
Development of Small RNA Delivery Systems for Lung Cancer Therapy

Abstract
:1. Introduction
2. The Development of siRNA-Based Therapeutics for Lung Cancer Treatment

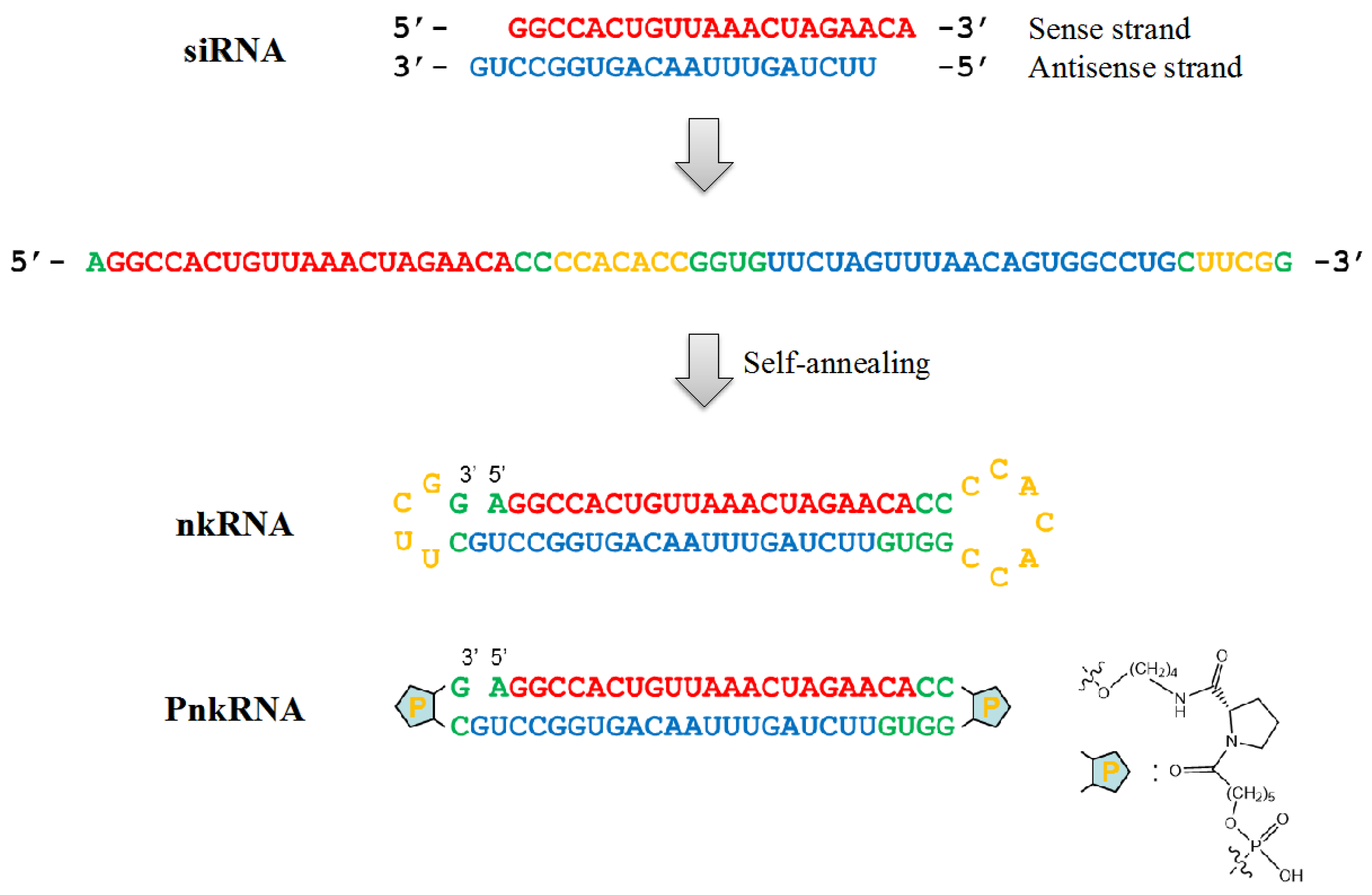{kind=link}
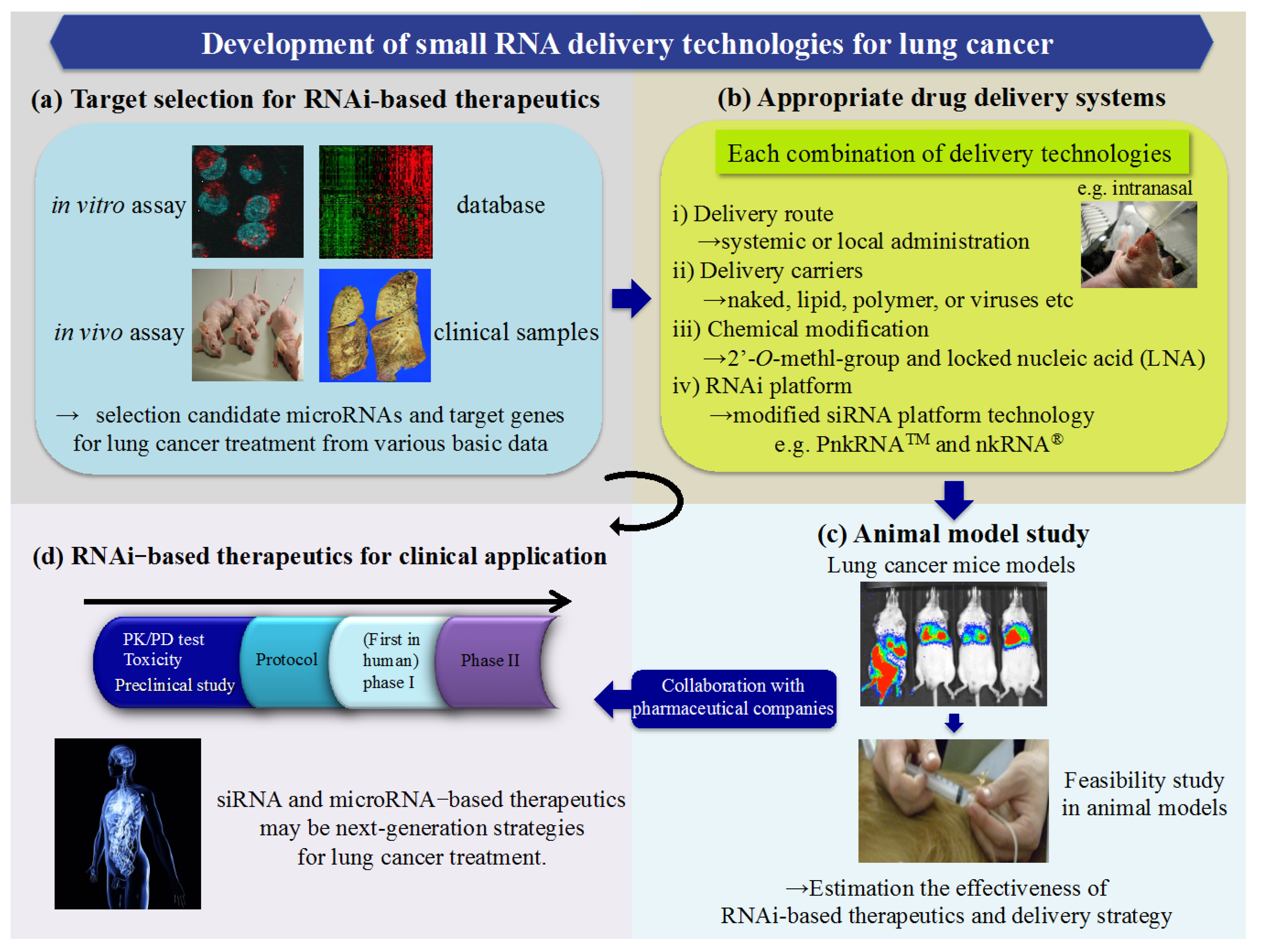{kind=link}
| Drug | Target Gene | Delivery Methods | Disease | Vehicle | Phase | Year |
|---|---|---|---|---|---|---|
| CALAA-01 | RRM2 | Intravenous injection | Solid tumors | Cyclodextrin nanoparticle | I | 2008 |
| TKM 080301 | PLK1 | Intravenous injection | Solid tumors with liver involvement | Lipid nanoparticle (LNP) | I/II | 2010 |
| ALN-VSP02 | KSP/VEGF | Intravenous injection | Solid tumors with liver involvement | Lipid nanoparticle (LNP) | I | 2009 |
| Atu027 | PKN3 | Intravenous injection | Solid tumors | Lipid nanoparticle (LNP) | I | 2009 |
| siG12D LODER | KRAS-G12D | EUS biopsy needle | Pancreatic ductal adenocarcinoma | LODER polymer | II | 2011 |
| siRNA-EphA2-DOPC | EphA2 | Intravenous injection | Solid tumors | DOPC | I | 2012 |
| Target Gene | Administration | Type of siRNA Delivery | References |
|---|---|---|---|
| RPN2 | Intratracheal | Naked nucleic acids | [20] |
| C7orf24 | Intratumoral | Naked nucleic acids | [27] |
| Mcl1 | Intratracheal | Ethylphosphocholine-based lipoplexes | [28] |
| CD31 | Intravenous | AtuFECT01 lipoplexes | [29] |
| IGF-1R | Intravenous | Magnetic lipoplexes | [56] |
| Survivin | Intravenous | Liposomes | [31] |
| MRP1 | Inhalation | Liposomes | [34] |
| Luciferase | Inhalation | Chitosan | [36] |
| Bcl-2 | Intravenous | Cationic bovine serum albumin | [38] |
| Akt1 | Inhalation | Glycerol propoxylate triacrylate-spermine | [39,40] |
| NPT2b | Inhalation | Glycerol propoxylate triacrylate-spermine | [41] |
| MDM2 | Intravenous | Poly(methacryloyloxy ethyl phosphorylcholine)-block-poly(diisopropanolamine ethyl methacrylate) (PDMA-b-PDPA) | [42] |
| STAT3 | Intraperitoneal | PEI and poly-l-lactic-co-glycolic acid (PLGA) | [44] |
| MDM2, c-myc, VEGF | Intravenous | LCP | [43] |
| VEGF | Intravenous | LCP | [46] |
| c-Myc | Intratracheal | Arginine-glycine-aspartic acid (RGD) gold nanoparticles | [45] |
| MRP1 and Bcl-2 | Inhalation | Lutein hormone releasing hormone (LHRH)-modified mesoporous silica nanoparticles (MSN) | [35] |
3. The Development of microRNA-Based Therapeutics for Lung Cancer Treatment
| microRNA | Modulation Strategy | Diseases | Status | Company |
|---|---|---|---|---|
| miR-10b | Inhibition | Glioblastoma | Preclinical | Regulus Therapeutics |
| miR-21 | Inhibition | Hepatocellular carcinoma | Preclinical | Regulus Therapeutics |
| miR-155 | Inhibition | Hematological malignancies | Preclinical | miRagen Therapeutics |
| miR-221 | Inhibition | Hepatocellular carcinoma | Preclinical | Sanofi |
| let-7 | Replacement | Lung cancer | Preclinical | Mirna Therapeutics |
| miR-16 | Replacement | Cancer | Preclinical | Mirna Therapeutics |
| miR-34 | Replacement | Hepatocellular carcinoma | Phase I | Mirna Therapeutics |
| miRNA | Administration | Modulation Strategy | Delivery Technology | References |
|---|---|---|---|---|
| let-7 | Intranasal | Replacement | Adenoviruses | [67] |
