
Psoralea glandulosa as a Potential Source of Anticancer Agents for Melanoma Treatment

Abstract
:1. Introduction

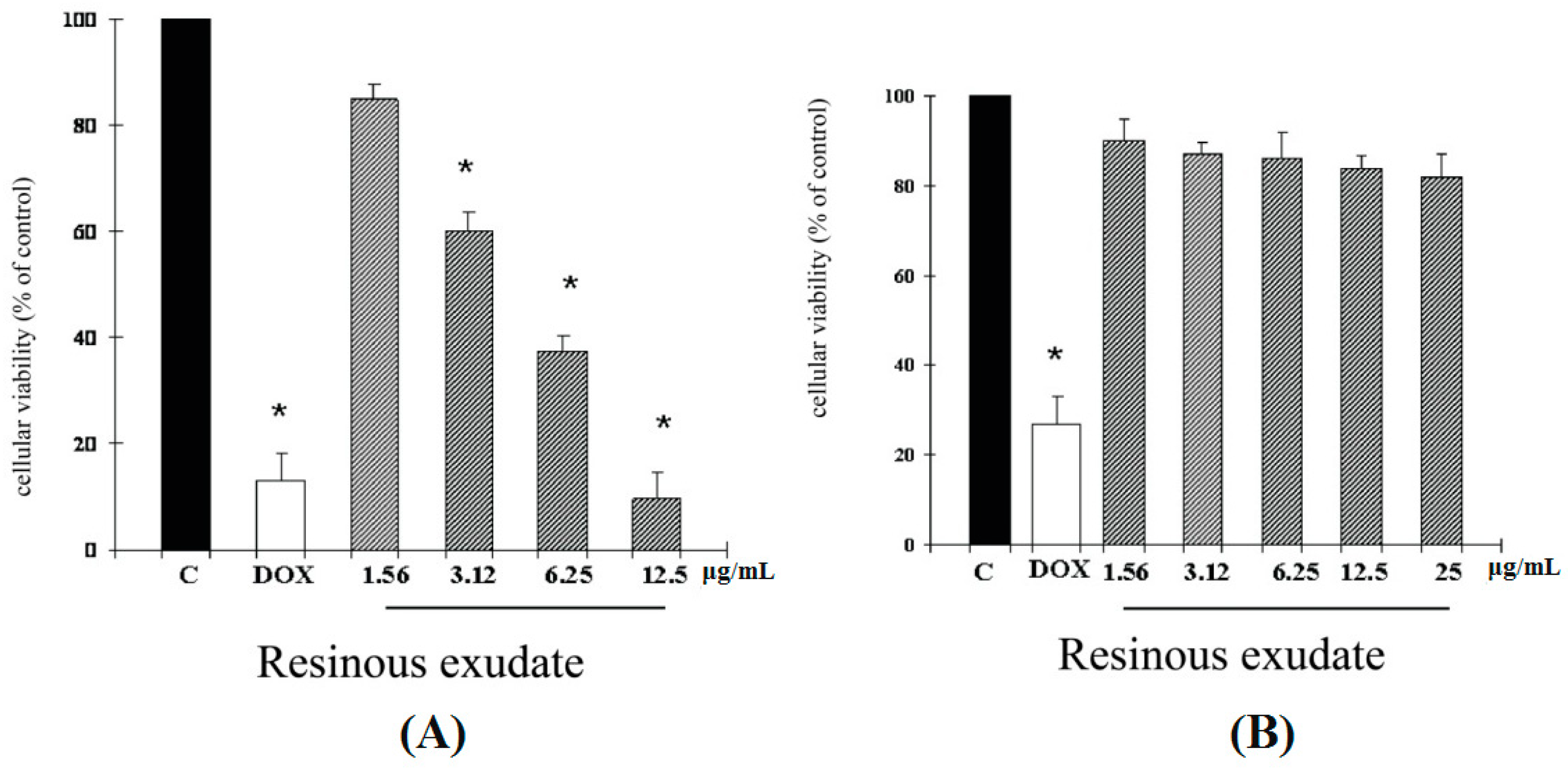
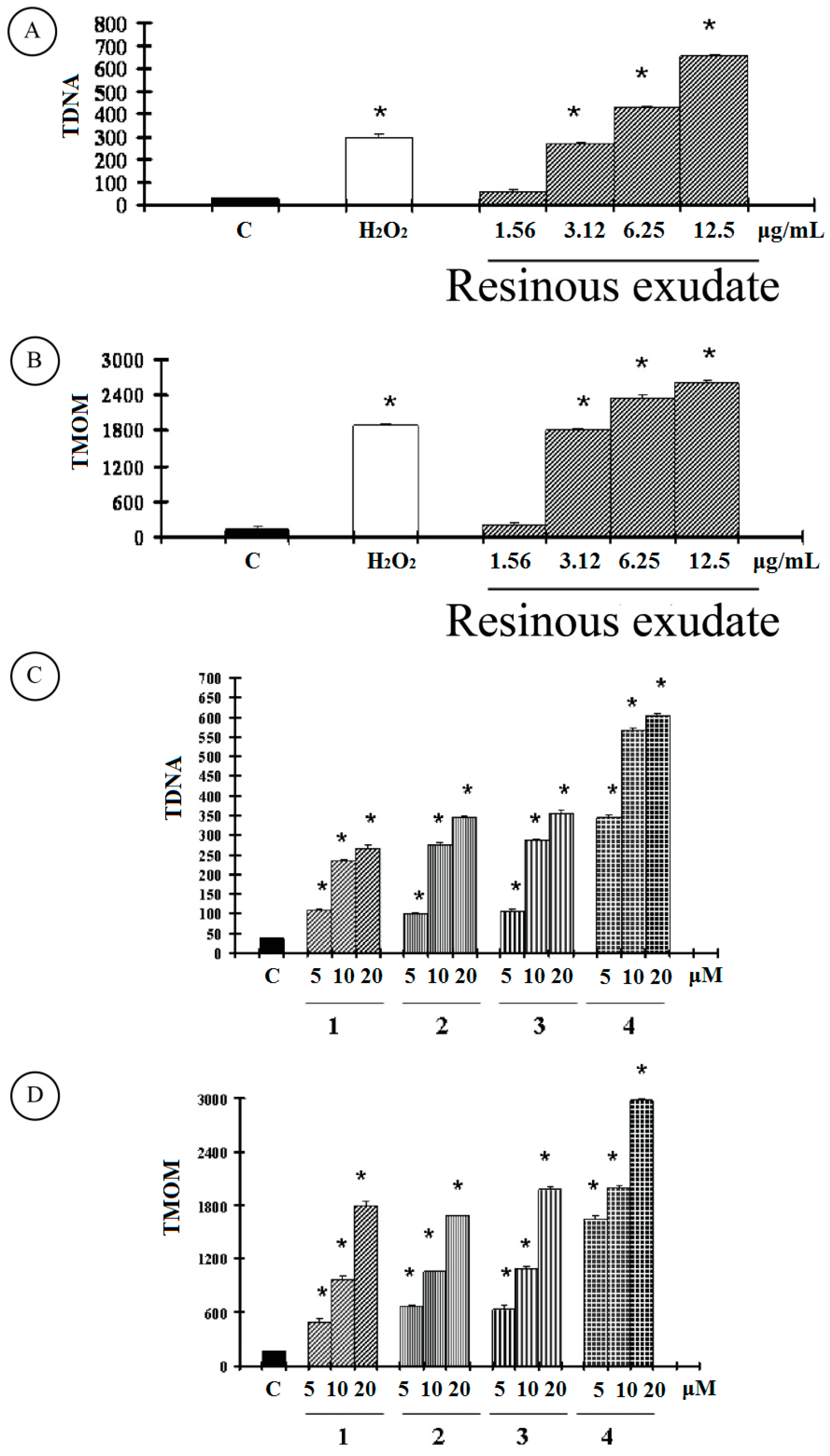
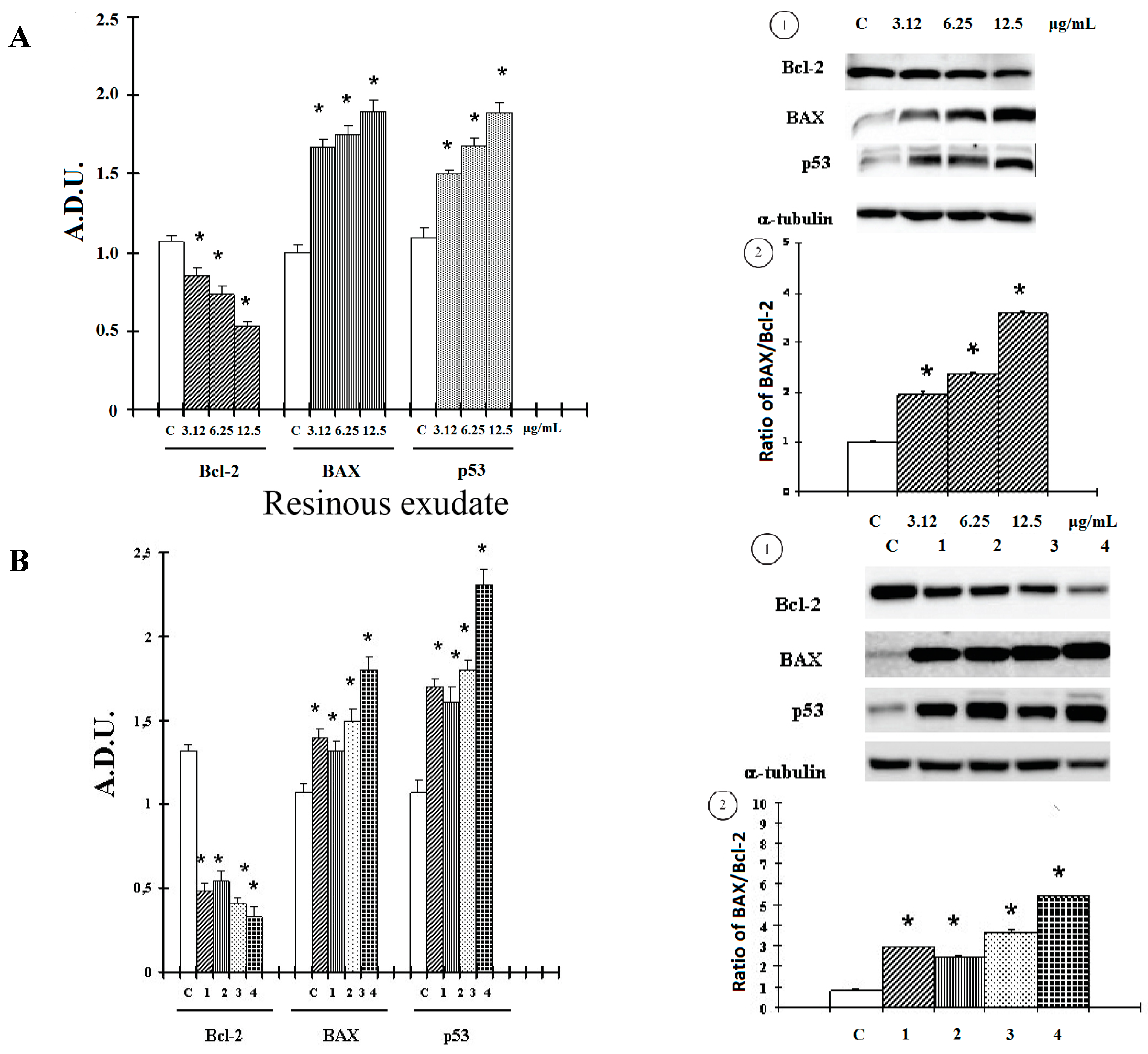
2. Results and Discussion
2.1. Results


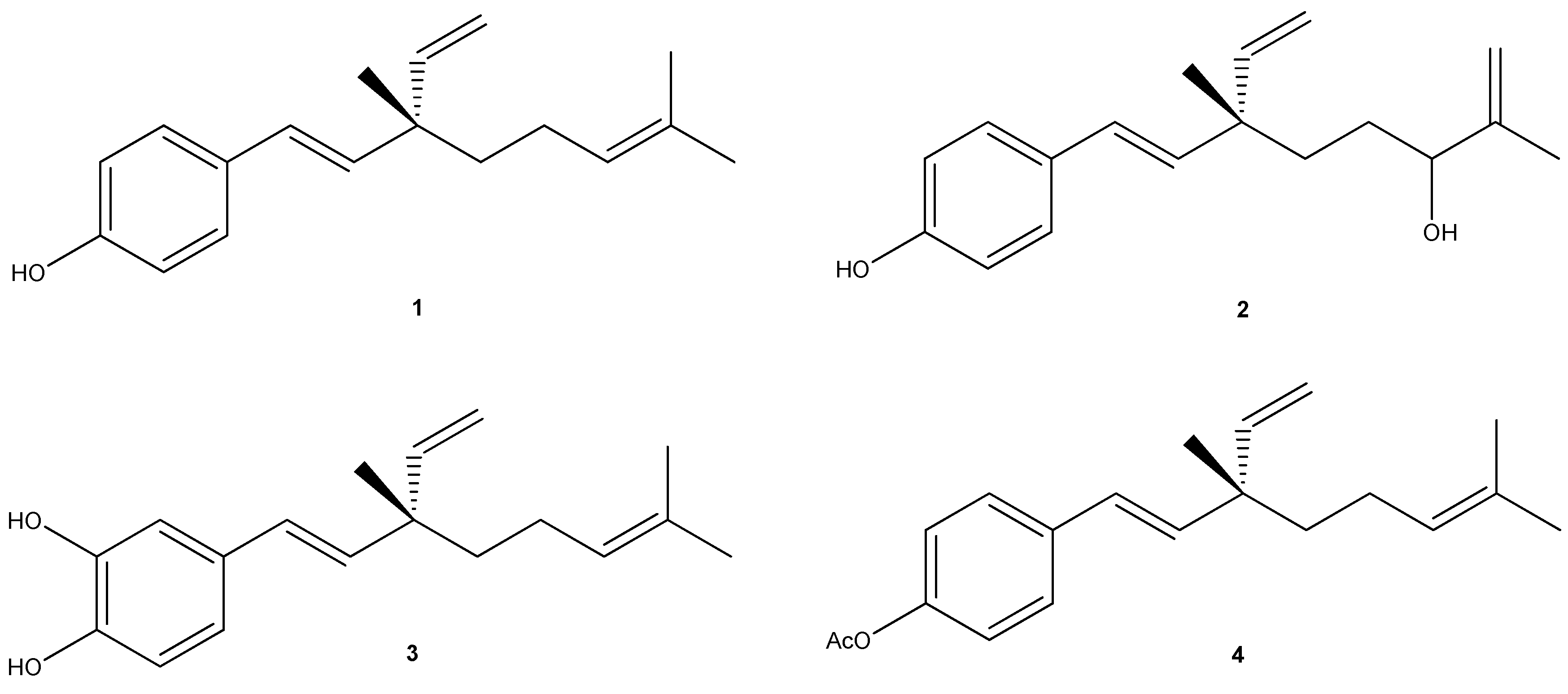{kind=link}
{kind=link}
{kind=link}
{kind=link}
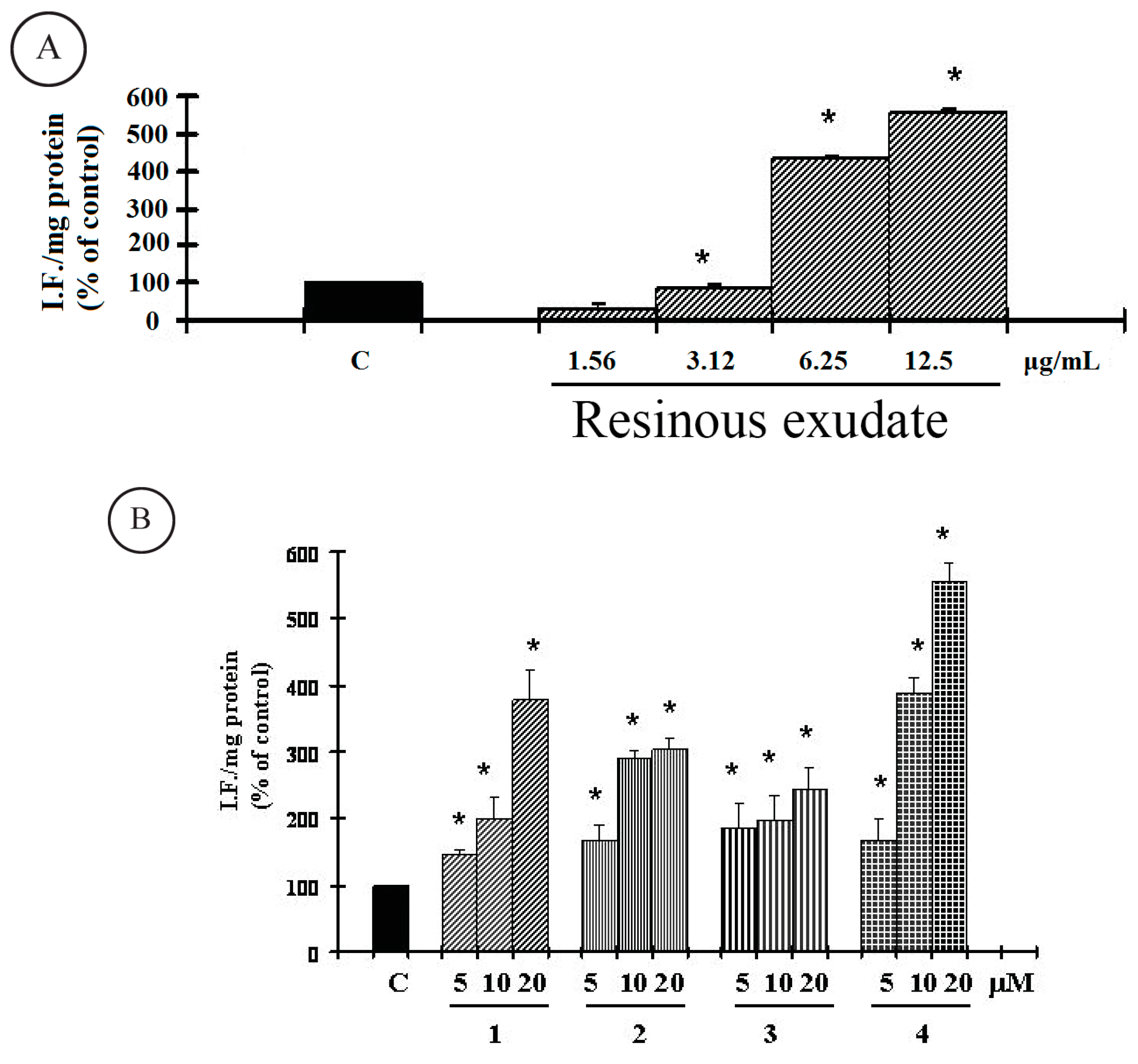{kind=link}
| Sample | IC50 a (µM) | % LDH b |
|---|---|---|
| 1 | 29.3 ± 0.4 | 5.9 ± 0.6 |
| 2 | 32.3 ± 0.6 | 6.1 ± 0.5 |
| 3 | 35.1 ± 0.5 | 6.9 ± 0.4 |
| 4 | 16.9 ± 0.4 | 7.3 ± 0.4 |
| Doxorubicin | 0.20 ± 0.01 | - |
| Negative Control | 100 | 6.5 ± 0.9 |
| Sample | Treatment | %LDH a |
|---|---|---|
| Resinous exudate | 3.12 μg/mL | 7.5 ± 0.6 |
| 6.25 μg/mL | 9.7 ± 0.5 | |
| 12.5 μg/mL | 8.9 ± 0.7 | |
| 25 μg/mL | 8.8 ± 0.6 | |
| H2O2 | 1 μM | 5.8± 0.7 |
| 1000 μM | 71.3± 1.7 * | |
| Negative control | 0.25% | 7.5 ± 0.7 |

| Sample | Treatment | OD405 nm/mg Protein |
|---|---|---|
| Resinous exudate | 3.12 μg/mL | 0.41 ± 0.03 * |
| 6.25 μg/mL | 0.55 ± 0.06 * | |
| 12.5 μg/mL | 0.84 ± 0.05 * | |
| Compound 1 | 5 μM | 0.45 ± 0.05 * |
| 10 μM | 0.63 ± 0.04 * | |
| 20 μM | 0.89 ± 0.09 * | |
| Compound 2 | 5 μM | 0.38 ± 0.06 * |
| 10 μM | 0.55 ± 0.04 * | |
| 20 μM | 0.77 ± 0.09 * | |
| Compound 3 | 5 μM | 0.39 ± 0.04 * |
