
Chloroplast-Expressed MSI-99 in Tobacco Improves Disease Resistance and Displays Inhibitory Effect against Rice Blast Fungus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Construction of the Expression Vector for Tobacco Chloroplast Transformation

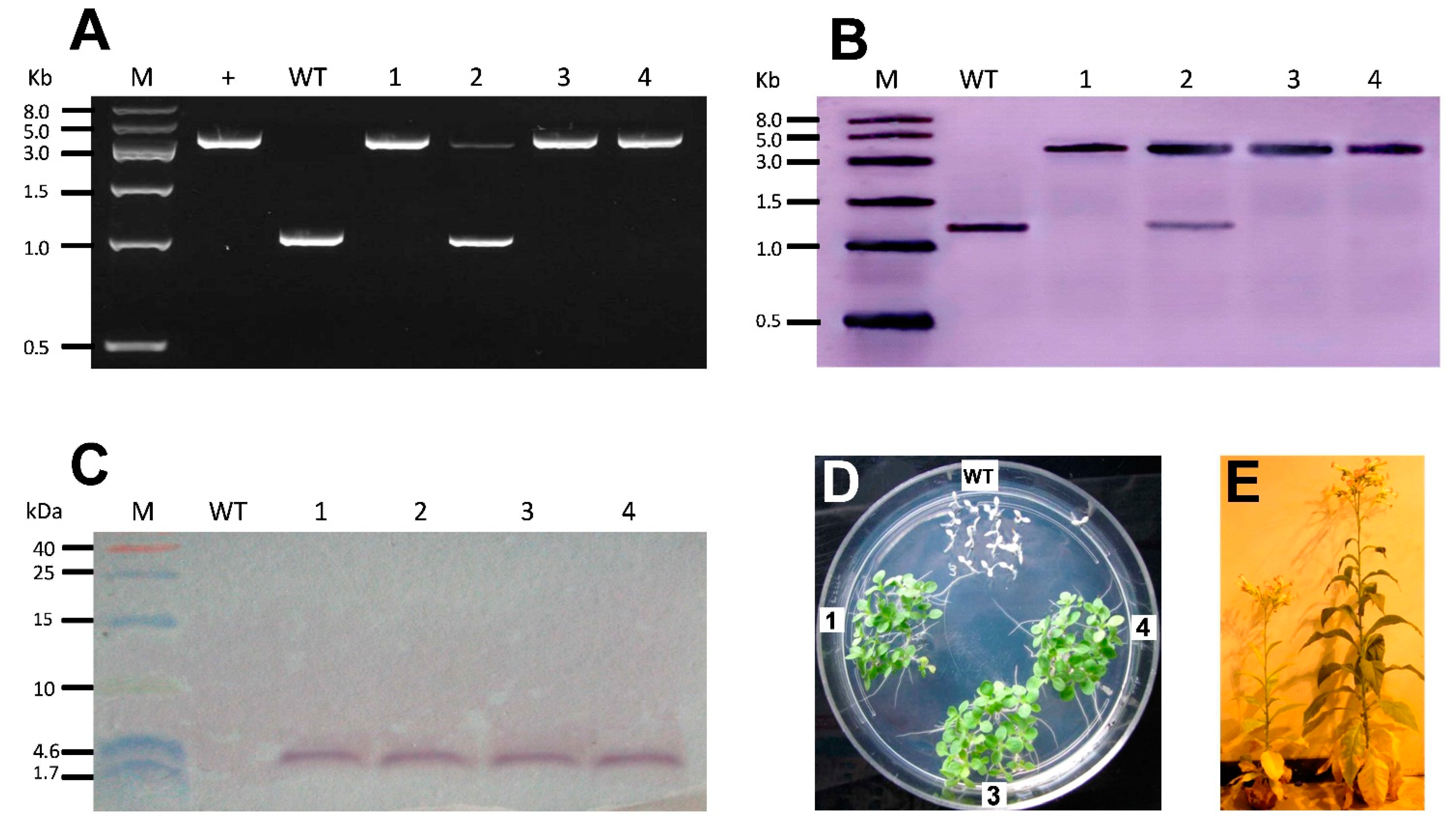
2.2. Selection of Homoplastomic Plants, Molecular Testing and Antibiotic Resistance of T1 Offspring

2.3. Quantification of MSI-99 Expression in T1 Generation

2.4. In Vivo Assessment of Green Fluorescent Protein (GFP) Expression

2.5. Resistance to the Fungal Disease Caused by Alternaria alternata and in Vitro Antifungal Activity of MSI-99 against Rice Blast Pathogens