| Intravenous | Replacement | Neutral liposomes | [61] | |
| Intratracheal | Replacement | Lentiviruses | [65] | |
| miR-7 | Intratumoral | Replacement | Cationic liposomes | [69] |
| miR-29b | Intravenous | Replacement | Cationic liposomes | [70] |
| miR-34a | Intratumoral | Replacement | Neutral liposomes | [71] |
| miR-145 | Intratumoral | Replacement | PEI | [72] |
| miR-150 | Intratumoral | Inhibition | Cationic liposomes | [73] |
| miR-200c | Intravenous | Replacement | Liposomes (NOV340) | [74] |
4. A Novel RNAi Platform for Lung Cancer Treatment

5. Conclusions

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zamore, P.D.; Tuschl, T.; Sharp, P.A.; Bartel, D.P. RNAi: Double-stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals. Cell 2000, 101, 25–33. [Google Scholar] [PubMed]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Elbashir, S.M.; Harborth, J.; Lendeckel, W.; Yalcin, A.; Weber, K.; Tuschl, T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 2001, 411, 494–498. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, E.; Caudy, A.A.; Hammond, S.M.; Hannon, G.J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 2001, 409, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, L.; Rossi, J.J. RNAi therapeutics: Principles, prospects and challenges. Adv. Drug Deliv. Rev. 2007, 59, 75–86. [Google Scholar] [CrossRef] [PubMed]
- De Fougerolles, A.; Vornlocher, H.P.; Maraganore, J.; Lieberman, J. Interfering with disease: A progress report on siRNA-based therapeutics. Nat. Rev. Drug Deliv. 2007, 6, 443–453. [Google Scholar]
- Check, E. A crucial test. Nat. Med. 2005, 11, 243–244. [Google Scholar] [CrossRef] [PubMed]
- DeVincenzo, J.; Cehelsky, J.E.; Alvarez, R.; Elbashir, S.; Harborth, J.; Toudjarska, I.; Nechev, L.; Murugaiah, V.; van Vliet, A.; Vaishnaw, A.K.; et al. Evaluation of the safety, tolerability and pharmacokinetics of ALN-RSV01, a novel RNAi antiviral therapeutic directed against respiratory syncytial virus (RSV). Antivir. Res. 2008, 77, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Boudreau, R.L.; Martins, I.; Davidson, B.L. Artificial microRNAs as siRNA shuttles: Improved safety as compared to shRNAs in vitro and in vivo. Mol. Ther. 2009, 17, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Burchard, J.; Jackson, A.L.; Malkov, V.; Needham, R.H.; Tan, Y.; Bartz, S.R.; Dai, H.; Sachs, A.B.; Linsley, P.S. MicroRNA-like off-target transcript regulation by siRNAs is species specific. RNA 2009, 15, 308–315. [Google Scholar] [CrossRef] [PubMed]
- De Veer, M.J.; Sledz, C.A.; Williams, B.R. Detection of foreign RNA: Implications for RNAi. Immunol. Cell Biol. 2005, 83, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.; Jung, S.; Keller, S.; Gregory, R.I.; Diederichs, S. Many roads to maturity: MicroRNA biogenesis pathways and their regulation. Nat. Cell Biol. 2009, 11, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Croce, C.M. Causes and consequences of microRNA dysregulation in cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Pasquinelli, A.E.; Hunter, S.; Bracht, J. MicroRNAs: A developing story. Curr. Opin. Genet. Dev. 2005, 15, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, S.S.; Owonikoko, T.K.; Khuri, F.R. Lung cancer: New biological insights and recent therapeutic advances. CA Cancer J. Clin. 2011, 61, 91–112. [Google Scholar] [CrossRef] [PubMed]