| 10 μM | 0.62 ± 0.04 * | |
| 20 μM | 0.81 ± 0.06 * | |
| Compound 4 | 5 μM | 0.65 ± 0.03 * |
| 10 μM | 0.73 ± 0.04 * | |
| 20 μM | 0.97 ± 0.06 * | |
| H2O2 | 1 μM | 0.89 ± 0.09 * |
| Negative control | 0.25% | 0.27 ± 0.06 |


2.2. Discussion
3. Experimental Section
3.1. Plant Material
3.2. Chemical and Reagents
3.3. Preparation of the Resinous Exudates
3.4. Isolation of Natural Compounds 1–3
3.5. Procedure for Synthesis of Compound 4
3.6. Cell Culture
3.7. MTT Bioassay
3.8. Lactate Dehydrogenase (LDH) Release
3.9. Activity of Caspase-3
3.10. DNA Analysis by COMET Assay
3.11. Western Blot Analysis
3.12. Reactive Oxygen Species Assay
3.13. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Smalley, K.S. Understanding melanoma signaling networks as the basis for molecular targeted therapy. J. Investig. Dermatol. 2010, 130, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Madrid, A.; Espinoza, L.; González, C.; Mellado, M.; Villena, J.; Santander, R.; Silva, V.; Montenegro, I. Antifungal study of the resinous exudate and of meroterpenoids isolated from Psoralea glandulosa (Fabaceae). J. Ethnopharmacol. 2012, 144, 809–811. [Google Scholar] [CrossRef] [PubMed]