2.6. In Plant Resistance to Rice Blast Magnaporthe oryzae

3. Experimental Section
3.1. Plant Materials and Growth Conditions
3.2. Construction of the Tobacco Chloroplast Transformation Vector
3.3. Chloroplast Transformation and Homoplastomic Selection
3.4. PCR Amplification
3.5. Southern Blot Analysis
3.6. Tricine-SDS-PAGE, Western Blot Analysis and ELISA Assay
3.7. Assessment of GFP Expression
3.8. In Planta Assay for Fungal Disease Resistance in Transgenic Tobacco and in Vitro Evaluation of Antimicrobial Activity against the Growth of E. coli
3.9. In Vitro and in Vivo Bioassays of Antifungal Activity of MSI-99
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Keymanesh, K.; Soltani, S.; Sardari, S. Application of antimicrobial peptides in agriculture and food industry. World J. Microbiol. Biotechnol. 2009, 25, 933–944. [Google Scholar] [CrossRef]
- Kasetsomboon, T.; Kate-Ngam, S.; Sriwongchai, T.; Zhou, B.; Jantasuriyarat, C. Sequence variation of avirulence gene AVR-Pita1 in rice blast fungus, Magnaporthe oryzae. Mycol. Prog. 2013, 12, 617–628. [Google Scholar] [CrossRef]
- Xiang, K.; Li, W.T.; Chen, X.W.; Yuan, G.S.; Chen, W.L.; Zhang, Z.M.; Shen, Y.O.; Lin, H.J.; Pan, G.T. Clustering and expression analysis of chitinases in maize and rice. J. Life Sci. 2013, 7, 244–251. [Google Scholar]
- Kouzai, Y.; Mochizuki, S.; Saito, A.; Ando, A.; Minami, E.; Nishizawa, Y. Expression of a bacterial chitosanase in rice plants improves disease resistance to the rice blast fungus Magnaporthe oryzae. Plant Cell Rep. 2012, 31, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Chen, S.; Yang, J.; Zhou, S.; Zeng, L.; Han, J.; Wang, L.; Pan, Q. The identification of Pi50 (t), a new member of the rice blast resistance Pi2/Pi9 multigene family. Theor. Appl. Genet. 2012, 124, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.V.; Yedery, R.D.; Aranha, C. Antimicrobial peptides: Premises and promises. Int. J. Antimicrob. Agents 2004, 24, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Hayes, B.M.; Anderson, M.A.; Traven, A.; van der Weerden, N.L.; Bleackley, M.R. Activation of stress signalling pathways enhances tolerance of fungi to chemical fungicides and antifungal proteins. Cell. Mol. Life Sci. 2014, 71, 2651–2666. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lawrence, C.B.; Xing, H.Y.; Babbitt, R.A.; Bass, W.T.; Maiti, I.B.; Everett, N.P. Enhanced disease resistance conferred by expression of an antimicrobial magainin analog in transgenic tobacco. Planta 2001, 212, 635–639. [Google Scholar] [CrossRef] [PubMed]
- DeGray, G.; Rajasekaran, K.; Smith, F.; Sanford, J.; Daniell, H. Expression of an antimicrobial peptide via the chloroplast genome to control phytopathogenic bacteria and fungi. Plant Physiol. 2001, 127, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Ganapathi, T.R.; Mukherjee, P.K.; Bapat, V.A. MSI-99, a magainin analogue, imparts enhanced disease resistance in transgenic tobacco and banana. Planta 2003, 216, 587–596. [Google Scholar] [PubMed]
- Alan, A.R.; Blowers, A.; Earle, E.D. Expression of a magainin-type antimicrobial peptide gene (MSI-99) in tomato enhances resistance to bacterial speck disease. Plant Cell Rep. 2004, 22, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.B.; Liu, S.P.; Zhu, Y.P.; Xie, C.; Jue, D.W.; Chen, M.; Kaleri, A.H.; Yang, Q. Expression of the MSI-99m gene in transgenic potato plants confers resistance to Phytophthora infestans and Ralstonia solanacearum. Plant Mol. Biol. Rep. 2013, 31, 418–424. [Google Scholar] [CrossRef]
- Sang, X.; Jue, D.; Yang, L.; Bai, X.; Chen, M.; Yang, Q. Genetic transformation of Brassica napus with MSI-99m gene increases resistance in transgenic plants to Sclerotinia sclerotiorum. Mol. Plant Breed. 2013, 4, 247–253. [Google Scholar]
- Clarke, J.L.; Daniell, H. Plastid biotechnology for crop production: Present status and future perspectives. Plant Mol. Biol. 2011, 76, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Daniell, H. Production of biopharmaceuticals and vaccines in plants via the chloroplast genome. Biotechnol. J. 2006, 1, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Grevich, J.J.; Daniell, H. Chloroplast genetic engineering: recent advances and future perspectives. Crit. Rev. Plant Sci. 2005, 24, 83–107. [Google Scholar] [CrossRef]
- Maliga, P. Plastid transformation in higher plants. Annu. Rev. Plant Biol. 2004, 55, 289–313. [Google Scholar] [CrossRef] [PubMed]