- Pao, W.; Girard, N. New driver mutations in non-small-cell lung cancer. Lancet Oncol. 2011, 12, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Turrisi, A.T.; Sherman, C.A. The treatment of limited small cell lung cancer: A report of the progress made and future prospects. Eur. J. Cancer 2002, 38, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Kleinman, M.E.; Yamada, K.; Takeda, A.; Chandrasekaran, V.; Nozaki, M.; Baffi, J.Z.; Albuquerque, R.J.; Yamasaki, S.; Itaya, M.; Pan, Y.; et al. Sequence- and target-independent angiogenesis suppression by siRNA via TLR3. Nature 2008, 452, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Takeshita, F.; Mizutani, T.; Ohgi, T.; Kuwano, K.; Ochiya, T. A novel platform to enable inhaled naked RNAi medicine for lung cancer. Sci. Rep. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Davidson, B.L.; McCray, P.B., Jr. Current prospects for RNA interference-based therapies. Nat. Rev. Genet. 2011, 12, 329–340. [Google Scholar] [CrossRef] [PubMed]
- DeVincenzo, J.; Lambkin-Williams, R.; Wilkinson, T.; Cehelsky, J.; Nochur, S.; Walsh, E.; Meyers, R.; Gollob, J.; Vaishnaw, A. A randomized, double-blind, placebo-controlled study of an RNAi-based therapy directed against respiratory syncytial virus. Proc. Natl. Acad. Sci. USA 2010, 107, 8800–8805. [Google Scholar] [CrossRef] [PubMed]
- Bouchie, A. Companies in footrace to deliver RNAi. Nat. Biotechnol. 2012, 30, 1154–1157. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Takeshita, F.; Kuwano, K.; Ochiya, T. RNAi therapeutic platforms for lung diseases. Pharmaceuticals 2013, 6, 223–250. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.E.; Zuckerman, J.E.; Choi, C.H.; Seligson, D.; Tolcher, A.; Alabi, C.A.; Yen, Y.; Heidel, J.D.; Ribas, A. Evidence of RNAi in humans from systemically administered siRNA via targeted nanoparticles. Nature 2010, 464, 1067–1070. [Google Scholar] [CrossRef] [PubMed]
- Landen, C.N., Jr.; Chavez-Reyes, A.; Bucana, C.; Schmandt, R.; Deavers, M.T.; Lopez-Berestein, G.; Sood, A.K. Therapeutic EphA2 gene targeting in vivo using neutral liposomal small interfering RNA delivery. Cancer Res. 2005, 65, 6910–6918. [Google Scholar] [CrossRef] [PubMed]
- Hama, S.; Arata, M.; Nakamura, I.; Kasetani, T.; Itakura, S.; Tsuchiya, H.; Yoshiki, T.; Kogure, K. Prevention of tumor growth by needle-free jet injection of anti-C7orf24 siRNA. Cancer Gene Ther. 2012, 19, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Shim, G.; Choi, H.W.; Lee, S.; Choi, J.; Yu, Y.H.; Park, D.E.; Choi, Y.; Kim, C.W.; Oh, Y.K. Enhanced intrapulmonary delivery of anticancer siRNA for lung cancer therapy using cationic ethylphosphocholine-based nanolipoplexes. Mol. Ther. 2013, 21, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Fehring, V.; Schaeper, U.; Ahrens, K.; Santel, A.; Keil, O.; Eisermann, M.; Giese, K.; Kaufmann, J. Delivery of therapeutic siRNA to the lung endothelium