- Madrid, A.; Espinoza, L.; Mellado, M.; Montenegro, I.; González, C.; Santander, R.; Villena, J.; Jara, C. Study of the chemical composition of the resinous exudate isolated from Psoralea glandulosa and evaluation of the antioxidant properties of the terpenoids and the resin. Blacpma 2013, 12, 338–345. [Google Scholar]
- Majeed, R.; Reddy, M.V.; Chinthakindi, P.K.; Sangwan, P.L.; Hamid, A.; Chashoo, G.; Saxena, A.K.; Koul, S. Bakuchiol derivatives as novel and potent cytotoxic agents: A report. Eur. J. Med. Chem. 2012, 49, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Labbé, C.; Faini, F.; Coll, J.; Connolly, J. Bakuchiol derivatives from the leaves of Psoralea glandulosa. Phytochemistry 1996, 42, 1299–1303. [Google Scholar] [CrossRef]
- Russo, A.; Piovano, M.; Lombardo, L.; Garbarino, J.; Cardile, V. Lichen metabolites prevent UV light and nitric oxide-mediated plasmid DNA damage and induce apoptosis in human melanoma cells. Life Sci. 2008, 83, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, R.W.; Ruefli, A.A.; Lowe, S.W. Apoptosis: A link between cancer genetics and chemotherapy. Cell 2002, 108, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Bednarek, I.; Sypniewski, D.; Klama-Baryla, A.; Galka, S.; Machnik, G. Single-cell gel electrophoresis (comet assay) as a tool for apoptosis determination in tumor cell lines HL-60 and Jurkat cultures treated with anisomycin. Ann. Acad. Med. 2006, 60, 278–284. [Google Scholar]