- Bock, R. Engineering chloroplasts for high-level foreign protein expression. In Chloroplast Biotechnology: Methods in Molecular Biology; Maliga, P., Ed.; Humana Press: New York, NY, USA, 2014; Volume 1132, pp. 93–106. [Google Scholar]
- Day, A.; Goldschmidt-Clermont, M. The chloroplast transformation toolbox: Selectable markers and marker removal. Plant Biotechnol. J. 2011, 9, 540–553. [Google Scholar] [CrossRef] [PubMed]
- Scotti, N.; Cardi, T. Transgene-induced pleiotropic effects in transplastomic plants. Biotechnol. Lett. 2014, 36, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Boman, A.; Sun, C.X.; Andersson, M.; Jörnvall, H.; Mutt, V.; Boman, H.G. Antibacterial peptides from pig intestine: Isolation of a mammalian cecropin. Proc. Natl. Acad. Sci. USA 1989, 86, 9159–9162. [Google Scholar] [CrossRef] [PubMed]
- Papo, N.; Shai, Y. Host defense peptides as new weapons in cancer treatment. Cell. Mol. Life Sci. 2005, 62, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Hajji, M.; Jellouli, K.; Hmidet, N.; Balti, R.; Sellami-Kamoun, A.; Nasri, M. A highly thermostable antimicrobial peptide from Aspergillus clavatus ES1: Biochemical and molecular characterization. J. Ind. Microbiol. Biotechnol. 2010, 37, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.K.; Lee, M.J.; Jung, H.G.; Go, H.J.; Kim, Y.J.; Park, N.G. Antimicrobial function of SHβAP, a novel hemoglobin β chain-related antimicrobial peptide, isolated from the liver of skipjack tuna, Katsuwonus pelamis. Fish Shellfish Immun. 2014, 37, 173–183. [Google Scholar] [CrossRef]
- Hou, B.K.; Zhou, Y.H.; Wan, L.H.; Zhang, Z.L.; Shen, G.F.; Chen, Z.H.; Hu, Z.M. Chloroplast transformation in oilseed rape. Transgenic Res. 2003, 12, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Lössl, A.G.; Waheed, M.T. Chloroplast-derived vaccines against human diseases: Achievements, challenges and scopes. Plant Biotechnol. J. 2011, 9, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Oey, M.; Lohse, M.; Kreikemeyer, B.; Bock, R. Exhaustion of the chloroplast protein synthesis capacity by massive expression of a highly stable protein antibiotic. Plant J. 2009, 57, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.; Bock, R. High-level expression of a suite of thermostable cell wall-degrading enzymes from the chloroplast genome. Plant Mol. Biol. 2011, 76, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Bock, R. Genetic engineering of the chloroplast: Novel tools and new applications. Curr. Opin. Biotechnol. 2014, 26, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.Y.; Liu, Y.Z.; Lin, C.J.; Wang, Y.P.; Cai, Q.A.; Dong, Y.S.; Xing, S.C. Transformation of alfalfa chloroplasts and expression of green fluorescent protein in a forage crop. Biotechnol. Lett. 2011, 33, 2487–2494. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.W.; Lin, C.C.; Chen, J.J.; Tseng, M.J. Stable chloroplast transformation in cabbage (Brassica oleracea L. var. capitata L.) by particle bombardment. Plant Cell Rep. 2007, 26, 1733–1744. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.; Rajasekaran, K.; Jaynes, J.; Cleveland, T. Transgenic expression of a gene encoding a synthetic antimicrobial peptide results in inhibition of fungal growth in vitro and in planta. Plant Sci. 2000, 154, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, K.; Vera Cruz, C.M.; Gruissem, W.; Bhullar, N.K. Large scale germplasm screening for identification of novel rice blast resistance sources. Front. Plant Sci. 2014, 5, 505. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-P.; Wei, Z.-Y.; Zhang, Y.-Y.; Lin, C.-J.; Zhong, X.-F.; Wang, Y.-L.; Ma, J.-Y.; Ma, J.; Xing, S.-C. Chloroplast-Expressed MSI-99 in Tobacco Improves Disease Resistance and Displays Inhibitory Effect against Rice Blast Fungus. Int. J. Mol. Sci. 2015, 16, 4628-4641. https://doi.org/10.3390/ijms16034628
Wang Y-P, Wei Z-Y, Zhang Y-Y, Lin C-J, Zhong X-F, Wang Y-L, Ma J-Y, Ma J, Xing S-C. Chloroplast-Expressed MSI-99 in Tobacco Improves Disease Resistance and Displays Inhibitory Effect against Rice Blast Fungus. International Journal of Molecular Sciences. 2015; 16(3):4628-4641. https://doi.org/10.3390/ijms16034628
Chicago/Turabian StyleWang, Yun-Peng, Zheng-Yi Wei, Yu-Ying Zhang, Chun-Jing Lin, Xiao-Fang Zhong, Yue-Lin Wang, Jing-Yong Ma, Jian Ma, and Shao-Chen Xing. 2015. "Chloroplast-Expressed MSI-99 in Tobacco Improves Disease Resistance and Displays Inhibitory Effect against Rice Blast Fungus" International Journal of Molecular Sciences 16, no. 3: 4628-4641. https://doi.org/10.3390/ijms16034628