Expression changes of multiple brain-derived neurotrophic factor transcripts in selective spinal nerve ligation model and complete freund’s adjuvant model. Brain Res. 2008, 1206, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz, M.; Emamghoreishi, M.; Nekooeian, A.A.; Warsh, J.J.; Zare, H.R. Increased bcl-2 protein levels in rat primary astrocyte culture following chronic lithium treatment. Iran. J. Med. Sci. 2013, 38, 255–262. [Google Scholar] [PubMed]
- Shafiee, S.M.; Seghatoleslam, A.; Nikseresht, M.; Hosseini, S.V.; Alizadeh-Naeeni, M.; Safaei, A.; Owji, A.A. Expression status of UBE2Q2 in colorectal primary tumors and cell lines. Iran. J. Med. Sci. 2014, 39, 196–202. [Google Scholar] [PubMed]
- Ghavami, S.; Yeganeh, B.; Stelmack, G.L.; Kashani, H.H.; Sharma, P.; Cunnington, R.; Rattan, S.; Bathe, K.; Klonisch, T.; Dixon, I.M.; et al. Apoptosis, autophagy and ER stress in mevalonate cascade inhibition-induced cell death of human atrial fibroblasts. Cell Death Dis. 2012, 3, e330. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shojaei, S.; Ghavami, S.; Panjehshahin, M.R.; Owji, A.A. Effects of Ethanol on the Expression Level of Various BDNF mRNA Isoforms and Their Encoded Protein in the Hippocampus of Adult and Embryonic Rats. Int. J. Mol. Sci. 2015, 16, 30422-30437. https://doi.org/10.3390/ijms161226242
Shojaei S, Ghavami S, Panjehshahin MR, Owji AA. Effects of Ethanol on the Expression Level of Various BDNF mRNA Isoforms and Their Encoded Protein in the Hippocampus of Adult and Embryonic Rats. International Journal of Molecular Sciences. 2015; 16(12):30422-30437. https://doi.org/10.3390/ijms161226242
Chicago/Turabian StyleShojaei, Shahla, Saeid Ghavami, Mohammad Reza Panjehshahin, and Ali Akbar Owji. 2015. "Effects of Ethanol on the Expression Level of Various BDNF mRNA Isoforms and Their Encoded Protein in the Hippocampus of Adult and Embryonic Rats" International Journal of Molecular Sciences 16, no. 12: 30422-30437. https://doi.org/10.3390/ijms161226242
APA StyleShojaei, S., Ghavami, S., Panjehshahin, M. R., & Owji, A. A. (2015). Effects of Ethanol on the Expression Level of Various BDNF mRNA Isoforms and Their Encoded Protein in the Hippocampus of Adult and Embryonic Rats. International Journal of Molecular Sciences, 16(12), 30422-30437. https://doi.org/10.3390/ijms161226242