via novel Lipoplex formulation DACC. Mol. Ther. 2014, 22, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Dong, A.Q.; Kong, M.J.; Ma, Z.Y.; Qian, J.F.; Xu, X.H. Down-regulation of IGF-IR using small, interfering, hairpin RNA (siRNA) inhibits growth of human lung cancer cell line A549 in vitro and in nude mice. Cell Biol. Int. 2007, 31, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Liu, S.; Zhang, J.; Zhang, S.; Cheng, L.; Li, C.; Zhang, X.; Dail, L.; Fan, P.; Dai, L.; et al. Enhancement of cisplatin sensitivity in lung cancer xenografts by liposome-mediated delivery of the plasmid expressing small hairpin RNA targeting Survivin. J. Biomed. Nanotechnol. 2012, 8, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Kedinger, V.; Meulle, A.; Zounib, O.; Bonnet, M.E.; Gossart, J.B.; Benoit, E.; Messmer, M.; Shankaranarayanan, P.; Behr, J.P.; Erbacher, P.; et al. Sticky siRNAs targeting survivin and cyclin B1 exert an antitumoral effect on melanoma subcutaneous xenografts and lung metastases. BMC Cancer 2013, 13. [Google Scholar] [CrossRef]
- Bonnet, M.E.; Gossart, J.B.; Benoit, E.; Messmer, M.; Zounib, O.; Moreau, V.; Behr, J.P.; Lenne-Samuel, N.; Kedinger, V.; Meulle, A.; et al. Systemic delivery of sticky siRNAs targeting the cell cycle for lung tumor metastasis inhibition. J. Control. Release 2013, 170, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Mainelis, G.; Seshadri, S.; Garbuzenko, O.B.; Han, T.; Wang, Z.; Minko, T. Characterization and application of a nose-only exposure chamber for inhalation delivery of liposomal drugs and nucleic acids to mice. J. Aerosol Med. Pulm. Drug Deliv. 2013, 26, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Taratula, O.; Garbuzenko, O.B.; Chen, A.M.; Minko, T. Innovative strategy for treatment of lung cancer: Targeted nanotechnology-based inhalation co-delivery of anticancer drugs and siRNA. J. Drug Targets 2011, 19, 900–914. [Google Scholar] [CrossRef]
- Okuda, T.; Kito, D.; Oiwa, A.; Fukushima, M.; Hira, D.; Okamoto, H. Gene silencing in a mouse lung metastasis model by an inhalable dry small interfering RNA powder prepared using the supercritical carbon dioxide technique. Biol. Pharm. Bull. 2013, 36, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, Y.; Huang, L. Calcium phosphate nanoparticles with an asymmetric lipid bilayer coating for siRNA delivery to the tumor. J. Control. Release 2012, 158, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, Q.; Zhang, Z.; Gong, T.; Sun, X. Cationic bovine serum albumin based self-assembled nanoparticles as siRNA delivery vector for treating lung metastatic cancer. Small 2014, 10, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.L.; Hong, S.H.; Kim, Y.K.; Islam, M.A.; Kim, H.J.; Choi, Y.J.; Nah, J.W.; Lee, K.H.; Han, K.W.; Chae, C.; et al. Aerosol delivery of spermine-based poly(amino ester)/Akt1 shRNA complexes for lung cancer gene therapy. Int. J. Pharm. 2011, 420, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Kim, J.E.; Kim, Y.K.; Minai-Tehrani, A.; Shin, J.Y.; Kang, B.; Kim, H.J.; Cho, C.S.; Chae, C.; Jiang, H.L.; et al. Suppression of lung cancer progression by biocompatible glycerol triacrylate-spermine-mediated delivery of shAkt1. Int. J. Nanomed. 2012, 7, 2293–2306. [Google Scholar]