- Schulze-Osthoff, K.; Bakker, A.C.; Vanhaesebroeck, B.; Beyaert, R.; Jacob, W.A.; Fiers, W. Cytotoxic activity of tumor necrosis factor is mediated by early damage of mitochondrial functions: Evidence for the involvement of mitochondrial radical generation. J. Biol. Chem. 1992, 267, 5317–5323. [Google Scholar] [PubMed]
- Callagy, G.M.; Webber, M.J.; Pharoah, P.D.; Caldas, C. Meta-analysis confirms Bcl2 is an independent prognostic marker in breast cancer. BMC Cancer 2008, 8, 153. [Google Scholar] [CrossRef]
- Krajewski, S.; Krajewska, M.; Turner, B.C.; Pratt, C.; Howard, B.; Zapata, J.M.; Frenkel, V.; Robertson, S.; Ionov, Y.; Yamamoto, H.; et al. Prognostic significance of apoptosis regulators in breast cancer. Endocr. Relat. Cancer 1999, 6, 29–40. [Google Scholar]
- Kingston, C.; Jeeva, S.; Jeeva, G.M.; Kiruba, S.; Mishra, B.P.; Kannan, D. Indigenous Knowledge of using medicinal plants in treating skin diseases in Kanyakumari district, Southern India. Indian J. Tradit. Knowl. 2009, 8, 196–200. [Google Scholar]
- De Wet, H.; Nciki, S.; van Vuuren, S.F. Medicinal plants used for the treatment of various skin disorders by a rural community in northern Maputaland, South Africa. J. Ethnobiol. Ethnomed. 2013, 51, 1–9. [Google Scholar]