- Hong, S.H.; Minai-Tehrani, A.; Chang, S.H.; Jiang, H.L.; Lee, S.; Lee, A.Y.; Seo, H.W.; Chae, C.; Beck, G.R., Jr.; Cho, M.H. Knockdown of the sodium-dependent phosphate co-transporter 2b (NPT2b) suppresses lung tumorigenesis. PLoS One 2013, 8, e77121. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zou, Y.; Jiang, L.; Yin, Q.; He, X.; Chen, L.; Zhang, Z.; Gu, W.; Li, Y. Induction of apoptosis in non-small cell lung cancer by downregulation of MDM2 using pH-responsive PMPC-b-PDPA/siRNA complex nanoparticles. Biomaterials 2013, 34, 2738–2747. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, J.; Liu, F.; Huang, L. Systemic delivery of siRNA via LCP nanoparticle efficiently inhibits lung metastasis. Mol. Ther. 2012, 20, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Das, S.; Paul, A.; Samadder, A.; Bhattacharyya, S.S.; Khuda-Bukhsh, A.R. Assessment of drug delivery and anticancer potentials of nanoparticles-loaded siRNA targeting STAT3 in lung cancer, in vitro and in vivo. Toxicol. Lett. 2014, 225, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Conde, J.; Tian, F.; Hernandez, Y.; Bao, C.; Cui, D.; Janssen, K.P.; Ibarra, M.R.; Baptista, P.V.; Stoeger, T.; de la Fuente, J.M. In vivo tumor targeting via nanoparticle-mediated therapeutic siRNA coupled to inflammatory response in lung cancer mouse models. Biomaterials 2013, 34, 7744–7753. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Schwerbrock, N.M.; Rogers, A.B.; Kim, W.Y.; Huang, L. Codelivery of VEGF siRNA and gemcitabine monophosphate in a single nanoparticle formulation for effective treatment of NSCLC. Mol. Ther. 2013, 21, 1559–1569. [Google Scholar] [CrossRef] [PubMed]
- Dehousse, V.; Garbacki, N.; Colige, A.; Evrard, B. Development of pH-responsive nanocarriers using trimethylchitosans and methacrylic acid copolymer for siRNA delivery. Biomaterials 2010, 31, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, C.A.; Rice, K.G. Engineered nanoscaled polyplex gene delivery systems. Mol. Pharm. 2009, 6, 1277–1289. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Crawford, M.; Yu, B.; Mao, Y.; Nana-Sinkam, S.P.; Lee, L.J. MicroRNA delivery by cationic lipoplexes for lung cancer therapy. Mol. Pharm. 2011, 8, 1381–1389. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Guan, W.; Wang, D.; Zhang, Y.S.; Zeng, L.L.; Li, Z.P.; Wang, G.; Yang, Z.Z. Killing effect of Ad5/F35-APE1 siRNA recombinant adenovirus in combination with hematoporphrphyrin derivative-mediated photodynamic therapy on human nonsmall cell lung cancer. BioMed Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Cun, Y.; Li, M.; Qing, Y.; Jin, F.; Zhong, Z.; Dai, N.; Qian, C.; Sui, J.; Wang, D. Human apurinic/apyrimidinic endonuclease siRNA inhibits the angiogenesis induced by X-ray irradiation in lung cancer cells. Int. J. Med. Sci. 2013, 10, 870–882. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Cui, J.W.; Li, W.; Cai, L.; Song, W.; Wang, G.J. Silencing of the COPS3 gene by siRNA reduces proliferation of lung cancer cells most likely via induction of cell cycle arrest and apoptosis. Asian Pac. J. Cancer Prev. 2012, 13, 1043–1048. [Google Scholar] [CrossRef] [PubMed]
- Lian, Y.X.; Chen, R.; Xu, Y.H.; Peng, C.L.; Hu, H.C. Effect of protein-tyrosine phosphatase 4A3 by small interfering RNA on the proliferation of lung cancer. Gene 2012, 511, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yan, X.; Liu, N.; Zhou, J.; Liu, J.; Pang, H.; Cao, J.; Liu, Y.; Wang, Y.; Liu, L.; et al. Lentivirus-delivered ZEB-1 small interfering RNA inhibits lung adenocarcinoma cell growth in vitro and in vivo. J. Cancer Res. Clin. Oncol. 2012, 138, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Caino, M.C.; Lopez-Haber, C.; Kim, J.; Mochly-Rosen, D.; Kazanietz, M.G. Proteins kinase Cvarepsilon is required for non-small cell lung carcinoma growth and regulates the expression of apoptotic genes. Oncogene 2012, 31, 2593–2600. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ding, C.; Kong, M.; Dong, A.; Qian, J.; Jiang, D.; Shen, Z. Tumor-targeting magnetic lipoplex delivery of short hairpin RNA suppresses IGF-1R overexpression of lung adenocarcinoma A549 cells in vitro and in vivo. Biochem. Biophys. Res. Commun. 2011, 410, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from repression to activation: MicroRNAs can up-regulate translation. Science 2007, 318, 1931–1934. [Google Scholar] [CrossRef] [PubMed]
- Bruno, I.G.; Karam, R.; Huang, L.; Bhardwaj, A.; Lou, C.H.; Shum, E.Y.; Song, H.W.; Corbett, M.A.; Gifford, W.D.; Gecz, J.; et al. Identification of a microRNA that activates gene expression by repressing nonsense-mediated RNA decay. Mol. Cell 2011, 42, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Krutzfeldt, J.; Rajewsky, N.; Braich, R.; Rajeev, K.G.; Tuschl, T.; Manoharan, M.; Stoffel, M. Silencing of microRNAs in vivo with “antagomirs”. Nature 2005, 438, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Misso, G.; di Martino, M.T.; de Rosa, G.; Farooqi, A.A.; Lombardi, A.; Campani, V.; Zarone, M.R.; Gulla, A.; Tagliaferri, P.; Tassone, P.; et al. MiR-34: A new weapon against cancer? Mol. Ther. Nucleic Acids 2014, 3. [Google Scholar] [CrossRef]
- Trang, P.; Wiggins, J.F.; Daige, C.L.; Cho, C.; Omotola, M.; Brown, D.; Weidhaas, J.B.; Bader, A.G.; Slack, F.J. Systemic delivery of tumor suppressor microRNA mimics using a neutral lipid emulsion inhibits lung tumors in mice. Mol. Ther. 2011, 19, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Yanaihara, N.; Caplen, N.; Bowman, E.; Seike, M.; Kumamoto, K.; Yi, M.; Stephens, R.M.; Okamoto, A.; Yokota, J.; Tanaka, T.; et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell 2006, 9, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Takamizawa, J.; Konishi, H.; Yanagisawa, K.; Tomida, S.; Osada, H.; Endoh, H.; Harano, T.; Yatabe, Y.; Nagino, M.; Nimura, Y.; et al. Reduced expression of the let-7 microRNAs in human lung cancers in association with shortened postoperative survival. Cancer Res. 2004, 64, 3753–3756. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.D.; Esquela-Kerscher, A.; Stefani, G.; Byrom, M.; Kelnar, K.; Ovcharenko, D.; Wilson, M.; Wang, X.; Shelton, J.; Shingara, J.; et al. The let-7 microRNA represses cell proliferation pathways in human cells. Cancer Res. 2007, 67, 7713–7722. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.S.; Erkeland, S.J.; Pester, R.E.; Chen, C.Y.; Ebert, M.S.; Sharp, P.A.; Jacks, T. Suppression of non-small cell lung tumor development by the let-7 microRNA family. Proc. Natl. Acad. Sci. USA 2008, 105, 3903–3908. [Google Scholar] [CrossRef] [PubMed]