- Bapat, K.; Chintalwar, G.J.; Pandey, U.; Thakur, V.S.; Sarma, H.D.; Samuel, G.; Pillai, M.R.; Chattopadhyay, S.; Venkatesh, M. Preparation and in vitro evaluation of radioiodinatedbakuchiol as an anti tumor agent. Appl. Radiat. Isot. 2005, 62, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Katsura, H.; Tsukiyama, R.I.; Suzuki, A.; Kobayashi, M. In vitro antimicrobial activities of bakuchiol against oral microorganisms. Antimicrob. Agents Chemother. 2001, 45, 3009–3013. [Google Scholar] [CrossRef] [PubMed]
- Backhouse, C.N.; Delporte, C.L.; Negrete, R.E.; Erazo, S.; Zuniga, A.; Pinto, A.; Cassels, B.K. Active constituents isolated from Psoralea glandulosa L. with antiinflammatory and antipyretic activities. J. Ethnopharmacol. 2001, 78, 27–31. [Google Scholar]
- Park, E.; Zhao, Y.; Kim, Y.; Sohn, D. Bakuchiol-induced caspase-3-dependent apoptosis occurs through c-Jun NH2-terminal kinase-mediated mitochondrial translocation of Bax in rat liver myofibroblasts. Eur. J. Pharmacol. 2007, 559, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Cha, M.R.; Choi, C.W.; Lee, J.Y.; Kim, Y.S.; Yon, G.H.; Choi, S.; Ryu, S.H. Anti-proliferative effect of synthesized bakuchiol analogues on cultured human tumor cell lines. Bull. Korean Chem. Soc. 2012, 33, 2378–2380. [Google Scholar] [CrossRef]