- He, X.Y.; Chen, J.X.; Zhang, Z.; Li, C.L.; Peng, Q.L.; Peng, H.M. The let-7a microRNA protects from growth of lung carcinoma by suppression of k-Ras and c-Myc in nude mice. J. Cancer Res. Clin. Oncol. 2010, 136, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Esquela-Kerscher, A.; Trang, P.; Wiggins, J.F.; Patrawala, L.; Cheng, A.; Ford, L.; Weidhaas, J.B.; Brown, D.; Bader, A.G.; Slack, F.J. The let-7 microRNA reduces tumor growth in mouse models of lung cancer. Cell Cycle 2008, 7, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Trang, P.; Medina, P.P.; Wiggins, J.F.; Ruffino, L.; Kelnar, K.; Omotola, M.; Homer, R.; Brown, D.; Bader, A.G.; Weidhaas, J.B.; et al. Regression of murine lung tumors by the let-7 microRNA. Oncogene 2010, 29, 1580–1587. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.; Takigawa, N.; Ito, S.; Kashihara, H.; Ichihara, E.; Yasuda, T.; Shimizu, K.; Tanimoto, M.; Kiura, K. Liposomal delivery of microRNA-7-expressing plasmid overcomes epidermal growth factor receptor tyrosine kinase inhibitor-resistance in lung cancer cells. Mol. Cancer Ther. 2011, 10, 1720–1727. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Crawford, M.; Mao, Y.; Lee, R.J.; Davis, I.C.; Elton, T.S.; Lee, L.J.; Nana-Sinkam, S.P. Therapeutic delivery of microRNA-29b by cationic lipoplexes for lung cancer. Mol. Ther. Nucleic Acids 2013, 2. [Google Scholar] [CrossRef]
- Wiggins, J.F.; Ruffino, L.; Kelnar, K.; Omotola, M.; Patrawala, L.; Brown, D.; Bader, A.G. Development of a lung cancer therapeutic based on the tumor suppressor microRNA-34. Cancer Res. 2010, 70, 5923–5930. [Google Scholar] [CrossRef] [PubMed]
- Chiou, G.Y.; Cherng, J.Y.; Hsu, H.S.; Wang, M.L.; Tsai, C.M.; Lu, K.H.; Chien, Y.; Hung, S.C.; Chen, Y.W.; Wong, C.I.; et al. Cationic polyurethanes-short branch PEI-mediated delivery of mirR145 inhibited epithelial-mesenchymal transdifferentiation and cancer stem-like properties and in lung adenocarcinoma. J. Control. Release 2012, 159, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Zhang, Y.X.; Wang, P.Y.; Chi, Y.L.; Zhang, C.; Ma, Y.; Lv, C.J.; Xie, S.Y. Regression of A549 lung cancer tumors by anti-miR-150 vector. Oncol. Rep. 2012, 27, 129–134. [Google Scholar] [PubMed]
- Cortez, M.A.; Valdecanas, D.; Zhang, X.; Zhan, Y.; Bhardwaj, V.; Calin, G.A.; Komaki, R.; Giri, D.K.; Quini, C.C.; Wolfe, T.; et al. Therapeutic delivery of miR-200c enhances radiosensitivity in lung cancer. Mol. Ther. 2014, 22, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Fortunato, O.; Boeri, M.; Verri, C.; Moro, M.; Sozzi, G. Therapeutic use of microRNAs in lung cancer. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.C.; Rossi, J.J. RNA-based therapeutics: Current progress and future prospects. Chem. Biol. 2012, 19, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Merkel, O.M.; Rubinstein, I.; Kissel, T. siRNA delivery to the lung: What’s new? Adv. Drug Deliv. Rev. 2014, 75, 112–128. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.L.; Bartz, S.R.; Schelter, J.; Kobayashi, S.V.; Burchard, J.; Mao, M.; Li, B.; Cavet, G.; Linsley, P.S. Expression profiling reveals off-target gene regulation by RNAi. Nat. Biotechnol. 2003, 21, 635–637. [Google Scholar] [CrossRef] [PubMed]
- Bridge, A.J.; Pebernard, S.; Ducraux, A.; Nicoulaz, A.L.; Iggo, R. Induction of an interferon response by RNAi vectors in mammalian cells. Nat. Genet. 2003, 34, 263–264. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Lu, J.J.; Chen, J.; Klibanov, A.M. Non-viral siRNA delivery to the lung. Adv. Drug Deliv. Rev. 2007, 59, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.H.; Chae, S.Y.; Bae, Y.H.; Kim, S.W. Synthesis of biodegradable multi-block copolymers of poly(l-lysine) and poly(ethylene glycol) as a non-viral gene carrier. J. Control. Release 2004, 97, 567–574. [Google Scholar] [PubMed]
- Bramson, J.L.; Bodner, C.A.; Graham, R.W. Activation of host antitumoral responses by cationic lipid/DNA complexes. Cancer Gene Ther. 2000, 7, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, M.; Li, S.; Huang, L. LPD lipopolyplex initiates a potent cytokine response and inhibits tumor growth. Gene Ther. 1999, 6, 1867–1875. [Google Scholar] [CrossRef] [PubMed]
- Hamasaki, T.; Suzuki, H.; Shirohzu, H.; Matsumoto, T.; D’Alessandro-Gabazza, C.N.; Gil-Bernabe, P.; Boveda-Ruiz, D.; Naito, M.; Kobayashi, T.; Toda, M.; et al. Efficacy of a novel class of RNA interference therapeutic agents. PLoS One 2012, 7, e42655. [Google Scholar] [CrossRef] [PubMed]
- Honma, K.; Iwao-Koizumi, K.; Takeshita, F.; Yamamoto, Y.; Yoshida, T.; Nishio, K.; Nagahara, S.; Kato, K.; Ochiya, T. RPN2 gene confers docetaxel resistance in breast cancer. Nat. Med. 2008, 14, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Yagishita, S.; Takeshita, F.; Yamamoto, Y.; Kuwano, K.; Ochiya, T. Prognostic and therapeutic impact of RPN2-mediated tumor malignancy in non-small-cell lung cancer. Oncotarget 2015, 6, 3335–3345. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujita, Y.; Kuwano, K.; Ochiya, T. Development of Small RNA Delivery Systems for Lung Cancer Therapy. Int. J. Mol. Sci. 2015, 16, 5254-5270. https://doi.org/10.3390/ijms16035254
Fujita Y, Kuwano K, Ochiya T. Development of Small RNA Delivery Systems for Lung Cancer Therapy. International Journal of Molecular Sciences. 2015; 16(3):5254-5270. https://doi.org/10.3390/ijms16035254
Chicago/Turabian StyleFujita, Yu, Kazuyoshi Kuwano, and Takahiro Ochiya. 2015. "Development of Small RNA Delivery Systems for Lung Cancer Therapy" International Journal of Molecular Sciences 16, no. 3: 5254-5270. https://doi.org/10.3390/ijms16035254
APA StyleFujita, Y., Kuwano, K., & Ochiya, T. (2015). Development of Small RNA Delivery Systems for Lung Cancer Therapy. International Journal of Molecular Sciences, 16(3), 5254-5270. https://doi.org/10.3390/ijms16035254