- Jaña, F.; Faini, F.; Lapier, M.; Pavani, M.; Kemmerling, U.; Morello, A.; Maya, J.D.; Jara, J.; Parra, E.; Ferreira, J.; et al. Tumor cell death induced by the inhibition of mitochondrial electron transport: The effect of 3-hydroxybakuchiol. Toxicol. Appl. Pharmacol. 2013, 272, 356–364. [Google Scholar]
- Ohno, O.; Watabe, T.; Nakamura, K.; Kawagoshi, M.; Uotsu, N.; Chiba, T.; Yamada, M.; Yamaguchi, K.; Yamada, K.; Miyamoto, Y.; et al. Inhibitory effects of bakuchiol, bavachin, and isobavachalcone isolated from Piper longum on melanin production in B16 mouse melanoma cells. Biosci. Biotechnol. Biochem. 2010, 74, 1504–1506. [Google Scholar]
- Chopra, B.; Dhingra, A.K.; Dhar, K.L. Psoralea corylifolia L. (Buguchi)-Folklore to modern evidence: Review. Fitoterapia 2013, 90C, 44–56. [Google Scholar]
- Chen, Z.; Jin, K.; Gao, L.; Lou, G.; Jin, Y.; Yu, Y.; Lou, Y. Anti-tumor effects of bakuchiol, an analogue of resveratrol, on human lung adenocarcinoma A549 cell line. Eur. J. Pharmacol. 2010, 643, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Newman, D.J. Plants as a source of anti-cancer agents. J. Ethnopharmacol. 2005, 100, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.B.; Ye, Y.P.; Wu, X.D.; Sun, X.S. A stilbotriterpenic acid induces growth arrest and apoptosis in HeLa cells through mitochondria-related pathways and reactive oxygen species (ROS) production. Chem. Biodivers. 2009, 6, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Lim, W.; Kim, J.E.; Kim, I.; Jeong, J.; Ko, Y.; Song, J.; You, S.; Kim, D.; Kim, M.; et al. Cell death and intracellular distribution of hematoporphyrin in a KB cell line. Photomed. Laser Surg. 2009, 27, 453–460. [Google Scholar]
- Nguyen, D.M.; Hussain, M. The role of the mitochondria in mediating cytotoxicity of anti-cancer therapies. J. Bioenerg. Biomembr. 2007, 39, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.T.; Chang, H.L.; Shyue, S.K.; Hsu, S.L. Emodin induces apoptosis in human lung adenocarcinoma cells through a reactive oxygen species-dependent mitochondrial signaling pathway. Biochem. Pharmacol. 2005, 70, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Erazo, S.; García, R.; Delle Monache, F. Bakuchiol and others compounds from Psoralea glandulosa. Rev. Latinoam. Quim. 1990, 21, 62–62. [Google Scholar]
- Reddy, V.M.; Thota, N.; Sangwan, P.L.; Malhotra, P.; Ali, F.; Khan, I.A.; Chimni, S.S.; Koul, S. Novel bisstyryl derivatives of bakuchiol: Targeting oral cavity pathogens. Eur. J. Med. Chem. 2010, 45, 3125–3134. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Espinoza, L.; Caggia, S.; Garbarino, J.A.; Pena-Cortes, H.; Carvajal, M.; Cardile, V. A new jasmonic acid stereoisomeric derivative induces apoptosis via reactive oxygen species in human prostate cancer cells. Cancer Lett. 2012, 326, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; Tice, R.R.; Stephens, R.E.; Scheneider, E.L. A microgel electrophoresis technique for the direct quantitation of DNA damage and repair in individual fibroblasts cultured on microscope slides. Mutat. Res. 1991, 252, 289–296. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madrid, A.; Cardile, V.; González, C.; Montenegro, I.; Villena, J.; Caggia, S.; Graziano, A.; Russo, A. Psoralea glandulosa as a Potential Source of Anticancer Agents for Melanoma Treatment. Int. J. Mol. Sci. 2015, 16, 7944-7959. https://doi.org/10.3390/ijms16047944
Madrid A, Cardile V, González C, Montenegro I, Villena J, Caggia S, Graziano A, Russo A. Psoralea glandulosa as a Potential Source of Anticancer Agents for Melanoma Treatment. International Journal of Molecular Sciences. 2015; 16(4):7944-7959. https://doi.org/10.3390/ijms16047944
Chicago/Turabian StyleMadrid, Alejandro, Venera Cardile, César González, Ivan Montenegro, Joan Villena, Silvia Caggia, Adriana Graziano, and Alessandra Russo. 2015. "Psoralea glandulosa as a Potential Source of Anticancer Agents for Melanoma Treatment" International Journal of Molecular Sciences 16, no. 4: 7944-7959. https://doi.org/10.3390/ijms16047944